
Metabolic Consequences of the Water We Drink: A Study Based on Field Evidence and Animal Model Experimentation

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. DDD and DDE
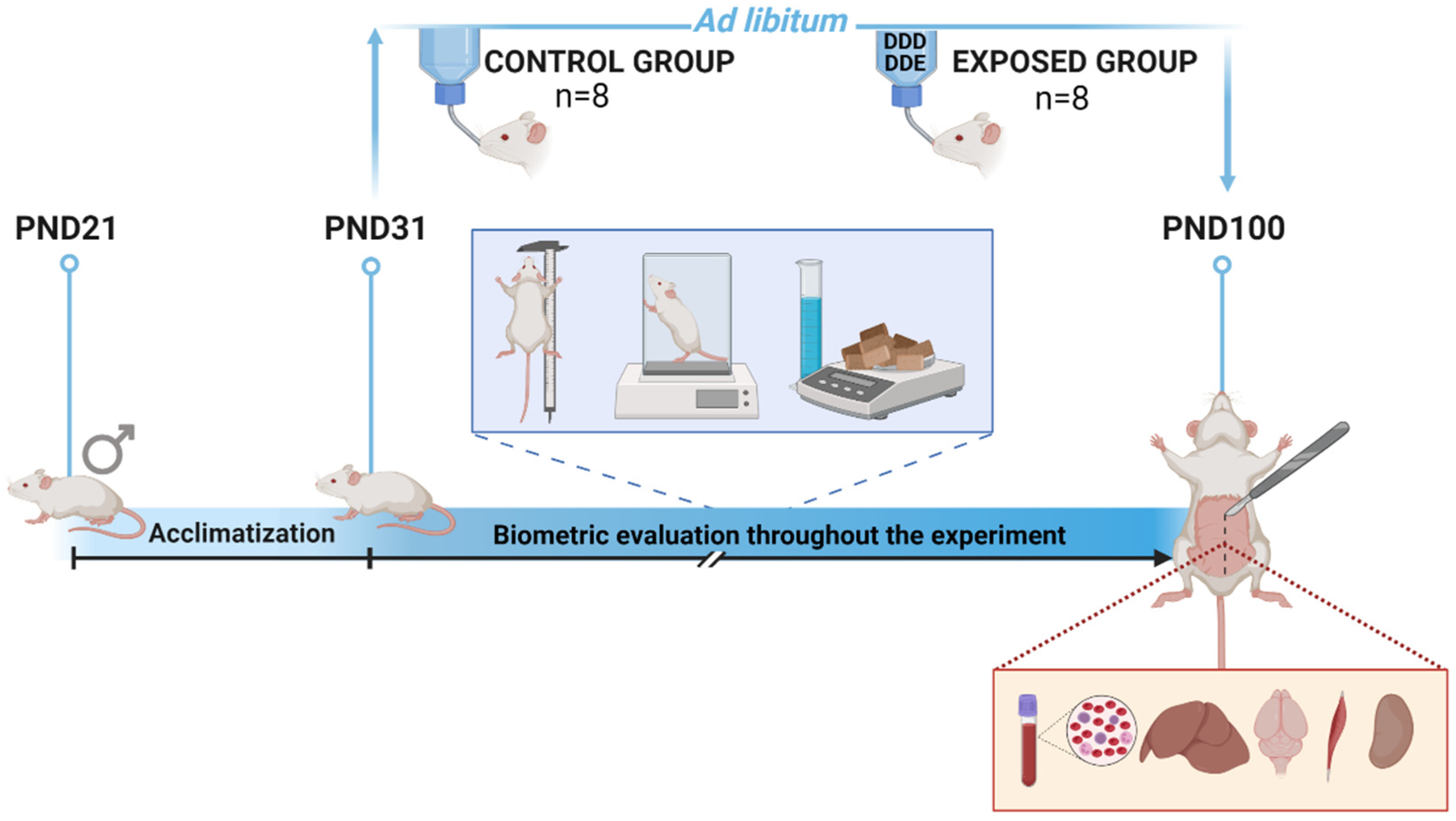
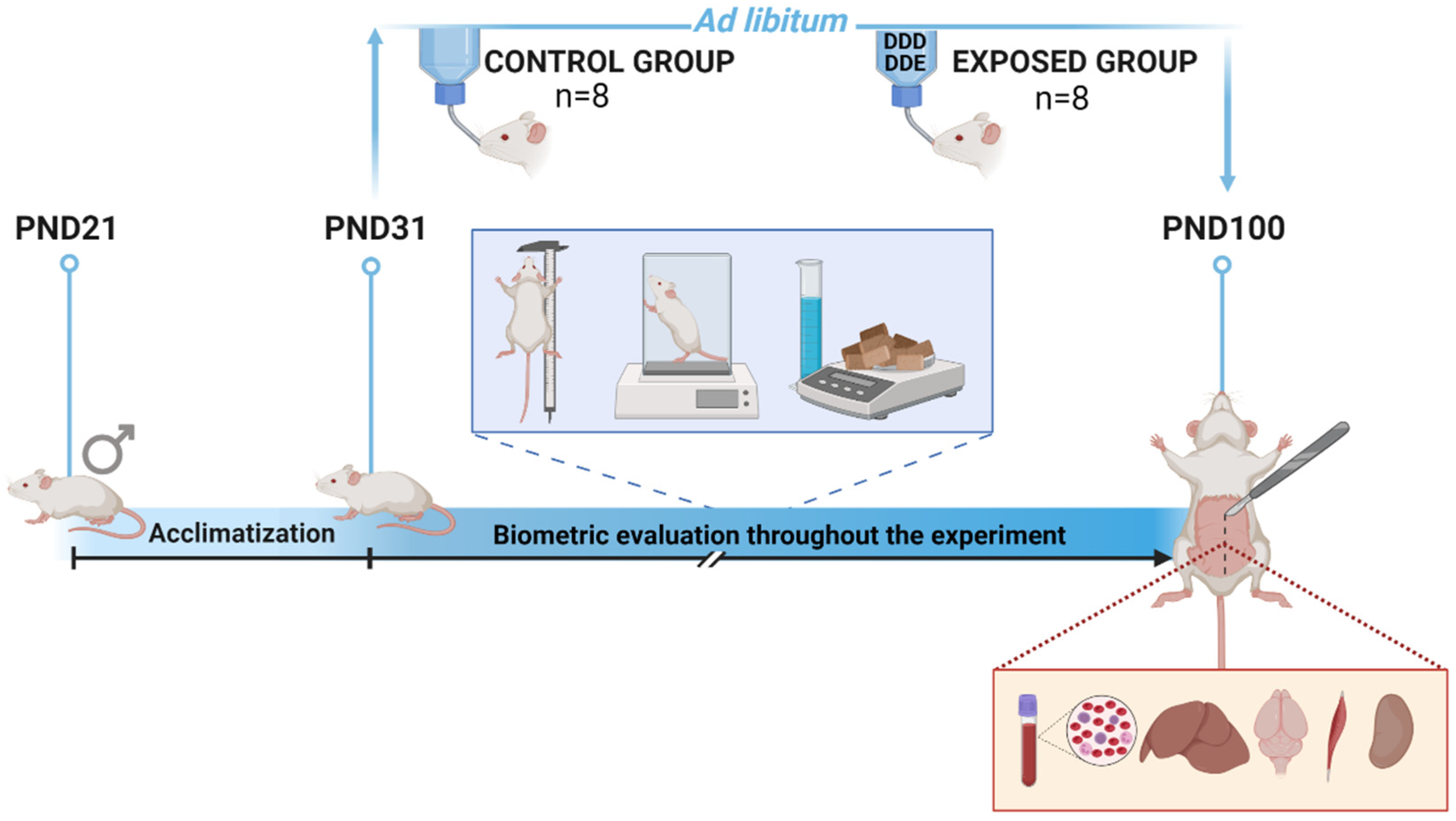
2.2. Experimental Design
2.3. Biometric Evaluation
2.4. Blood and Tissue Sampling
2.5. Antioxidant System and Liver Function Analyses
2.6. Statistical Analysis
3. Results
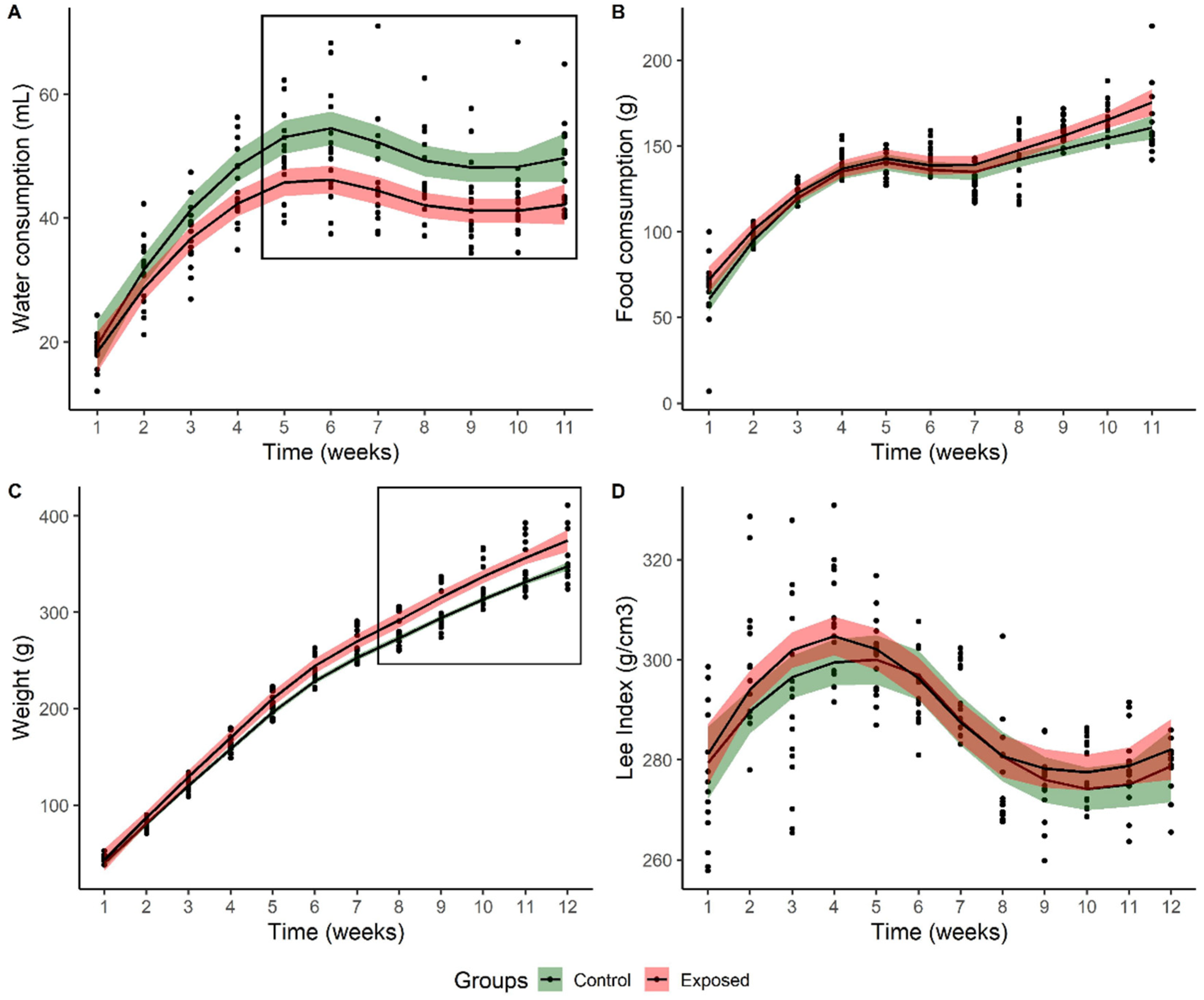
3.1. Biometric Variables
3.2. Blood Analyses
3.3. Antioxidant System and Damage Tissue Analyses
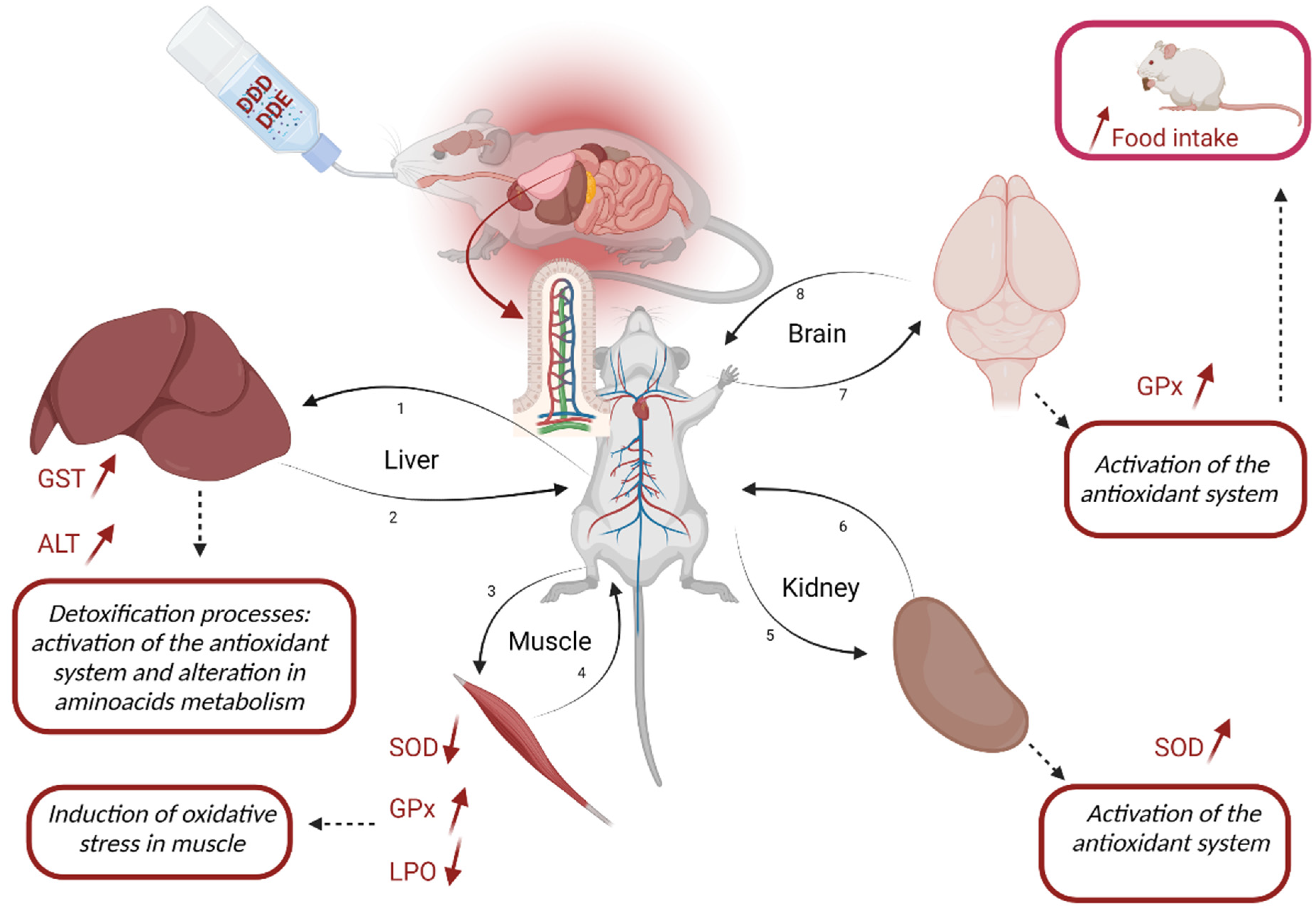
3.4. Integrative Analyses of the Antioxidant System and Tissue Damage
4. Discussion
4.1. Liver Impact
4.2. Muscle Impact
4.3. Kidney Impact
4.4. Brain Impact
4.5. Biometric Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vieira:, K.C.; Silva, C.T.; Silva, M.M.; Costa, A.S.V. Potential for environmental contamination of herbicides used in corn, soy and sugar cane cultures. Res. Soc. Dev. 2020, 9, e417997442. [Google Scholar] [CrossRef]
- Ismael, L.L.; Rocha, E.M.R. Estimativa de contaminação de águassubterrâneas e superficiaisporagrotóxicosemáreasucroalcooleira, Santa Rita/PB, Brasil. Ciência&SaúdeColetiva 2019, 24, 4665–4676. [Google Scholar]
- Santos, G.D.; Santos, J.L.O.; Leite, O.D. Avaliação do risco de contaminação das águas superficiais e subterrâneaspelo paraquat no perímetro irrigado Formoso, Bom Jesus da Lapa/Bahia. In SimpósioBrasileiro de RecursosHídricos, 12, 2017; ABRH—AssociaçãoBrasileira de RecursosHídricos: Florianópolis, SC, Brazil, 2017. [Google Scholar]
- Chaves, H.M.L.; Souza, M.A. Índice para a avaliação do risco de contaminação da águaporpesticidas: Desenvolvimento e validação. Rev. De Recur. 2015, 20, 286–297. [Google Scholar]
- Ribeiro, M.L.; Lourencetti, C.; Pereira, S.Y.; Marchi, M.R.R. Contaminação de águassubterrâneasporpesticidas: Avaliaçãopreliminar. Quim. Nova 2007, 30, 688–694. [Google Scholar] [CrossRef]
- Martini, L.F.D.; Caldas, S.S.; Bolzan, C.M.; Bundt, A.D.C.; Primel, E.G.; Avila, L.A. Risco de contaminação das águas de superfície e subterrâneasporagrotóxicosrecomendados para a cultura do arrozirrigado. Ciência Rural. 2012, 42, 1715–1721. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.A.; Correia, N.M.; Silva, J.R.M.; Velini, E.D.; Passos, A.B.R.J.; Durigan, J.C. Herbicide detection in groundwater in Córrego Rico-SP watershed. Rev. Planta Daninha 2015, 33, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, A.; Moraes, J.; Da Silva, M.R. Pesticidas no perfil de solos emáreas de plantação de cebolasemItuporanga, SC. Rev. De Eng. Agric. E Ambient.-Agriambiental 2011, 15, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.R.O.; Avila, L.A.; Agostinetto, D.; Bundt, A.D.C.; Primel, E.G.; CALDAS, S.S. Ocorrência de agrotóxicosemáguassubterrâneas de áreasadjacentes a lavouras de arrozirrigado. Quim. Nova 2011, 34, 748–752. [Google Scholar] [CrossRef] [Green Version]
- Brasil, Ministério da Saúde. Controle do Sistema de Informação de Vigilância da Qualidade da Água para Consumo Humano (SISAGUA). Available online: http://sisagua.saude.gov.br/sisagua/paginaExterna.jsf (accessed on 1 May 2022).
- Brasil. Trata. Instituto Trata Brasil. Águas Subterrâneas. 2019. Available online: https://tratabrasil.org.br/aguas-subterraneas-e-saneamento-basico/ (accessed on 15 January 2021).
- IBGE, Instituto de Geografia e Estatística. Panorama Municipal Santa Tereza do Oeste. Available online: https://cidades.ibge.gov.br/brasil/pr/santa-tereza-do-oeste/panorama (accessed on 17 June 2020).
- SIAGRO. Software Integrado do Agronegócio. Agrotóxicos no Paraná. Available online: http://www.adapar.pr.gov.br/pagina-389.html (accessed on 6 February 2020).
- Carneiro, F.F. (Org.) Dossiê ABRASCO: Um Alertasobreosimpactos dos Agrotóxicosnasaúde; EPSJV: Rio de Janeiro, RJ, Brazil; Expressão Popular: São Paulo, SP, Brazil, 2015. [Google Scholar]
- Faroon, O.; Ruiz, P.; Jones, D.; Keith, S.; Scinicariello, F.; McClure, P.; Llados, F.; Ingerman, L.; Citra, M.; Stevens, S. Toxicological Profile for DDT, DDE, and DDD: Draft for Public Comment; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2019. [Google Scholar]
- Tomiyama, N.; Takeda, M.; Watanabe, M.; Kobayashi, H.; Harada, T. A further study on the reliability of toxicokinetic parameters for predicting hepatotoxicity in rats receiving a 28-day repeated administration of DDT. J. Toxicol. Sci. 2004, 29, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Harada, T.; Ohtsuka, R.; Takeda, M.; Yoshida, T.; Enomoto, A.; Kojima, S.; Tomiyama, N.; Nakashima, N.; Ozaki, M. Hepatocarcinogenesis by DDT in Rats. J. Toxicol. Pathol. 2006, 19, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.R.; Roy, A.; Shalat, S.L.; Vonstein, R.T.; Hossain, M.M.; Buckley, B.; Gearing, M.; Levey, A.I.; German, D.C. Elevated serum pesticide levels and risk for Alzheimer disease. JAMA Neurol. 2014, 71, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Lee, Y.M.; Lee, H.W.; Jacobsjr, D.R.; Lee, D.-H. Associations between organochlorine pesticides and cognition in U.S. elders: National Health and Nutrition Examination Survey 1999–2002. Environ. Int. 2015, 75, 87–92. [Google Scholar] [CrossRef]
- Chiang, C.; Mahalingam, S.; Flaws, J.A. Environmental contaminants affecting fertility and somatic Health. In Seminars in Reproductive Medicine; NIH Public Access: New York, NY, USA, 2017; p. 241. [Google Scholar] [CrossRef]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: Effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Steffes, M.W.; Sjödin, A.; Jones, R.S.; Needham, L.L.; Jacobsjr, D.R. Low dose organochlorine pesticides and polychlorinated biphenyls predict obesity, dyslipidemia, and insulin resistance among people free of diabetes. PLoS ONE 2011, 6, e15977. [Google Scholar] [CrossRef] [Green Version]
- Roos, V.; Rönn, M.; Salihovic, S.; Lind, L.; Vanbavel, B.; Kullberg, J.; Johansson, L.; Ahlström, H.; Lind, P.M. Circulating levels of persistent organic pollutants in relation to visceral and subcutaneous adipose tissue by abdominal MRI. Obesity 2013, 21, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, J.P.; Ocaña-Riola, R.; Arrebola-Moreno, A.L.; Fernández-Rodríguez, M.; Martin-Olmedo, P.; Fernández, M.F.; Olea, N. Associations of accumulated exposure to persistent organic pollutants with serum lipids and obesity in an adult cohort from Southern Spain. Environ. Pollut. 2014, 195, 9–15. [Google Scholar] [CrossRef]
- Tang-Péronard, J.L.; Heitmann, B.L.; Jensen, T.K.; Vinggaard, A.M.; Madsbad, S.; Steuerwald, U.; Grandjean, P.; Weihe, P.; Nielsen, F.; Andersen, H.R. Prenatal exposure to persistent organochlorine pollutants is associated with high insulin levels in 5-year-old girls. Environ. Res. 2015, 142, 407–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gascon, M.; Vrijheid, M.; Martínez, D.; Ballester, F.; Basterrechea, M.; Blarduni, E.; Esplugues, A.; Vizcaino, E.; Grimalt, J.O.; Morales, E.; et al. Pre-natal exposure to dichlorodiphenyldichloroethylene and infant lower respiratory tract infections and wheeze. Eur. Respir. J. 2012, 39, 1188–1196. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.; Strøm, M.; Olsen, S.F.; Maslova, E.; Rantakokko, P.; Kiviranta, H.; Rytter, D.; Bech, B.H.; Hansen, L.V.; Halldorsson, T.I. Maternal concentrations of persistent organochlorine pollutants and the risk of asthma in offspring: Results from a prospective cohort with 20 years of follow-up. Environ. Health Perspect. 2014, 122, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Musumeci, M.R. Defensivos agrícolas e sua interação com a microbiota do solo. In Microbiologia do Solo; Cardoso, E.J.B.N., Tsai, S.M., Neves, M.C.P., Eds.; Sociedade Brasileira de Ciência do Solo: Campinas, SP, Brazil, 1992. [Google Scholar]
- ATSDR, Agency for Toxic Substances and Disease Registry. DDT, DDE e DDD—ToxFAQs™. Available online: https://www.atsdr.cdc.gov/toxfaqs/ToxFAQS_Foreign_Language_PDFs/tfacts35_portuguese.pdf (accessed on 1 February 2021).
- Mansouri, A.; Cregut, M.; Abbes, C.; Durand, M.-J.; Landoulsi, A.; Thouand, G. The environmental issues of DDT pollution and bioremediation: A multidisciplinary review. Appl. Biochem. Biotechnol. 2016, 181, 309–339. [Google Scholar] [CrossRef]
- Almeida, F.V.; Centeno, A.J.; Bisinoti, M.C.; Jardim, W.F. Substâncias tóxicas persistentes (STP) no Brasil. Química Nova 2007, 30, 1976–1985. [Google Scholar] [CrossRef] [Green Version]
- Kirsch, C.B.; Bach, A.; Prasniewski, V.; Paoletti, M.; Da Silva, K.A.; De Oliveira, C.M.R.; Szinwelski, N.; Guimarães, A.T.B.; Pereira, N.; Ferreira, N.C. Are forests providing their ecosystem service to guarantee the biological community? In Proceedings of the SETAC Europe 30th Annual Meeting, Virtual, 3–7 May 2020. [Google Scholar]
- Vasseghian, Y.; Hosseinzadeh, S.; Khataee, A.; Dragoi, E.-N. The concentration of persistent organic pollutants in water resources: A global systematic review, meta-analysis and probabilistic risk assessment. Sci. Total Environ. 2021, 796, 149000. [Google Scholar] [CrossRef] [PubMed]
- Pestana, D.; Teixeira, D.; Meireles, M.; Marques, C.; Norberto, S.; SÁ, C.; Fernandes, V.C.; Correia-Sá, L.; Faria, A.; Guardão, L.; et al. Adipose tissue dysfunction as a central mechanism leading to dysmetabolic obesity triggered by chronic exposure to p,p’-DDE. Sci. Rep. 2017, 7, 2738. [Google Scholar] [CrossRef] [PubMed]
- Briggs, C.; Bain, B.J. Basic Haematological Techniques. In Dacie and Lewis Practical Hematology, 12th ed.; Bain, B.J., Bates, I., Laffan, M.A., Eds.; Elsevier Health: Amsterdam, The Netherlands, 2017; pp. 18–49. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Crouch, R.K.; Gandy, S.E.; Kimsey, G.; Galbraith, R.A.; Galbraith, G.M.P.; BUSE, M.G. The Inhibition of Islet Superoxide Dismutase by Diabetogenic Drugs. Diabetes 1981, 30, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Keen, J.H.; Habig, W.H.; Jakoby, W.B. Mechanism for the several activities of the glutathione S-transferases. J. Biol. Chem. 1976, 251, 6183–6188. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. In Methods in Enzymology; Academic Press: New York, NY, USA, 1984; pp. 114–120. [Google Scholar] [CrossRef]
- Sies, H.; Koch, O.R.; Martino, E.; Boveris, A. Increased biliary glutathione disulfide release in chronically ethanol-treated rats. FEBS Lett. 1979, 103, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1975, 129, 302–310. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 May 2022).
- Fernandes, C.L.F.; Volcao, L.M.; Ramires, P.F.; Moura, R.R.; Silva Junior, F.M.R. Distribution of pesticides in agricultural and urban soils of Brazil: A critical review. Environ. Sci. Process. Impacts 2020, 22, 256–270. [Google Scholar] [CrossRef]
- Moreira, H.S.B.; Machado, R.A.; Aquino, S.N.D.; Rangel, A.L.C.A.; Martelli Júnior, H.; Coletta, R.D. Epidemiological features of patients with nonsyndromic cleft lip and/or palate in Western Parana. Braz. J. Oral Sci. 2016, 15, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Brasil, Lei Nº 11.936, de 14 de Maio de 2009. Proíbe a Fabricação, a Importação, a Exportação, a Manutençãoemestoque, a Comercialização e o Uso de Diclorodifeniltricloretano (DDT) e Dáoutrasprovidências. Available online: http://www.planalto.gov.br/ccivil_03/_ato2007-2010/2009/lei/l11936.htm (accessed on 17 June 2021).
- ATSDR, Agency for Toxic Substances and Disease Registry. Toxicological Profile for DDT, DDE, and DDD. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp35.pdf (accessed on 23 June 2022).
- Turusov, V.; Rakitsky, V.; Tomatis, L. Dichlorodiphenyltrichloroethane (DDT): Ubiquity, persistence, and risks. Environ. Health Perspect. 2002, 110, 125–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; LI, S.; Cirillo, P.; Krigbaum, N.; Tran, V.; Ishikawa, T.; Cohn, B. Metabolome Wide Association Study of serum DDT and DDE in Pregnancy and Early Postpartum. Reprod. Toxicol. 2020, 92, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, Q.; Xu, C.; Shao, W.; Zhang, C.; Liu, H.; Jiang, Z.; Gu, A. Organochloride pesticides impaired mitochondrial function in hepatocytes and aggravated disorders of fatty acid metabolism. Sci. Rep. 2017, 7, 46339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliaccio, V.; Scudiero, R.; Sica, R.; Lionetti, L.; Putti, R. Oxidative stress and mitochondrial uncoupling protein 2 expression in hepatic steatosis induced by exposure to xenobiotic DDE and high fat diet in male Wistar rats. PLoS ONE 2019, 14, e0215955. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Yin, S.; Tang, M.; Liu, K.; Yang, F.; Liu, W. Environmental exposure to DDT and its metabolites in cord serum: Distribution, enantiomeric patterns, and effects on infant birth outcomes. Sci. Total Environ. 2017, 580, 491–498. [Google Scholar] [CrossRef]
- Kao, C.C.; Que, D.E.; Bongo, S.J.; Tayo, L.L.; Lin, Y.H.; Lin, C.W.; Gou, Y.Y.; Hsu, W.L.; Shy, C.G.; Huang, K.L.; et al. Residue levels of organochlorine pesticides in breast milk and its associations with cord blood thyroid hormones and the offspring’s neurodevelopment. Int. J. Environ. Res. Public Health 2019, 16, 1438. [Google Scholar] [CrossRef] [Green Version]
- Skinner, M.K.; Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Haque, M.; Nilsson, E.E. Ancestral dichlorodiphenyltrichloroethane (DDT) exposure promotes epigenetic transgenerational inheritance of obesity. BMC Med. 2013, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Jandacek, R.J.; Rider, T.; Yang, Q.; Woollett, L.A.; Tso, P. Lymphatic and portal vein absorption of organochlorine compounds in rats. Am. J. Physiol.—Gastrointest. Liver Physiol. 2009, 296, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Timoumi, R.; Amara, I.; Neffati, F.; Najjar, M.F.; Golli-Bennour, E.E.; Bacha, H.; Abid-Essefi, S. Acute triflumuron exposure induces oxidative stress responses in liver and kidney of Balb/C mice. Environ. Sci. Pollut. Res. 2018, 26, 3723–3730. [Google Scholar] [CrossRef]
- Smith, A.G. Toxicology of organochlorine insecticide. In Pesticide Toxicology and International Regulation; Mars, T.C., Ballantyne, B., Eds.; Wiley: Somerset, NJ, USA, 2004; pp. 27–87. [Google Scholar] [CrossRef]
- Huber, P.C.; Almeida, W.P.; Fátima, A. Glutationa e enzimas relacionadas: Papel biológico e importância em processos patológicos. Química Nova 2008, 31, 1170–1179. [Google Scholar] [CrossRef] [Green Version]
- Chehade, L.; Khouri, H.; Malatier-Ségard, J.; Caron, A.; Mauger, J.F.; Chapados, N.A.; Aguer, A. Acute exposure to environmentally relevant levels of DDT alters muscle mitochondrial function in vivo in rats but not in vitro in L6 myotubes: A pilot study. Toxicol. Rep. 2022, 5, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Cell biology of ischemia/reperfusion injury. Int. Rev. Cell. Mol. Biol. 2012, 298, 229–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Role of metabolic H2O2 generation: Redox signaling and oxidative stress. J. Biol. Chem. 2014, 289, 8735–8741. [Google Scholar] [CrossRef] [Green Version]
- Lefranc, C.; Friederich-Persson, M.; Palacios-Ramirez, R.; Nguyen Dinh Cat, A. Mitochondrial oxidative stress in obesity: Role of the mineralocorticoid receptor. J. Endocrinol. 2018, 238, 143–159. [Google Scholar] [CrossRef]
- Migliaccio, V.; Lionetti, L.; Putti, R.; Scudiero, R. Exposure to dichlorodiphenyldichloroethylene (DDE) and metallothionein levels in rats fed with normocaloric or high-fat diet: A review. Int. J. Mol. Sci. 2020, 10, 1903. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, N.G.; Morgado, R.G.; Amaro, A.; Machado, A.L.; Soares, A.M.; Loureiro, S. The effects of temperature, soil moisture and UV radiation on biomarkers and energy reserves of the isopod Porcellionides pruinosus. Appl. Soil Ecol. 2016, 107, 224–236. [Google Scholar] [CrossRef]
- Marouani, N.; Hallegue, D.; Sakly, M.; Benkhalifa, M.; Rhouma, K.B.; Tebourb, O. Involvement of oxidative stress in the mechanism of p, p’-DDT-induced nephrotoxicity in adult rats. Gen. Physiol. Biophys. 2017, 36, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Siddarth, M.; Datta, S.K.; Mustafa, M.D.; Ahmed, R.S.; Banerjee, B.D.; Kalra, O.M.P.; Tripathi, A.K. Increased level of organochlorine pesticides in chronic kidney disease patients of unknown etiology: Role of GSTM1/GSTT1 polymorphism. Chemosphere 2014, 96, 174–179. [Google Scholar] [CrossRef]
- Hall, J.E. Tratado de Fisiologia Médica, 12th ed; Elsevier: Rio de Janeiro, RJ, Brazil, 2011. [Google Scholar]
- Hawkins, B.T.; Davis, T.P. The blood-brain barrier/neurovascular unit in Health and disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef]
- Gupta, A.; Agarwal, R.; Shukla, G.S. Functional impairment of blood-brain barrier following pesticide exposure during early development in rats. Hum. Exp. Toxicol. 1999, 18, 174–179. [Google Scholar]
- Forster, M.J.; Dubey, A.; Dawson, K.M.; Stutts, W.A.; Lal, H.; Sohal, R.S. Age-related losses of cognitive function and motor skills in mice are associated with oxidative protein damage in the brain. Proc. Natl. Acad. Sci. USA 1996, 93, 4765–4769. [Google Scholar] [CrossRef] [Green Version]
- Falco, A.D.; Cukierman, D.S.; Hauser-Davis, R.A.; Rey, N.A. Doença de Alzheimer: Hipótesesetiológicas e perspectivas de tratamento. Química Nova 2016, 39, 63–80. [Google Scholar] [CrossRef]
- Ishikawa, T.; Graham, J.L.; Stanhope, K.L.; Havel, P.J.; La Merril, M.A. Effect of DDT exposure on lipids and energy balance in obese Sprague-Dawley rats before and after weight loss. Toxicol. Rep. 2015, 2, 990–995. [Google Scholar] [CrossRef] [Green Version]
- Romero, C.E.M.; Zanesco, A. O papel dos hormônios leptina e grelina na gênese da obesidade. Rev. De Nutr. 2006, 19, 85–91. [Google Scholar] [CrossRef] [Green Version]
- La Merrill, M.; Karey, E.; Moshier, E.; Lindtner, C.; La Frano, M.R.; Newman, J.W.; Buettner, C. Perinatal exposure of mice to the pesticide DDT impairs energy expenditure and metabolism in adult female offspring. PLoS ONE 2014, 9, e103337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.-J.; Zhang, Y.; Gerhard, M.; Mejias-Luque, R.; Zhang, L.; Vieth, M.; Ma, J.-L.; Bajbouj, M.; Suchanek, S.; Liu, W.-D.; et al. Association between gut microbiota and Helicobacter pylori-related gastric lesions in a high-risk population of gastric cancer. Front. Cell. Infect. Microbiol. 2018, 8, 202–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhao, F.; Xu, Y.; Qiu, J.; Qian, Y. Gut Flora-Mediated Metabolic Health, the Risk Produced by Dietary Exposure to Acetamiprid and Tebuconazole. Foods 2021, 10, 835–853. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Sun, Q.; Yue, Y.; Yoon, K.S.; Whang, K.Y.; Marshall Clark, J.; Park, Y. 4,4’-Dichlorodiphenyltrichloroethane (DDT) and 4,4’-dichlorodiphenyldichloroethylene (DDE) promote adipogenesis in 3T3-L1 adipocyte cell culture. Pestic. Biochem. Physiol. 2016, 131, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Ballantyne, C.M. Skeletal muscle inflammation and insulin resistance in obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef]
- Kumar, A.; Davuluri, G.; Welch, N.; Kim, A.; Gangadhariah, M.; Allawy, A.; Priyadarshini, A.; McMullen, M.R.; Sandlers, Y.; Willard, B.; et al. Oxidative stress mediates ethanol-induced skeletal muscle mitochondrial dysfunction and dysregulated protein synthesis and autophagy. Free Radic. Biol. Med. 2019, 145, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Chevrier, J.; Rosas, L.G.; Anderson, H.A.; Bornman, M.S.; Bouwman, H.; Chen, A.; Cohn, B.A.; Jager, C.; Henshel, D.S.; et al. The Pine River statement: Human health consequences of DDT use. Environ. Health Perspect. 2009, 117, 1359–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valvi, D.; Mendez, M.A.; Martinez, D.; Grimalt, J.O.; Torrent, M.; Sunyer, J.; Vrijheid, M. Prenatal concentrations of polychlorinated biphenyls, DDE, and DDT and overweight in children: A prospective birth cohort study. Environ. Health Perspect. 2012, 120, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Evaluation | Control | Exposure |
|---|---|---|
| Total body weight gain (g) | 288.75 ± 3.30 | 313.375 ± 10.01 * |
| Total food consumption (g) | 1436.00 ± 16.62 | 1504.62 ± 30.65 |
| Body weight gain rate | 6.65 ± 0.23 | 6.69 ± 0.21 |
| Feeding efficiency | 0.20 ± 0.003 | 0.21 ± 0.003 |
| Body weight gain per caloric intake | 0.053 ± 0.001 | 0.055 ± 0.001 |
| Control | Exposure | p-Value | |
|---|---|---|---|
| Hemoglobin concentration (g/dL) | 13.79 ± 0.60 | 14.75 ± 0.76 | 0.057 |
| Microhematocrit (%) | 41.38 ± 1.81 | 44.25 ± 2.27 | 0.151 |
| Red blood cells (106/mm3) | 8.21 ± 0.59 | 9.50 ± 0.61 | 0.1537 |
| White blood cells (103/mm3) | 5.67 ± 0.50 | 5.86 ± 0.53 | 0.8015 |
| Stick neutrophil (%) | 0.63 ± 0.37 | 0.25 ± 0.16 | 0.5613 |
| Segmented neutrophil (%) | 18.88 ± 2.53 | 19.75 ± 2.50 | 0.8095 |
| Lymphocyte (%) | 79.75 ± 2.72 | 78.75 ± 2.40 | 0.7871 |
| Monocyte (%) | 0.75 ± 0.25 | 1.25 ± 0.41 | 0.317 |
| Eosinophil (%) | 0 | 0 | - |
| Basophil (%) | 0 | 0 | - |
| Mean cell volume (fL) | 52.34 ± 4.39 | 48.37 ± 4.15 | 0.5223 |
| Mean cell hemoglobin (pg) | 17.45 ± 1.46 | 16.12 ± 1.38 | 0.5223 |
| Mean cell hemoglobin conc. (g/dL) | 33.33 ± 0.00 | 33.33 ± 0.00 | - |
| Tissue | Analysis | Control | Exposure | p-Value |
|---|---|---|---|---|
| Liver | SOD | 10.97 ± 1.69 | 12.53 ± 1.60 | 0.513 |
| GR | 29.44 ± 2.30 | 34.20 ± 1.26 | 0.090 | |
| GST | 116.30 ± 10.76 | 142.11 ± 5.36 * | 0.049 | |
| GPx | 5.79 ± 1.45 | 10.13 ± 2.52 | 0.158 | |
| LPO | 22.19 ± 1.93 | 19.87 ± 3.92 | 0.604 | |
| ALT | 23.64 ± 2.83 | 35.77 ± 4.02 * | 0.026 | |
| AST | 84.89 ± 14.90 | 119.48 ± 16.69 | 0.145 | |
| Muscle | SOD | 6.49 ± 0.37 | 5.14 ± 0.40 * | 0.026 |
| GR | 6.82 ± 0.36 | 6.55 ± 0.36 | 0.612 | |
| GST | 23.78 ± 0.49 | 22.35 ± 0.64 | 0.097 | |
| GPX | 83.07 ± 8.95 | 161.04 ± 7.20 * | <0.0001 | |
| LPO | 5.31 ± 0.64 | 1.77 ± 0.53 * | <0.001 | |
| Kidney | SOD | 12.29 ± 0.78 | 16.63 ± 1.11 * | 0.006 |
| GR | 29.68 ± 2.35 | 28.62 ± 3.02 | 0.786 | |
| GST | 37.14 ± 3.83 | 41.55 ± 2.51 | 0.352 | |
| GPx | 38.28 ± 7.85 | 59.01 ± 9.85 | 0.122 | |
| LPO | 1.45 ± 0.56 | 2.48 ± 0.44 | 0.073 £ | |
| Brain | SOD | 9.72 ± 0.65 | 9.25 ± 0.59 | 0.603 |
| GR | 14.39 ± 2.56 | 11.89 ± 1.13 | 0.396 | |
| GST | 31.19 ± 2.89 | 29.51 ± 0.62 | 0.878 £ | |
| GPx | 120.23 ± 4.18 | 151.90 ± 3.00 * | <0.0001 | |
| LPO | 21.52 ± 2.66 | 28.66 ± 2.80 | 0.086 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolfart, J.C.; Theodoro, J.L.; Silva, F.C.; de Oliveira, C.M.R.; Ferreira, N.G.C.; Bittencourt Guimarães, A.T. Metabolic Consequences of the Water We Drink: A Study Based on Field Evidence and Animal Model Experimentation. Toxics 2023, 11, 315. https://doi.org/10.3390/toxics11040315
Wolfart JC, Theodoro JL, Silva FC, de Oliveira CMR, Ferreira NGC, Bittencourt Guimarães AT. Metabolic Consequences of the Water We Drink: A Study Based on Field Evidence and Animal Model Experimentation. Toxics. 2023; 11(4):315. https://doi.org/10.3390/toxics11040315
Chicago/Turabian StyleWolfart, Janaína Caroline, João Lucas Theodoro, Fernanda Coleraus Silva, Cíntia Mara Ribas de Oliveira, Nuno G. C. Ferreira, and Ana Tereza Bittencourt Guimarães. 2023. "Metabolic Consequences of the Water We Drink: A Study Based on Field Evidence and Animal Model Experimentation" Toxics 11, no. 4: 315. https://doi.org/10.3390/toxics11040315
APA StyleWolfart, J. C., Theodoro, J. L., Silva, F. C., de Oliveira, C. M. R., Ferreira, N. G. C., & Bittencourt Guimarães, A. T. (2023). Metabolic Consequences of the Water We Drink: A Study Based on Field Evidence and Animal Model Experimentation. Toxics, 11(4), 315. https://doi.org/10.3390/toxics11040315