



Per- and Polyfluoroalkyl Substances: Impacts on Morphology, Behavior and Lipid Levels in Zebrafish Embryos



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Analytical Chemistry
2.2. Zebrafish Husbandry and Exposure
2.3. Behavior Data Collection and Analysis
2.4. Lipid Extraction and Analysis
2.5. Statistical Analysis
3. Results
3.1. Analytical Chemistry
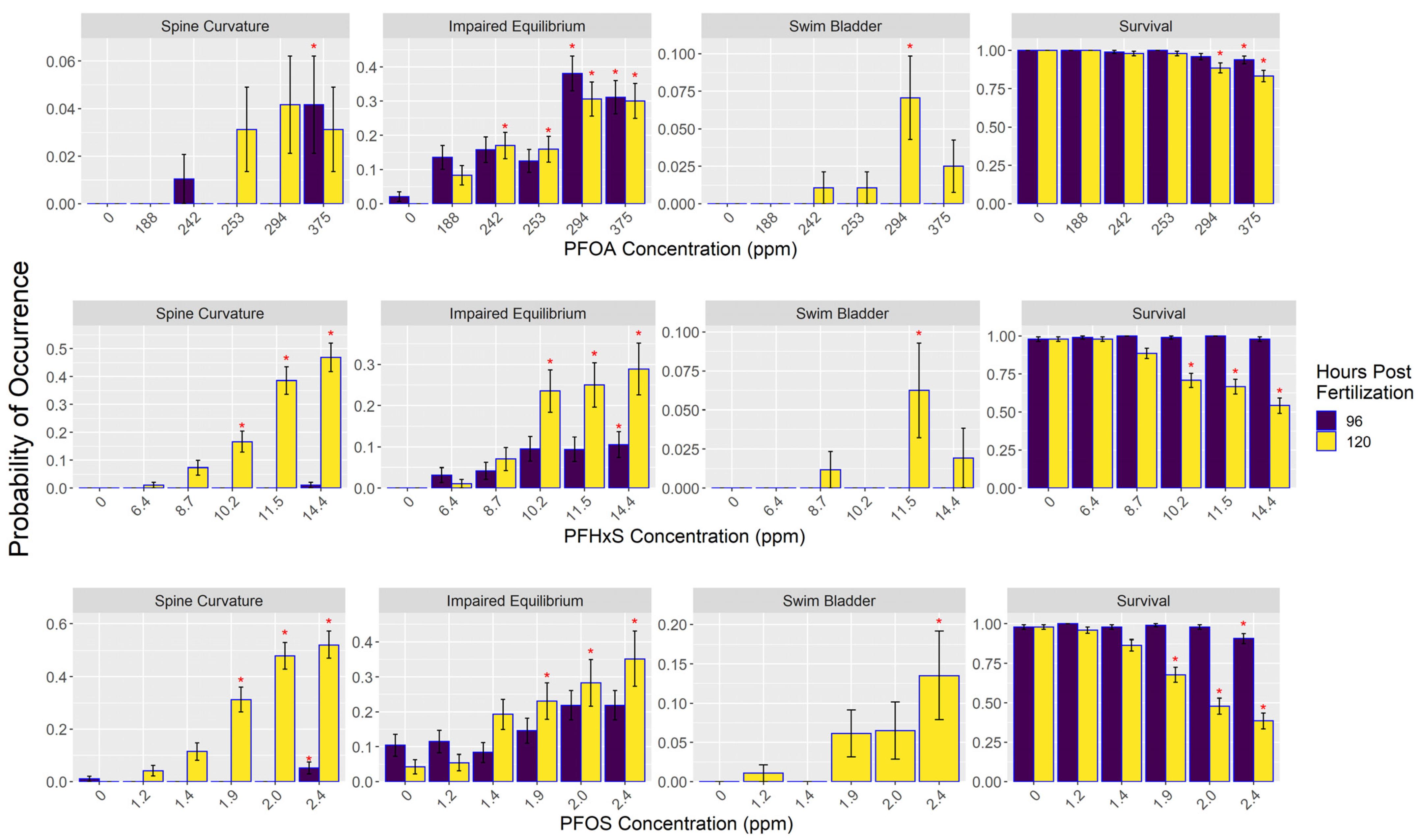
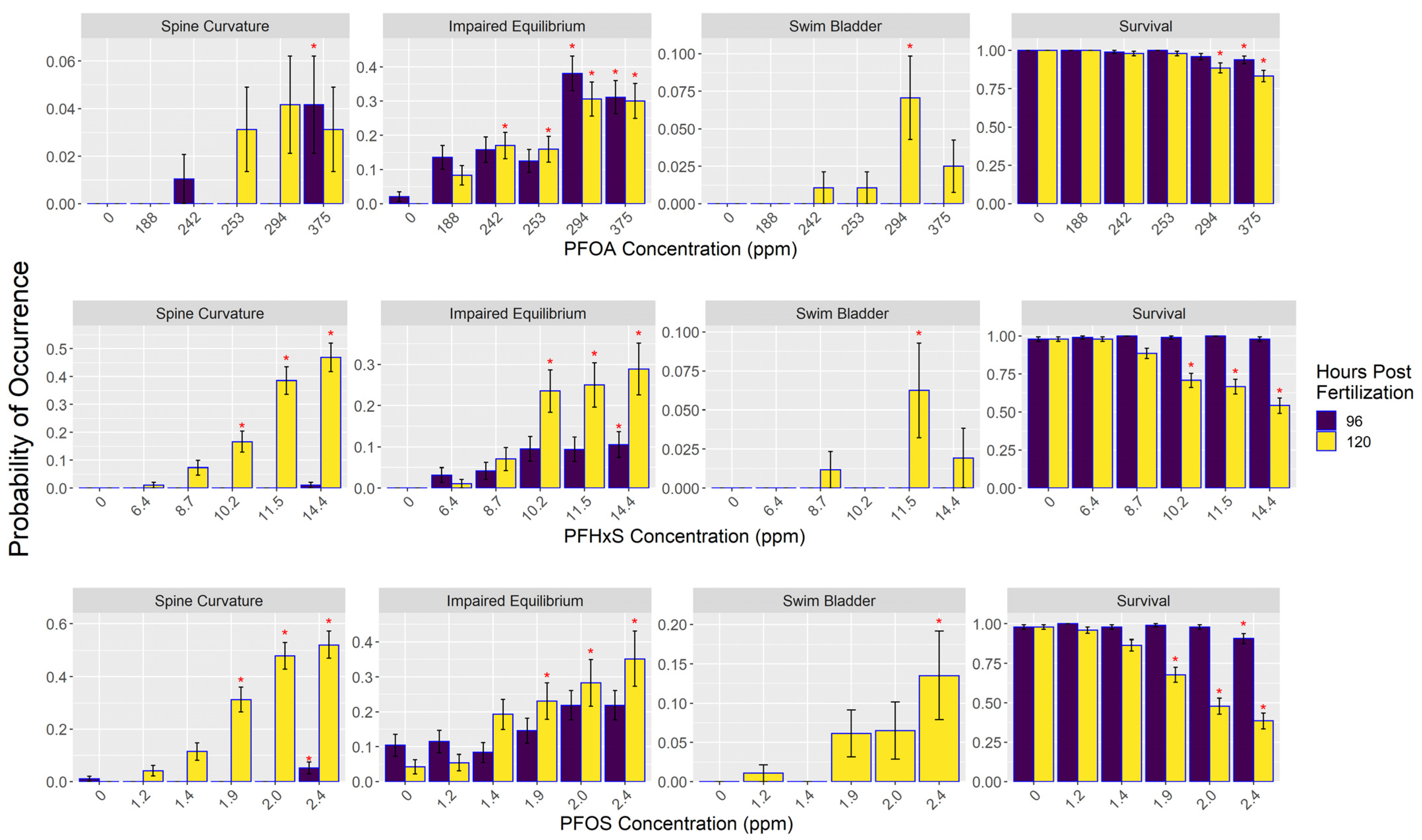
3.2. Morphometric Results
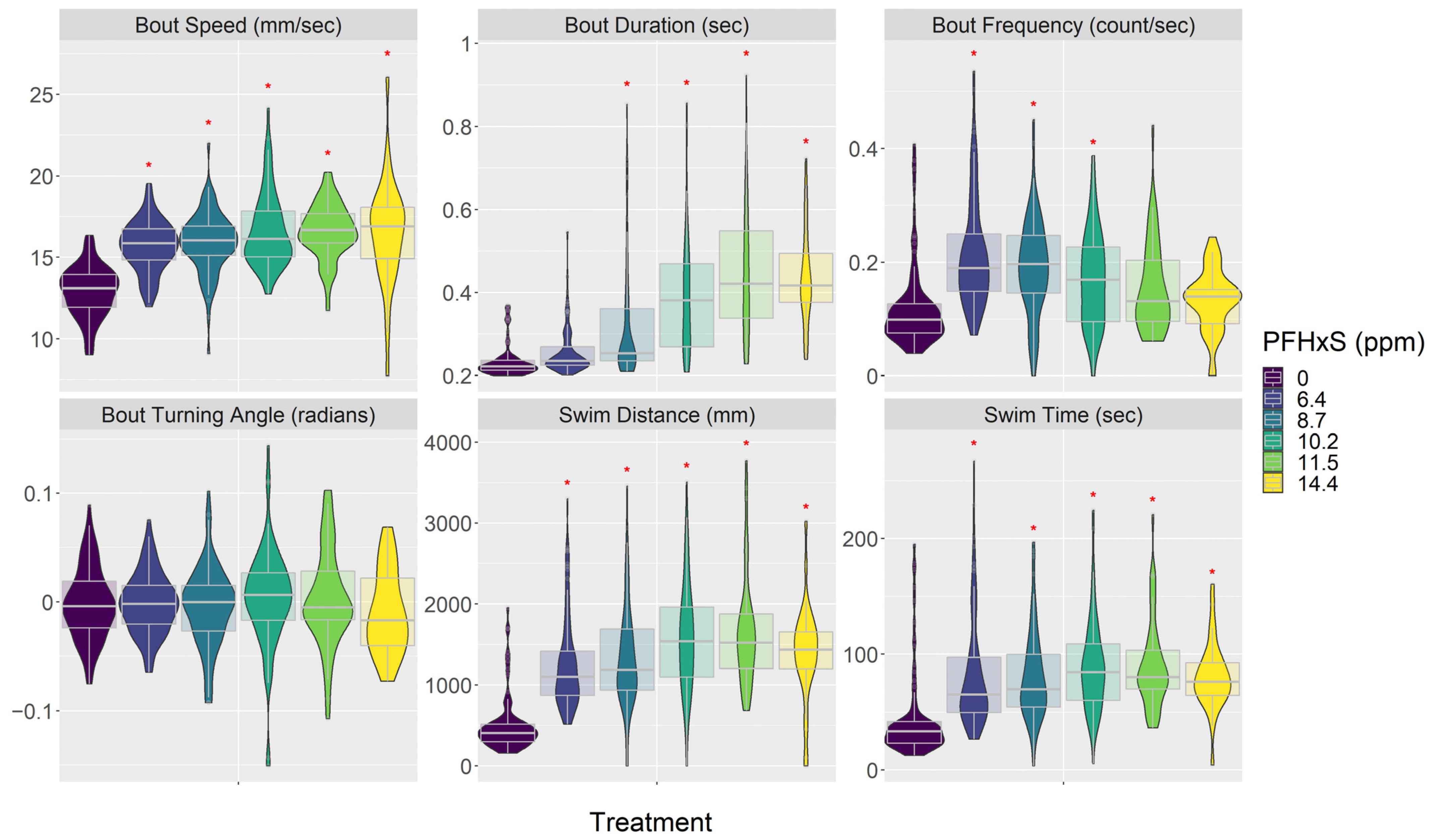
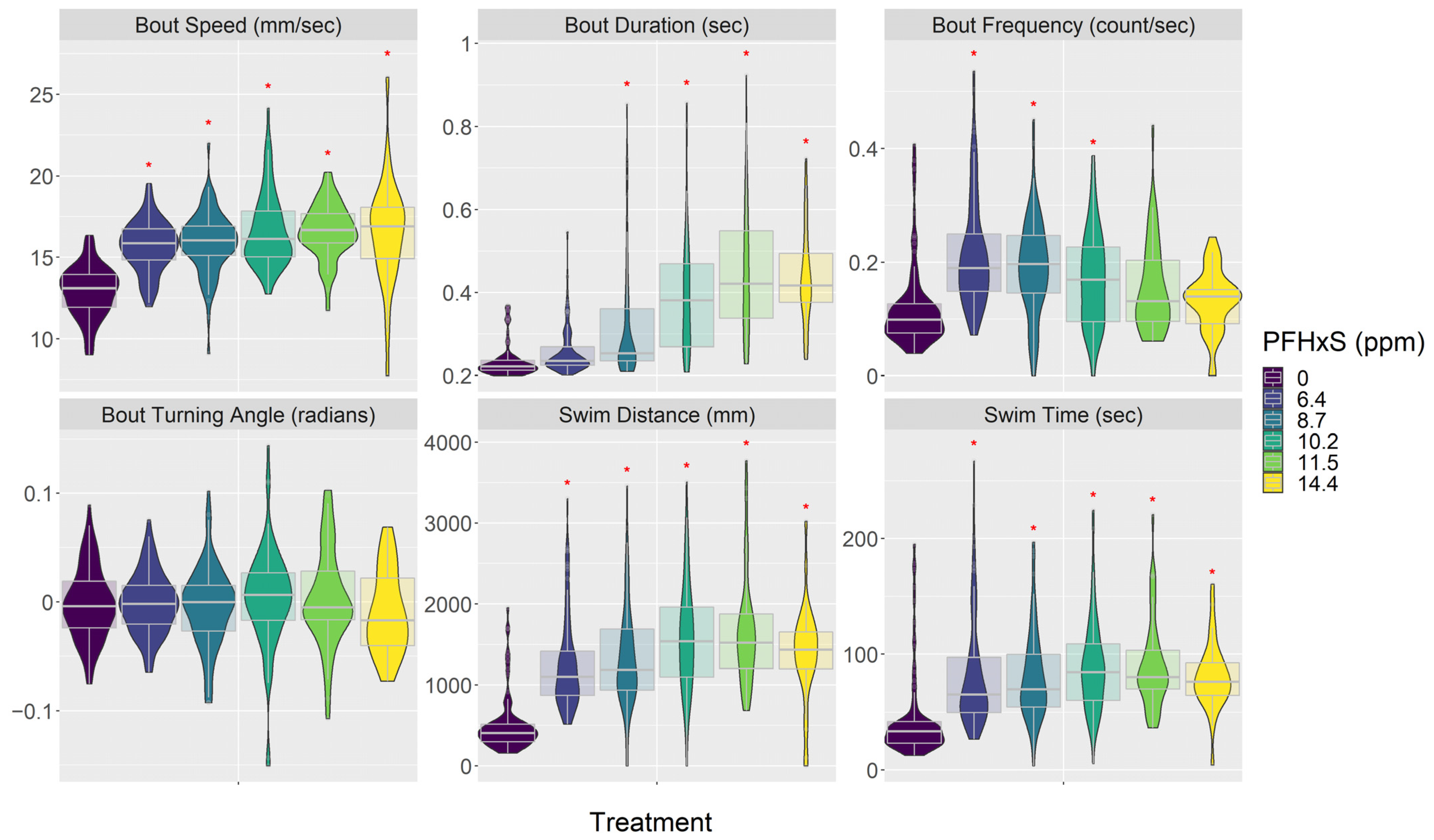
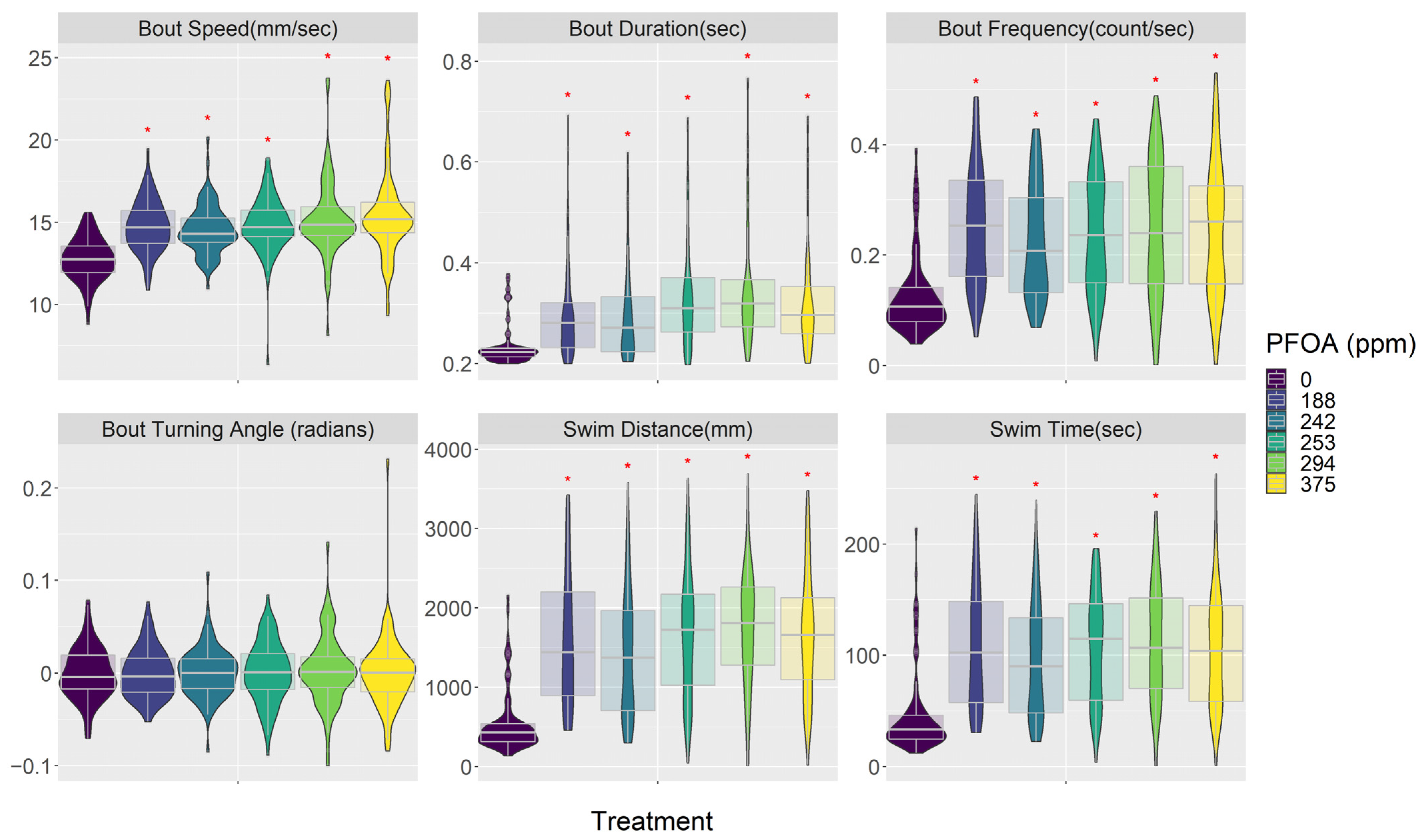
3.3. Behavior (Visual Motor Response)
3.4. Lipidomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Annunziato, K.M.; Doherty, J.; Lee, J.; Clark, J.M.; Liang, W.; Clark, C.W.; Nguyen, M.; Roy, M.A.; Timme-Laragy, A.R. Chemical Characterization of a Legacy Aqueous Film-Forming Foam Sample and Developmental Toxicity in Zebrafish (Danio rerio). Environ. Health Perspect. 2020, 128, 097006. [Google Scholar] [CrossRef]
- DeWitt, J.C.; Blossom, S.J.; Schaider, L.A. Exposure to per-fluoroalkyl and polyfluoroalkyl substances leads to immunotoxicity: Epidemiological and toxicological evidence. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 148–156. [Google Scholar] [CrossRef]
- Glüge, J.; Scheringer, M.; Cousins, I.T.; DeWitt, J.C.; Goldenman, G.; Herzke, D.; Lohmann, R.; Ng, C.A.; Trier, X.; Wang, Z. An overview of the uses of per- and polyfluoroalkyl substances (PFAS). Environ. Sci. Process. Impacts 2020, 22, 2345–2373. [Google Scholar] [CrossRef]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl Acids: A Review of Monitoring and Toxicological Findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [PubMed]
- Sima, M.W.; Jaffé, P.R. A critical review of modeling Poly- and Perfluoroalkyl Substances (PFAS) in the soil-water environment. Sci. Total Environ. 2021, 757, 143793. [Google Scholar] [CrossRef] [PubMed]
- Ankley, G.T.; Cureton, P.; Hoke, R.A.; Houde, M.; Kumar, A.; Kurias, J.; Lanno, R.; McCarthy, C.; Newsted, J.; Salice, C.J.; et al. Assessing the Ecological Risks of Per- and Polyfluoroalkyl Substances: Current State-of-the Science and a Proposed Path Forward. Environ. Toxicol. Chem. 2021, 40, 564–605. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.C.; Ikonomou, M.G.; Blair, J.D.; Surridge, B.; Hoover, D.; Grace, R.; Gobas, F.A. Perfluoroalkyl Contaminants in an Arctic Marine Food Web: Trophic Magnification and Wildlife Exposure. Environ. Sci. Technol. 2009, 43, 4037–4043. [Google Scholar] [CrossRef] [PubMed]
- Miranda, D.A.; Benskin, J.P.; Awad, R.; Lepoint, G.; Leonel, J.; Hatje, V. Bioaccumulation of Per- and polyfluoroalkyl substances (PFASs) in a tropical estuarine food web. Sci. Total Environ. 2021, 754, 142146. [Google Scholar] [CrossRef] [PubMed]
- Miranda, D.D.A.; Peaslee, G.F.; Zachritz, A.M.; Lamberti, G.A. A worldwide evaluation of trophic magnification of per- and polyfluoroalkyl substances in aquatic ecosystems. Integr. Environ. Assess. Manag. 2022, 18, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Savoca, D.; Pace, A. Bioaccumulation, Biodistribution, Toxicology and Biomonitoring of Organofluorine Compounds in Aquatic Organisms. Int. J. Mol. Sci. 2021, 22, 6276. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological profile for Perfluoroalkyls; U.S. Department of Health and Human Services, Public Health Service: Atlanta, GA, USA, 2021. [Google Scholar]
- Bell, E.M.; De Guise, S.; McCutcheon, J.R.; Lei, Y.; Levin, M.; Li, B.; Rusling, J.F.; Lawrence, D.A.; Cavallari, J.M.; O’Connell, C.; et al. Exposure, health effects, sensing, and remediation of the emerging PFAS contaminants—Scientific challenges and potential research directions. Sci. Total Environ. 2021, 780, 146399. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Hwang, G.W.; Naganuma, A.; Satoh, M. Methylmercury toxic mechanism related to protein degradation and chemokine transcription. Environ. Health Prev. Med. 2020, 25, 30. [Google Scholar] [CrossRef]
- Sunderland, E.M.; Hu, X.C.; Dassuncao, C.; Tokranov, A.K.; Wagner, C.C.; Allen, J.G. A review of the pathways of human exposure to poly- and perfluoroalkyl substances (PFASs) and present understanding of health effects. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 131–147. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, J.; Liu, S.; Wang, C.; Jin, Y.; Lai, H.; Tu, W. Aberrant hepatic lipis metabolism associated with gut microbiota dysbiosis triggers hepatotoxicity of novel PFOS alternatives in adult zebrafish. Environ. Int. 2022, 166, 107351. [Google Scholar] [CrossRef]
- Bambino, K.; Chu, J. Zebrafish in Toxicology and Environmental Health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar] [CrossRef]
- Braunbeck, T.; Kais, B.; Lammer, E.; Otte, J.; Schneider, K.; Stengel, D.; Strecker, R. The fish embryo test (FET): Origin, applications, and future. Environ. Sci. Pollut. Res. 2015, 22, 16247–16261. [Google Scholar] [CrossRef]
- Hahn, M.E.; Sadler, K.C. Casting a wide net: Use of diverse model organisms to advance toxicology. Dis. Models Mech. 2020, 13, dmm043844. [Google Scholar] [CrossRef] [PubMed]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2013. [Google Scholar] [CrossRef]
- Tanguay, R.L. The Rise of Zebrafish as a Model for Toxicology. Toxicol. Sci. 2018, 163, 3–4. [Google Scholar] [CrossRef] [PubMed]
- von Hellfeld, R.; Brotzmann, K.; Baumann, L.; Strecker, R.; Braunbeck, T. Adverse effects in the fish embryo acute toxicity (FET) test: A catalogue of unspecific morphological changes versus more specific effects in zebrafish (Danio rerio) embryos. Environ. Sci. Eur. 2020, 32, 122. [Google Scholar] [CrossRef]
- Annunziato, K.M.; Jantzen, C.E.; Gronske, M.C.; Cooper, K.R. Subtle morphometric, behavioral and gene expression effects in larval zebrafish exposed to PFHxA, PFHxS and 6:2 FTOH. Aquat. Toxicol. 2019, 208, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Gaballah, S.; Swank, A.; Sobus, J.R.; Howey, X.M.; Schmid, J.; Catron, T.; McCord, J.; Hines, E.; Strynar, M.; Tal, T. Evaluation of Developmental Toxicity, Developmental Neurotoxicity, and Tissue Dose in Zebrafish Exposed to GenX and Other PFAS. Environ. Health Perspect. 2020, 128, 047005. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, S.; Liu, X.; Lu, S.; Wu, Q.; Xie, P. Evaluation of the acute toxicity and neurodevelopmental inhibition of perfluorohexanoic acid (PFHxA) in zebrafish embryos. Ecotoxicol. Environ. Saf. 2021, 225, 112733. [Google Scholar] [CrossRef]
- Huang, H.; Huang, C.; Wang, L.; Ye, X.; Bai, C.; Simonich, M.T.; Tanguay, R.L.; Dong, Q. Toxicity, uptake kinetics and behavior assessment in zebrafish embryos following exposure to perfluorooctanesulphonicacid (PFOS). Aquat. Toxicol. 2010, 98, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Jantzen, C.E.; Annunziato, K.A.; Bugel, S.M.; Cooper, K.R. PFOS, PFNA, and PFOA sub-lethal exposure to embryonic zebrafish have different toxicity profiles in terms of morphometrics, behavior and gene expression. Aquat. Toxicol. 2016, 175, 160–170. [Google Scholar] [CrossRef]
- Lee, H.; Sung, E.J.; Seo, S.; Min, E.K.; Lee, J.Y.; Shim, I.; Kim, P.; Kim, T.Y.; Lee, S.; Kim, K.T. Integrated multi-omics analysis reveals the underlying molecular mechanism for developmental neurotoxicity of perfluorooctanesulfonic acid in zebrafish. Environ. Int. 2021, 157, 106802. [Google Scholar] [CrossRef]
- Liu, X.; Liu, S.; Qiu, W.; Magnuson, J.T.; Liu, Z.; Yang, G.; Chen, H.; Li, Y.; Xu, X.; Zheng, C. Cardiotoxicity of PFOA, PFOS, and PFOSA in early life stage zebrafish: Molecular changes to behavioral-level response. Sustain. Horiz. 2022, 3, 100027. [Google Scholar] [CrossRef]
- Mylroie, J.E.; Wilbanks, M.S.; Kimble, A.N.; To, K.T.; Cox, C.S.; McLeod, S.J.; Gust, K.A.; Moore, D.W.; Perkins, E.J.; Garcia-Reyero, N. Perfluorooctanesulfonic Acid–Induced Toxicity on Zebrafish Embryos in the Presence or Absence of the Chorion. Environ. Toxicol. Chem. 2021, 40, 780–791. [Google Scholar] [CrossRef]
- Truong, L.; Rericha, Y.; Thunga, P.; Marvel, S.; Wallis, D.; Simonich, M.T.; Field, J.A.; Cao, D.; Reif, D.M.; Tanguay, R.L. Systematic developmental toxicity assessment of a structurally diverse library of PFAS in zebrafish. J. Hazard. Mater. 2022, 431, 128615. [Google Scholar] [CrossRef] [PubMed]
- Ulhaq, M.; Carlsson, G.; Örn, S.; Norrgren, L. Comparison of developmental toxicity of seven perfluoroalkyl acids to zebrafish embryos. Environ. Toxicol. Pharmacol. 2013, 36, 423–426. [Google Scholar] [CrossRef]
- Vogs, C.; Johanson, G.; Näslund, M.; Wulff, S.; Sjödin, M.; Hellstrandh, M.; Lindberg, J.; Wincent, E. Toxicokinetics of Perfluorinated Alkyl Acids Influences Their Toxic Potency in the Zebrafish Embryo (Danio rerio). Environ. Sci. Technol. 2019, 53, 3898–3907. [Google Scholar] [CrossRef]
- Wasel, O.; Thompson, K.M.; Gao, Y.; Godfrey, A.E.; Gao, J.; Mahapatra, C.T.; Lee, L.S.; Sepúlveda, M.S.; Freeman, J.L. Comparison of zebrafish in vitro and in vivo developmental toxicity assessments of perfluoroalkyl acids (PFAAs). J. Toxicol. Environ. Health Part A 2021, 84, 125–136. [Google Scholar] [CrossRef]
- Wasel, O.; Thompson, K.M.; Freeman, J.L. Assessment of unique behavioral, morphological, and molecular alterations in the comparative developmental toxicity profiles of PFOA, PFHxA, and PFBA using the zebrafish model system. Environ. Int. 2022, 170, 107642. [Google Scholar] [CrossRef]
- Zheng, X.-M.; Liu, H.L.; Shi, W.; Wei, S.; Giesy, J.P.; Yu, H.-X. Effects of perfluorinated compounds on development of zebrafish embryos. Environ. Sci. Pollut. Res. 2012, 19, 2498–2505. [Google Scholar] [CrossRef]
- Hagenaars, A.; Stinckens, E.; Vergauwen, L.; Bervoets, L.; Knapen, D. PFOS affects posterior swim bladder chamber inflation and swimming performance of zebrafish larvae. Aquat. Toxicol. 2014, 157, 225–235. [Google Scholar] [CrossRef]
- Khezri, A.; Fraser, T.W.; Nourizadeh-Lillabadi, R.; Kamstra, J.H.; Berg, V.; Zimmer, K.E.; Ropstad, E. A Mixture of Persistent Organic Pollutants and Perfluorooctanesulfonic Acid Induces Similar Behavioural Responses, but Different Gene Expression Profiles in Zebrafish Larvae. Int. J. Mol. Sci. 2017, 18, 291. [Google Scholar] [CrossRef]
- Menger, F.; Pohl, J.; Ahrens, L.; Carlsson, G.; Örn, S. Behavioural effects and bioconcentration of per- and polyfluoroalkyl substances (PFASs) in zebrafish (Danio rerio) embryos. Chemosphere 2020, 245, 125573. [Google Scholar] [CrossRef]
- Rericha, Y.; Cao, D.; Truong, L.; Simonich, M.; Field, J.A.; Tanguay, R.L. Behavior Effects of Structurally Diverse Per- and Polyfluoroalkyl Substances in Zebrafish. Chem. Res. Toxicol. 2021, 34, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Lv, S.; Nie, S.; Liu, J.; Tong, S.; Kang, N.; Xiao, Y.; Dong, Q.; Huang, C.; Yang, D. Chronic perfluorooctane sulfonate (PFOS) exposure induces hepatic steatosis in zebrafish. Aquat. Toxicol. 2016, 176, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Fai Tse, W.K.; Li, J.W.; Tse, A.C.K.; Chan, T.F.; Ho, J.C.H.; Wu, R.S.S.; Wong, C.K.C.; Lai, K.P. Fatty liver disease induced by perfluorooctane sulfonate: Novel insight from transcriptome analysis. Chemosphere 2016, 159, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Gebreab, K.Y.; Eeza, M.N.; Bai, T.; Zuberi, Z.; Matysik, J.; O’Shea, K.E.; Alia, A.; Berry, J.P. Comparative toxicometabolomics of perfluorooctanoic acid (PFOA) and next-generation perfluoroalkyl substances. Environ. Pollut. 2020, 265, 114928. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, Y.; Wang, Q.; Yi, J.; Lai, H.; Sun, L.; Mennigen, J.A.; Tu, W. Concentration-dependent toxicokinetics of novel PFOS alternatives and their chronic combined toxicity in adult zebrafish. Sci. Total Environ. 2022, 839, 156388. [Google Scholar] [CrossRef]
- Sen, P.; Qadri, S.; Luukkonen, P.K.; Ragnarsdottir, O.; McGlinchey, A.; Jäntti, S.; Juuti, A.; Arola, J.; Schlezinger, J.J.; Webster, T.F.; et al. Exposure to environmental contaminants is associated with altered hepatic lipid metabolism in non-alcoholic fatty liver disease. J. Hepatol. 2022, 76, 283–293. [Google Scholar] [CrossRef]
- Tan, Q.; Zhang, M.; Geng, L.; Xia, Z.; Li, C.; Usman, M.; Du, Y.; Wei, L.; Bi, H. Hormesis of methylmercury-human serum albumin conjugate on N9 microglia via ERK/MAPKs and STAT3 signaling pathways. Toxicol. Appl. Pharmacol. 2019, 362, 59–66. [Google Scholar] [CrossRef]
- Beale, D.J.; Sinclair, G.M.; Shah, R.; Paten, A.M.; Kumar, A.; Long, S.M.; Vardy, S.; Jones, O.A. A review of omics-based PFAS exposure studies reveals common biochemcial response pathways. Sci. Total Environ. 2022, 845, 157255. [Google Scholar] [CrossRef]
- Haimbaugh, A.; Wu, C.C.; Akemann, C.; Meyer, D.N.; Connell, M.; Abdi, M.; Khalaf, A.; Johnson, D.; Baker, T.R. Multi- and transgenerational effects of developmental exposure to environmental levels of PFAS and PFAS mixture in zebrafish (Danio rerio). Toxics 2022, 10, 334. [Google Scholar] [CrossRef]
- Lee, J.W.; Choi, K.; Park, K.; Seong, C.; Do Yu, S.; Kim, P. Adverse effects of perfluoroalkyl acids on fish and other aquatic organisms: A review. Sci. Total Environ. 2020, 707, 135334. [Google Scholar] [CrossRef]
- Martínez, R.; Navarro-Martín, L.; Luccarelli, C.; Codina, A.E.; Raldúa, D.; Barata, C.; Tauler, R.; Piña, B. Unravelling the mechanisms of PFOS toxicity by combining morphological and transcriptomic analyses in zebrafish embryos. Sci. Total Environ. 2019, 674, 462–471. [Google Scholar] [CrossRef]
- Ortiz-Villanueva, E.; Jaumot, J.; Martínez, R.; Navarro-Martín, L.; Piña, B.; Tauler, R. Assessment of endocrine disruptors effects on zebrafish (Danio rerio) embryos by untargeted LC-HRMS metabolomic analysis. Sci. Total Environ. 2018, 635, 156–166. [Google Scholar] [CrossRef]
- Xu, M.; Legradi, J.; Leonards, P. Using comprehensive lipid profiling to study effects of PFHxS during different stages of early zebrafish development. Sci. Total Environ. 2022, 808, 151739. [Google Scholar] [CrossRef] [PubMed]
- Sant, K.E.; Sinno, P.P.; Jacobs, H.M.; Timme-Laragy, A.R. Nrf2a modulates the embryonic antioxidant response to perfluorooctanesulfonic acid (PFOS) in the zebrafish, Danio rerio. Aquat. Toxicol. 2018, 198, 92–102. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency (U.S. EPA). Drinking Water Health Advisory for Perfluorooctanoic Acid (PFOA); EPA Document Number: 822-R-16-005; Office of Water (4304T), Health and Ecological Criteria Division EPA: Washington, DC, USA, 2016. Available online: https://www.epa.gov/sites/default/files/2016-05/documents/pfoa_health_advisory_final-plain.pdf (accessed on 20 February 2024).
- Campbell, T.Y.; Vecitis, C.D.; Mader, B.T.; Hoffmann, M.R. Perfluorinated surfactant chain-length effects on sonochemical kinetics. J. Phys. Chem. A 2009, 113, 9834–9842. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 74483, Perfluorooctanesulfonic Acid. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Perfluorooctanesulfonic-acid (accessed on 20 February 2024).
- Varga, Z.M. Aquaculture, husbandry, and shipping at the Zebrafish International Resource Center. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2016; pp. 509–534. [Google Scholar] [CrossRef]
- Varga, Z.M.; Murray, K.N. Health monitoring and disease prevention at the Zebrafish International Resource Center. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2016; pp. 535–551. [Google Scholar] [CrossRef]
- Emran, F.; Rihel, J.; Dowling, J.E. A Behavioral Assay to Measure Responsiveness of Zebrafish to Changes in Light Intensities. J. Vis. Exp. 2008, 20, e923. [Google Scholar] [CrossRef]
- Albers, J.L. Effects of Neurotoxic Contaminants on Larval Fish, from Genes and Behavior to Populations, Michigan State University, East Lansing, 2022. ProQuest. Available online: https://ezproxy.msu.edu/login?url=https://www.proquest.com/dissertations-theses/effects-neurotoxic-contaminants-on-larval-fish/docview/2728171784/se-2 (accessed on 9 May 2023).
- Sparkes, B.L.; Slone, E.E.A.; Roth, M.; Welti, R.; Fleming, S.D. Intestinal lipid alterations occur prior to antibody-induced prostaglandin E2 production in a mouse model of ischemia/reperfusion. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2010, 1801, 517–525. [Google Scholar] [CrossRef]
- Zhou, Z.; Marepally, S.R.; Nune, D.S.; Pallakollu, P.; Ragan, G.; Roth, M.R.; Wang, L.; Lushington, G.H.; Visvanathan, M.; Welti, R. LipidomeDB Data Calculation Environment: Online Processing of Direct-Infusion Mass Spectral Data for Lipid Profiles. Lipids 2011, 46, 879–884. [Google Scholar] [CrossRef]
- Wheeler, R.E. Permutation Tests for Linear Models in R. 2016. Available online: https://cran.r-project.org/web/packages/lmPerm/vignettes/lmPerm.pdf (accessed on 22 September 2021).
- Savi, M.K.; Mangamana, E.T.; Deguenon, J.M.; Hounmenou, C.G.; Kakaï, R.G. Determination of Lethal Concentrations Using an R Software Function Integrating the Abbott Correction. J. Agric. Sci. Technol. A 2017, 7, 25–30. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, version 4.0.4, x86_64-w64-mingw32/x64 (64-bit); R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.; Dennis, E.A. Update of the LIPID MAPS Comprehensive Classification System for Lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef]
- Rewerts, J.N.; Christie, E.C.; Robel, A.E.; Anderson, T.A.; McCarthy, C.; Salice, C.J.; and Field, J.A. Key Considerations for Accurate Exposures in Ecotoxicological Assessments of Perfluorinated Carboxylates and Sulfonates. Environ. Toxicol. Chem. 2021, 40, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, Z.; Zheng, X.; Ma, Z.; Liu, H.; Giesy, J.P.; Xie, Y.; and Yu, H. Comparison of waterborne and in ovo nanoinjection exposures to assess effects of PFOS on zebrafish embryos. Environ. Sci. Pollut. Res. 2015, 22, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Weiss-Errico, M.; Berry, J.; O’Shea, K. β-Cyclodextrin Attenuates Perfluorooctanoic Acid Toxicity in the Zebrafish Embryo Model. Toxics 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- von Krogh, K.; Higgins, J.; Saavedra Torres, Y.; Mocho, J.P. Screening of Anaesthetics in Adult Zebrafish (Danio rerio) for the Induction of Euthanasia by Overdose. Biology 2021, 10, 1133. [Google Scholar] [CrossRef] [PubMed]
- Leyden, C.; Brueggemann, T.; Debinski, F.; Simacek, C.A.; Dehmelt, F.A.; Arrenberg, A.B. Efficacy of Tricaine (MS-222) and Hypothermia as Anesthetic Agents for Blocking Sensorimotor Responses in Larval Zebrafish. Front. Vet. Sci. 2022, 9, 864573. [Google Scholar] [CrossRef]
- Chen, J.; Tanguay, R.L.; Tal, T.L.; Gai, Z.; Ma, X.; Bai, C.; Tilton, S.C.; Jin, D.; Yang, D.; Huang, C.; et al. Early life perfluorooctanesulphonic acid (PFOS) exposure impairs zebrafish organogenesis. Aquat. Toxicol. 2014, 150, 124–132. [Google Scholar] [CrossRef]
- Starnes, H.M.; Rock, K.D.; Jackson, T.W.; Belcher, S.M. A Critical Review and Meta-Analysis of Impacts of Per- and Polyfluorinated Substances on the Brain and Behavior. Front. Toxicol. 2022, 4, 881584. [Google Scholar] [CrossRef]
- Yu, T.; Zhou, G.; Cai, Z.; Liang, W.; Du, Y.; Wang, W. Behavioral effects of early-life exposure to perfluorooctanoic acid might synthetically link to multiple aspects of dopaminergic neuron development and dopamine functions in zebrafish larvae. Aquat. Toxicol. 2021, 238, 105926. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, A.; Hooser, B.; Abdelmoneim, A.; Horzmann, K.A.; Freemanc, J.L.; Sepúlveda, M.S. Thyroid disrupting effects of halogenated and next generation chemicals on the swim bladder development of zebrafish. Aquat. Toxicol. 2017, 193, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, G.; Lee, Y.-M.; Zoh, K.-D.; Choi, K. Thyroid disrupting effects of perfluoroundecanoic acid and perfluorotridecanoic acid in zebrafish (Danio rerio) and rat pituitary (GH3) cell line. Chemosphere 2021, 262, 128012. [Google Scholar] [CrossRef] [PubMed]
- Stinckens, E.; Vergauwen, L.; Ankley, G.T.; Blust, R.; Darras, V.M.; Villeneuve, D.L.; Witters, H.; Volz, D.C.; Knapen, D. An AOP-based alternative testing strategy to predict the impact of thyroid hormone disruption on swim bladder inflation in zebrafish. Aquat. Toxicol. 2018, 200, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ulhaq, M.; Örn, S.; Carlsson, G.; Morrison, D.A.; Norrgren, L. Locomotor behavior in zebrafish (Danio rerio) larvae exposed to perfluoroalkyl acids. Aquat. Toxicol. 2013, 144–145, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Chenglian, B.; Tao, X.; Xiaotong, W.; Yuanhong, C.; Daqing, J. PFOA-Induced Developmental Toxicity, Behavior Change and DNA Damage in Zebrafish Embryos. Asian J. Ecotoxicol. 2012, 7, 241–250. [Google Scholar]
- Murphy, C.A.; Albers, J.A.; Armstrong, B.M.; Firkus, T.J.; Ivan, L.N. Emerging Toxicological Methods for Fisheries Biologists in the Twenty-first Century. In Methods for Fish Biology, 2nd ed.; Midway, S., Hasler, C., Chakrabarty, P., Eds.; American Fisheries Society: Bethesda, MD, USA, 2022; pp. 551–592. [Google Scholar] [CrossRef]
- Brockmeier, E.K.; Hodges, G.; Hutchinson, T.H.; Butler, E.; Hecker, M.; Tollefsen, K.E.; Garcia-Reyero, N.; Kille, P.; Becker, D.; Chipman, K.; et al. The Role of Omics in the Application of Adverse Outcome Pathways for Chemical Risk Assessment. Toxicol. Sci. 2017, 158, 252–262. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.T. The Application of Omics Data to the Development of AOPs. In A Systems Biology Approach to Advancing Adverse Outcome Pathways for Risk Assessment; Garcia-Reyero, N., Murphy, C.A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 177–198. [Google Scholar] [CrossRef]
- Orešič, M.; McGlinchey, A.; Wheelock, C.E.; Hyötyläinen, T. Metabolic Signatures of the Exposome—Quantifying the Impact of Exposure to Environmental Chemicals on Human Health. Metabolites 2020, 10, 454. [Google Scholar] [CrossRef]
- Panieri, E.; Baralic, K.; Djukic-Cosic, D.; Buha Djordjevic, A.; Saso, L. PFAS Molecules: A Major Concern for the Human Health and the Environment. Toxics 2022, 10, 44. [Google Scholar] [CrossRef]
- Roth, K.; Imran, Z.; Liu, W.; Petriello, M.C. Diet as an Exposure Source and Mediator of Per- and Polyfluoroalkyl Substance (PFAS) Toxicity. Front. Toxicol. 2020, 2, 601149. [Google Scholar] [CrossRef]
- Yang, Z.; Fu, L.; Cao, M.; Li, F.; Li, J.; Chen, Z.; Guo, A.; Zhong, H.; Li, W.; Liang, Y.; et al. PFAS-Induced Lipidomic Dysregulations and their Assocations with Developmental Toxicity in Zebrafish Embryos. Sci. Total Environ. 2023, 861, 160691. [Google Scholar] [CrossRef]
- Sant, K.E.; Annunziato, K.; Conlin, S.; Teicher, G.; Chen, P.; Venezia, O.; Downes, G.B.; Park, Y.; Timme-Laragy, A.R. Developmental exposures to perfluorooctanesulfonic acid (PFOS) impact embryonic nutrition, pancreatic morphology, and adiposity in the zebrafish, Danio rerio. Environ. Pollut. 2021, 275, 116644. [Google Scholar] [CrossRef]
- Cui, Y.; Lv, S.; Liu, J.; Nie, S.; Chen, J.; Dong, Q.; Huang, C.; Yang, D. Chronic perfluorooctanesulfonic acid exposure disrupts lipid metabolism in zebrafish. Hum. Exp. Toxicol. 2017, 36, 207–217. [Google Scholar] [CrossRef]
- Glade, M.J.; Smith, K. Phosphatidylserine and the human brain. Nutrition 2015, 31, 781–786. [Google Scholar] [CrossRef]
- Leray, C. Introduction to Lipidomics: From Bacteria to Man; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2013. [Google Scholar]
- Hong, J.-R.; Lin, G.-H.; Lin, C.J.-F.; Wang, W.-P.; Lee, C.-C.; Lin, T.-L.; Wu, J.-L. Phosphatidylserine receptor is required for the engulfment of dead apoptotic cells and for normal embryonic development in zebrafish. Development 2004, 131, 5417–5427. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Kawana, H.; Nishino, Y.; Ito, Y.; Sato, H.; Onishi, H.; Kano, K.; Inoue, A.; Taketomi, Y.; Murakami, M.; et al. Abnormal male reproduction and embryonic development induced by downregulation of a phospholipid fatty acid-introducing enzyme Lpgat1 in zebrafish. Sci. Rep. 2022, 12, 7312. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.B.; Jenkins, D.; Chanudet, E.; Tasseva, G.; Ishida, M.; Anderson, G.; Docker, J.; Ryten, M.; Sa, J.; Saraiva, J.M.; et al. Gain-of-function mutations in the phosphatidylserine synthase 1 (PTDSS1) gene cause Lenz-Majewski syndrome. Nat. Genet. 2014, 46, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-J.; Kim, S.; Shim, H.-P.; Park, J.H.; Lee, G.; Kim, T.-Y.; Jo, M.-C.; Kwon, A.-Y.; Lee, M.; Lee, S.; et al. Phosphatidylserine synthase plays an essential role in glia and affects development, as well as the maintenance of neuronal function. Iscience 2021, 24, 102899. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, J.; Hino, K. Plasmalogen in the brain: Effects on cognitive functions and behaviors attributable to its properties. Brain Res. Bull. 2022, 188, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Dorninger, F.; König, T.; Scholze, P.; Berger, M.L.; Zeitler, G.; Wiesinger, C.; Gundacker, A.; Pollak, D.D.; Huck, S.; Just, W.W.; et al. Disturbed neurotransmitter homeostasis in ether lipid deficiency. Hum. Mol. Genet. 2019, 28, 2046–2061. [Google Scholar] [CrossRef] [PubMed]
- Fraher, D.; Sanigorski, A.; Mellett, N.A.; Meikle, P.J.; Sinclair, A.J.; Gibert, Y. Zebrafish embryonic lipidomic analysis reveals that the yolk cell is metabolically active in processing lipid. Cell Rep. 2016, 14, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, L.; Zhang, R.; Shi, Z.; Xie, C.; Hong, Y.; Wang, J.; Cai, Z. Spatially revealed perfluorooctane sulfonate-induced nephrotoxicity in mouse kidney using atmospheric pressure MALDI mass spectrometry imaging. Sci. Total Environ. 2022, 838, 156380. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.T.; Wang, L.; Jia, R.W.; Fu, X.H.; Chua, H. Lipophilic organic pollutants induce changes in phospholipid and membrane protein composition leading to Vero cell morphological change. J. Environ. Sci. Health Part B 2014, 49, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Viada, B.; Cámara, C.I.; Yudi, L.M. Destabilizing effect of perfluorodecanoic acid on simple membrane models. Soft Matter 2019, 15, 2447–2462. [Google Scholar] [CrossRef]
- Wójcik, A.; Perczyk, P.; Wydro, P.; Broniatowski, M. Effects of water soluble perfluorinated pollutants on phospholipids in model soil decomposer membranes. Biochim. Biophys. Acta (BBA)—Biomembr. 2018, 1860, 2576–2587. [Google Scholar] [CrossRef]
- Arukwe, A.; Cangialosi, M.V.; Letcher, R.J.; Rocha, E.; Mortensen, A.S. Changes in morphometry and association between whole-body fatty acids and steroid hormone profiles in relation to bioaccumulation patterns in salmon larvae exposed to perfluorooctane sulfonic or perfluorooctane carboxylic acids. Aquat. Toxicol. 2013, 130–131, 219–230. [Google Scholar] [CrossRef]
- Epand, R.M. Cholesterol in Bilayers of Sphingomyelin or Dihydrosphingomyelin at Concentrations Found in Ocular Lens Membranes. Biophys. J. 2003, 84, 3102–3110. [Google Scholar] [CrossRef]
- Ryu, S.B. Phospholipid-derived signaling mediated by phospholipase A in plants. Trends Plant Sci. 2004, 9, 229–235. [Google Scholar] [CrossRef]
- Okochi, E.; Nishimaki-Mogami, T.; Suzuki, K.; Takahashi, A. Perfluorooctanoic acid, a peroxisome-proliferating hypolipidemic agent, dissociates apolipoprotein B48 from lipoprotein particles and decreases secretion of very low density lipoproteins by cultured rat hepatocytes. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 1999, 1437, 393–401. [Google Scholar] [CrossRef]
- Blanc, M.; Alfonso, S.; Bégout, M.L.; Barrachina, C.; Hyötyläinen, T.; Keiter, S.H.; Cousin, X. An environmentally relevant mixture of polychlorinated biphenyls (PCBs) and polybrominated diphenylethers (PBDEs) disrupts mitochondrial function, lipid metabolism and neurotransmission in the brain of exposed zebrafish and their unexposed F2 offspring. Sci. Total Environ. 2021, 754, 142097. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, P. The impaired swim bladder via ROS-mediated inhibition of the Wnt/Hedgehog pathway in zebrafish embryos exposed to eight toxic chemicals and binary chemical mixtures. Chemosphere 2023, 338, 139593. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wu, P.; Wang, L.; Li, Q.; Li, X.; Luo, Y. Toxicity of per-and polyfluoroalkyl substances to aquatic vertebrates. Front. Environ. Sci. 2023, 11, 37. [Google Scholar] [CrossRef]
- Chen, J.; Zheng, L.; Tian, L.; Wang, N.; Lei, L.; Wang, Y.; Dong, Q.; Huang, C.; Yang, D. Chronic PFOS exposure disrupts thyroid structure and function in zebrafish. Bull. Environ. Contam. Toxicol. 2018, 101, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Stinckens, E.; Vergauwen, L.; Blackwell, B.R.; Ankley, G.T.; Villeneuve, D.L.; Knapen, D. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environ. Sci. Technol. 2020, 54, 6213–6223. [Google Scholar] [CrossRef] [PubMed]
- Horie, Y.; Nomura, M.; Okamoto, K.; Takahashi, C.; Sato, T.; Miyagawa, S.; Okamura, H.; Iguchi, T. Effect of thyroid hormone-disrupting chemicals on swim bladder inflation and thyroid hormone-related gene expression in Japanese medaka and zebrafish. J. Appl. Toxicol. 2022, 42, 1385–1395. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
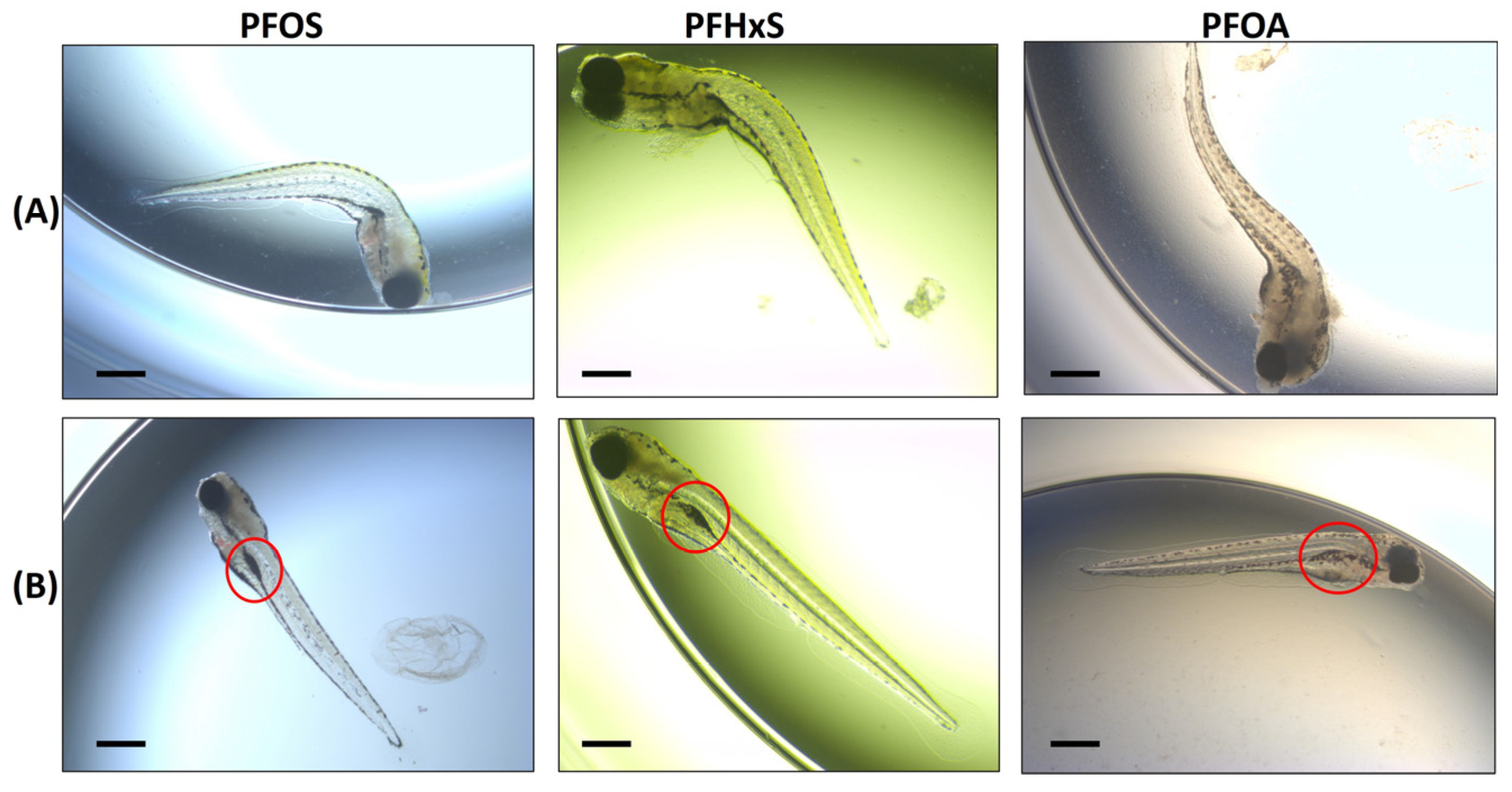
| Morphological Endpoint | Definition |
|---|---|
| Mortality | Lack of heartbeat or obvious necrosis or ruptured pericardium. |
| Spine Curvature | Curvature of the spine/upper tail similar to the scoliosis phenotype described in von Hellfeld et al. (2020) [21]. Documented on live and dead embryos. |
| Swim Bladder | Incidences where the swim bladder was either underinflated or uninflated. Documented on live embryos. |
| Impaired Equilibrium | Embryos that were either on their side or had difficulties staying in the upright or “dorsal-up” position, which can also be described as “listing”. Documented on live embryos with no external stimulus applied. |
| Chemical | Nominal Concentration ppm | Media Concentration ppm (% Recovery) | Embryo Tissue Concentration ng/mg (Std) | Average Number of Embryos in 3 Pools | ||
|---|---|---|---|---|---|---|
| PFOA | 0 | <LOD of 20 ng/L | 12.24 (6.1) | 16.0 | ||
| 200 | 188 | (94.0) | 487 | (85.9) | 16.0 | |
| 250 | 242 | (96.8) | 263 | (25.0) | 15.7 | |
| 300 | 253 | (84.3) | 305 | (10.4) | 16.0 | |
| 350 | 294 | (84.0) | 361 | (49.3) | 15.3 | |
| 375 | 375 | (100.0) | 485 | (50.6) | 13.7 | |
| PFHxS | 0 | <LOD of 20 ng/L | 0.2 (0.3) | 16.0 | ||
| 7.5 | 6.35 | (84.7) | 69.0 | (20.9) | 15.7 | |
| 10 | 8.70 | (87.0) | 91.5 | (12.3) | 13.7 | |
| 12.5 | 10.15 | (81.2) | 71.4 | (7.0) | 14.3 | |
| 15 | 11.50 | (76.7) | 76.0 | (25.5) | 11.3 | |
| 17.5 | 14.35 | (82.0) | 94.5 | (13.6) | 7.3 | |
| PFOS | 0 | <LOD of 20 ng/L | 2.32 (2.8) | 8.0 | ||
| 1 | 1.21 | (120.9) | 148 | (36.7) | 8.0 a | |
| 1.25 | 1.40 | (111.9) | 227 | (77.7) | 8.0 a | |
| 1.5 | 1.86 | (124.2) | 197 | (53.1) | 8.0 a | |
| 1.75 | 2.03 | (116.1) | 183 | (153.6) | 8.0 a | |
| 2 | 2.42 | (121.0) | 103 | (78.5) | 8.0 a | |
| Control versus Dose Trend | |||||
|---|---|---|---|---|---|
| Lipid Group | 188 ppm | 242 ppm | 253 ppm | 294 ppm | 375 ppm |
| PE(34:1) | - | - | - | Decrease | - |
| PE(36:5) | Decrease | - | - | - | - |
| PS(34:1) | - | - | - | Decrease | Decrease * |
| PS(38:4) | - | - | Decrease | - | Decrease |
| PS(40:5) | - | - | - | Decrease | - |
| PS(42:6) | Decrease | - | Decrease | - | Decrease |
| PS(42:9) | Decrease | - | - | Decrease * | Decrease |
| PS(44:12) | Decrease | - | - | - | - |
| Total PS | Decrease | - | - | - | - |
| ePE(36:1) | - | - | - | Decrease | - |
| Total ePC | - | - | - | Decrease * | Decrease * |
| Total LysoPE | - | - | - | Decrease | - |
| DSM(16:0) | - | - | - | - | Decrease * |
| Total SM and DSM | - | - | - | Decrease | - |
| Control verses Dose Trend | |||||
|---|---|---|---|---|---|
| Lipid Group | 6.4 ppm | 8.7 ppm | 10.2 ppm | 11.5 ppm | 14.4 ppm |
| ePC(34:1) | - | - | - | - 1 | Decrease 1 |
| ePC(36:2) | - | - | - | - 1 | Decrease *,1 |
| Total ePC | - | - | - | - 1 | Decrease 1 |
| PC(32:1) | - | - | - | - 1 | Decrease 1 |
| PC(34:3) | - | - | - | - 1 | Decrease 1 |
| PE(36:5) | - | - | - | - 1 | Decrease 1 |
| PE(46:12) | - | - | - | - 1 | Decrease 1 |
| PS(38:4) | - | - | Decrease | - 1 | Decrease 1 |
| PS(40:5) | - | - | - | - 1 | Decrease 1 |
| Control verses Dose Trend | |||||
|---|---|---|---|---|---|
| Lipid Group | 1.21 ppm | 1.40 ppm | 1.86 ppm | 2.03 ppm | 2.42 ppm |
| FA(16:0) | - | - | - | - | Increase 1 |
| FA(18:1) | - | - | - | - | Increase 1 |
| FA(18:2) | - | - | - | - | Increase 1 |
| FA(20:1) | - | - | - | - | Increase 1 |
| FA(20:2) | - | - | - | - | Increase 1 |
| FA(20:5) | - | - | - | - | Increase 1 |
| FA(22:6) | - | - | - | - | Increase 1 |
| PA(34:1) | - | Decrease | Decrease * | Decrease * | Decrease 1 |
| PE(32:1) | - | - | - | - | Increase *,1 |
| PE(38:3) | - | - | - | - | Decrease 1 |
| PE(38:4) | - | - | - | - | Decrease 1 |
| PE(38:5) | - | - | - | Decrease | Decrease 1 |
| PE(40:8) | - | - | - | - | Decrease 1 |
| PE(42:10) | - | - | - | - | Decrease 1 |
| PE(42:8) | - | - | - | Decrease | Decrease 1 |
| PS(40:5) | - | - | - | Decrease | Decrease 1 |
| PS(40:6) | - | - | - | Decrease | Decrease 1 |
| Total_PS | - | - | - | - | Decrease 1 |
| ePE(40:2) | - | - | - | - | Decrease 1 |
| Total_ePE | - | - | - | - | Decrease 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albers, J.; Mylroie, J.; Kimble, A.; Steward, C.; Chapman, K.; Wilbanks, M.; Perkins, E.; Garcia-Reyero, N. Per- and Polyfluoroalkyl Substances: Impacts on Morphology, Behavior and Lipid Levels in Zebrafish Embryos. Toxics 2024, 12, 192. https://doi.org/10.3390/toxics12030192
Albers J, Mylroie J, Kimble A, Steward C, Chapman K, Wilbanks M, Perkins E, Garcia-Reyero N. Per- and Polyfluoroalkyl Substances: Impacts on Morphology, Behavior and Lipid Levels in Zebrafish Embryos. Toxics. 2024; 12(3):192. https://doi.org/10.3390/toxics12030192
Chicago/Turabian StyleAlbers, Janice, John Mylroie, Ashley Kimble, Catherine Steward, Kacy Chapman, Mitchell Wilbanks, Edward Perkins, and Natàlia Garcia-Reyero. 2024. "Per- and Polyfluoroalkyl Substances: Impacts on Morphology, Behavior and Lipid Levels in Zebrafish Embryos" Toxics 12, no. 3: 192. https://doi.org/10.3390/toxics12030192