
FABP4 Is an Indispensable Factor for Regulating Cellular Metabolic Functions of the Human Retinal Choroid

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Two-Dimensional (2D) Culture of HOCF Cells, HNPCE Cells, RB Cells and ARPE19 Cells
2.2. Measurement of Seahorse Cellular Metabolic Functions
2.3. RNA Sequencing, Gene Function and Analysis of Pathways
2.4. Other Analytical Methods
3. Results
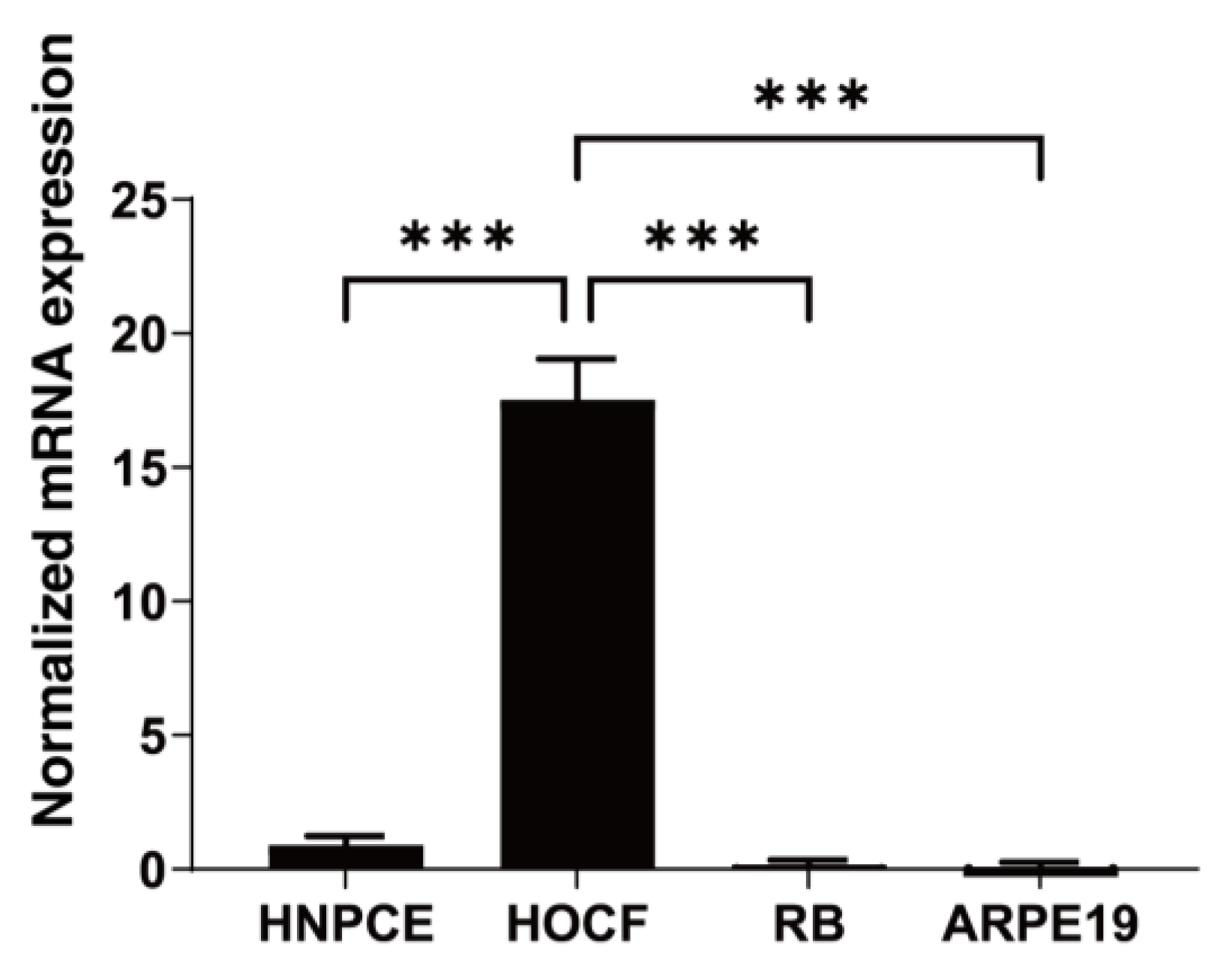
3.1. HOCF Cells Are the Primary Origin of Intraocularly Produced FABP4 among Cells Constituting the Intraocular Tissues
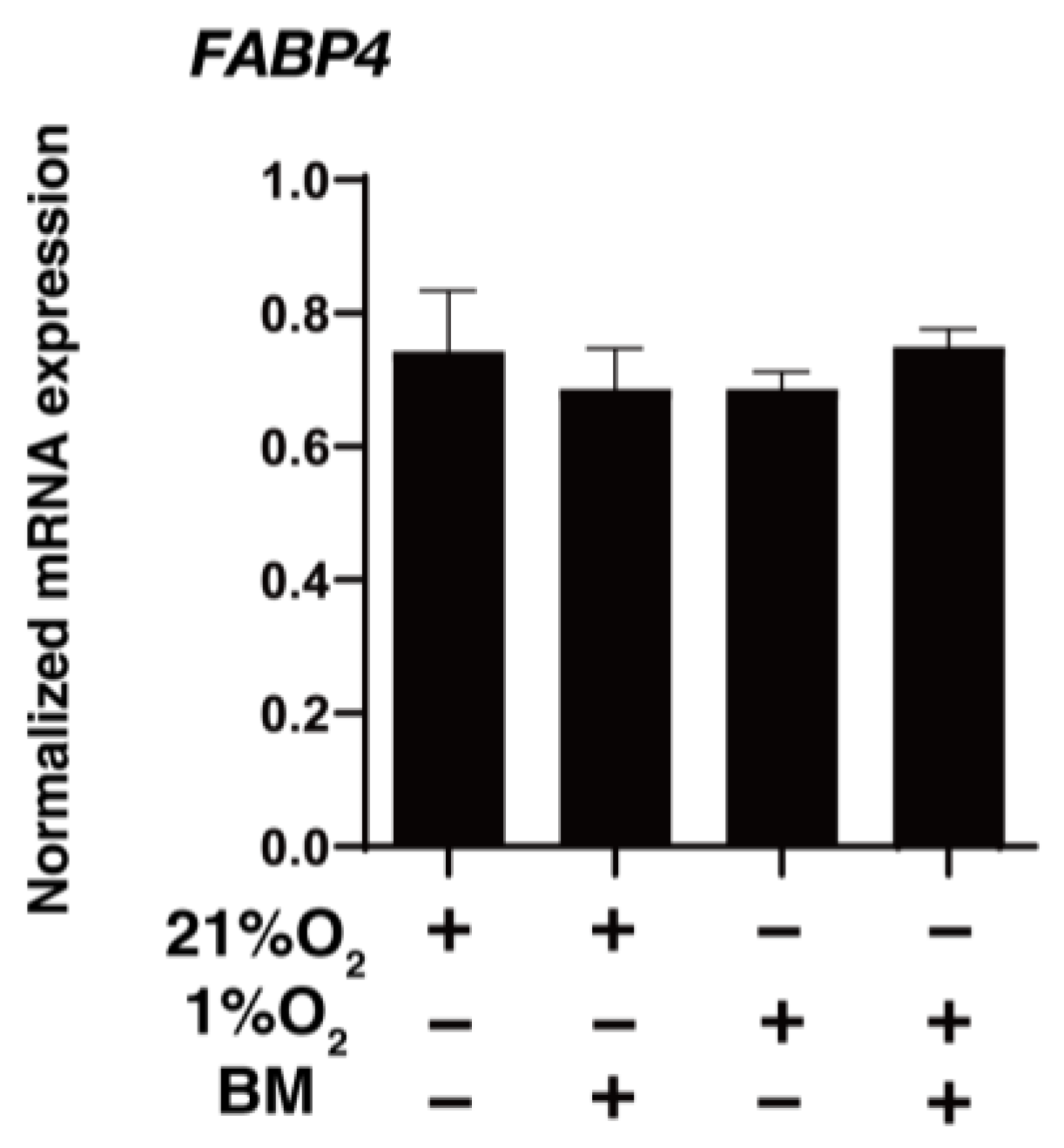
3.2. Effects of FABP4 on Cellular Metabolic Functions in HOCF Cells
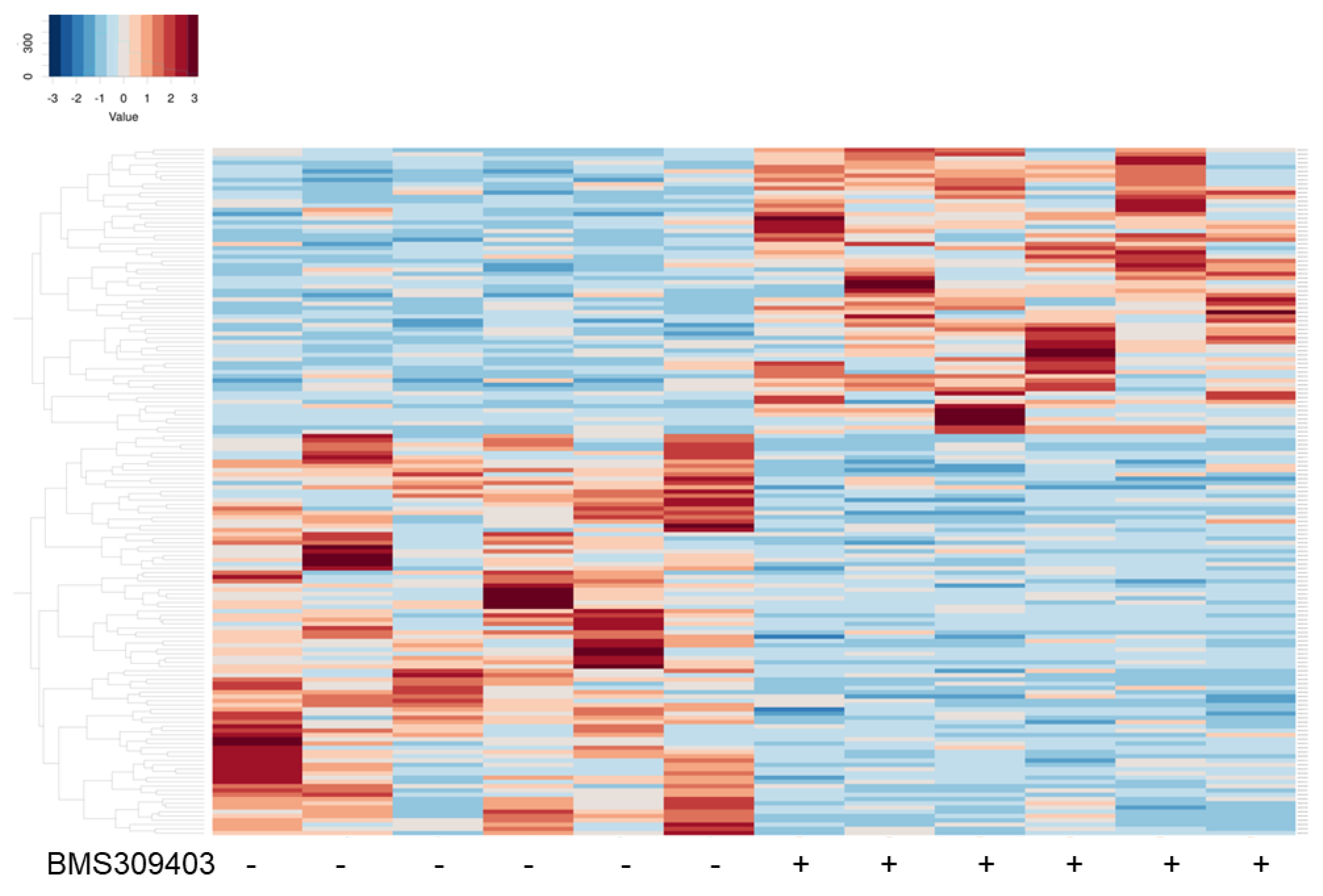
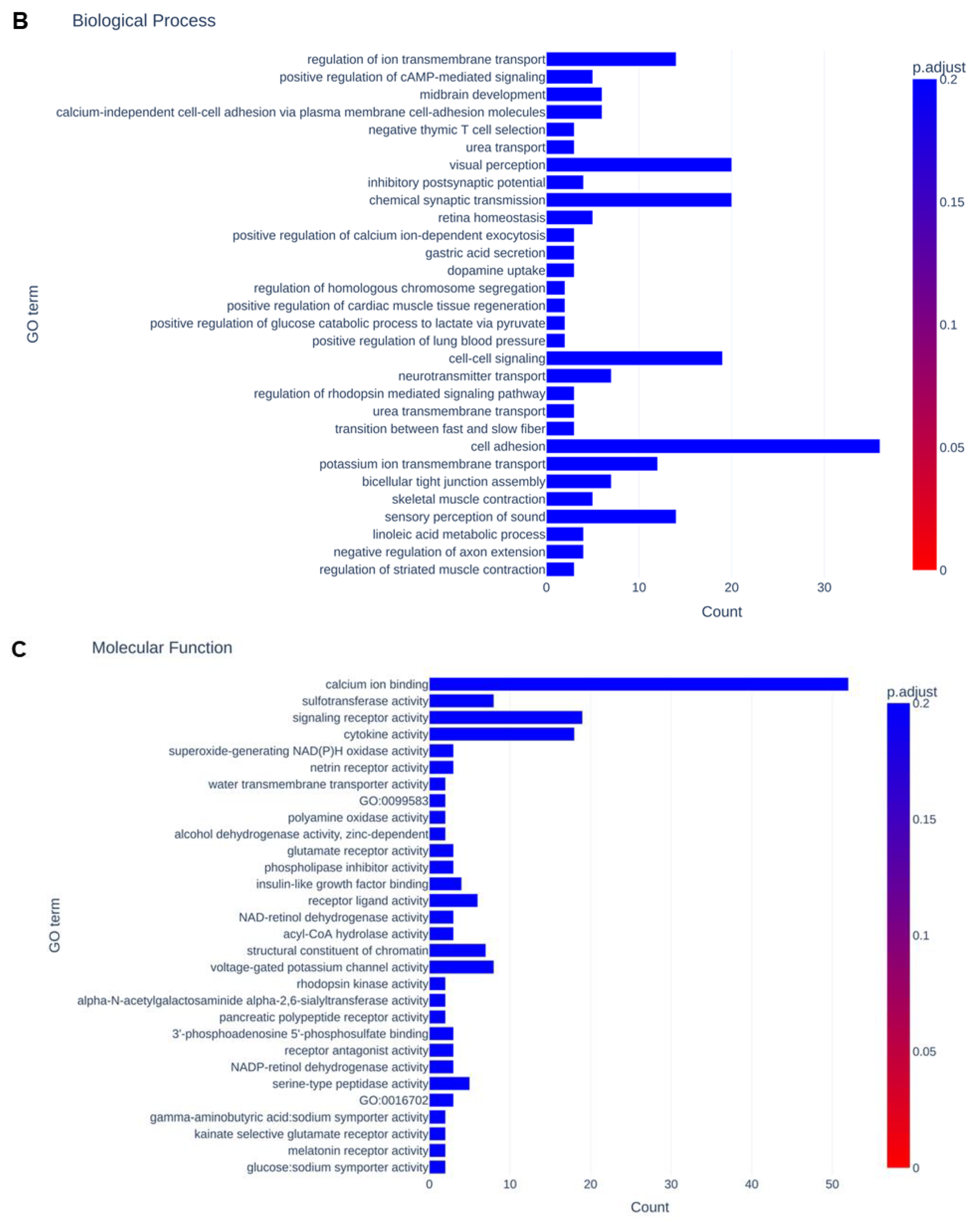
3.3. Functional Analysis of Differentially Expressed Genes That Were Modulated by the Pharmacological Inhibition of FABP4
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furuhashi, M.; Hotamisligil, G.S. Fatty acid-binding proteins: Role in metabolic diseases and potential as drug targets. Nat. Rev. Drug Discov. 2008, 7, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Bernlohr, D.A. Metabolic functions of FABPs—Mechanisms and therapeutic implications. Nat. Rev. Endocrinol. 2015, 11, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Wang, Y.; Xu, J.Y.; Stejskal, D.; Tam, S.; Zhang, J.; Wat, N.M.; Wong, W.K.; Lam, K.S. Adipocyte fatty acid-binding protein is a plasma biomarker closely associated with obesity and metabolic syndrome. Clin. Chem. 2006, 52, 405–413. [Google Scholar] [CrossRef]
- Cabré, A.; Lázaro, I.; Girona, J.; Manzanares, J.M.; Marimón, F.; Plana, N.; Heras, M.; Masana, L. Plasma fatty acid binding protein 4 is associated with atherogenic dyslipidemia in diabetes. J. Lipid Res. 2008, 49, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Yeung, D.C.; Xu, A.; Cheung, C.W.; Wat, N.M.; Yau, M.H.; Fong, C.H.; Chau, M.T.; Lam, K.S. Serum adipocyte fatty acid-binding protein levels were independently associated with carotid atherosclerosis. Arter. Thromb. Vasc. Biol. 2007, 27, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Ma, L.; Fu, P. Role of Fatty Acid Binding Protein 4 (FABP4) in Kidney Disease. Curr. Med. Chem. 2020, 27, 3657–3664. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Calvo, R.; Girona, J.; Alegret, J.M.; Bosquet, A.; Ibarretxe, D.; Masana, L. Role of the fatty acid-binding protein 4 in heart failure and cardiovascular disease. J. Endocrinol. 2017, 233, R173–R184. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Zhao, X. Therapeutic Implications of FABP4 in Cancer: An Emerging Target to Tackle Cancer. Front. Pharmacol. 2022, 13, 948610. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Furuhashi, M.; Ida, Y.; Ohguro, H.; Watanabe, M.; Suzuki, S.; Hikage, F. Detection of significantly high vitreous concentrations of fatty acid-binding protein 4 in patients with proliferative diabetic retinopathy. Sci. Rep. 2021, 11, 12382. [Google Scholar] [CrossRef] [PubMed]
- Hikage, F.; Furuhashi, M.; Ida, Y.; Ohguro, H.; Watanabe, M.; Suzuki, S.; Itoh, K. Fatty acid-binding protein 4 is an independent factor in the pathogenesis of retinal vein occlusion. PLoS ONE 2021, 16, e0245763. [Google Scholar] [CrossRef]
- Flaxman, S.R.; Bourne, R.R.A.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; et al. Global causes of blindness and distance vision impairment 1990-2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.; Mitchell, P.; Wong, T.Y. Diabetic retinopathy. Lancet 2010, 376, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Xu, Y.; Zha, M.; Zhang, Y.; Rudan, I. Global epidemiology of retinal vein occlusion: A systematic review and meta-analysis of prevalence, incidence, and risk factors. J. Glob. Health 2019, 9, 010427. [Google Scholar] [CrossRef]
- Rogers, S.; McIntosh, R.L.; Cheung, N.; Lim, L.; Wang, J.J.; Mitchell, P.; Kowalski, J.W.; Nguyen, H.; Wong, T.Y. The prevalence of retinal vein occlusion: Pooled data from population studies from the United States, Europe, Asia, and Australia. Ophthalmology 2010, 117, 313–319.e311. [Google Scholar] [CrossRef] [PubMed]
- Umetsu, A.; Furuhashi, M.; Watanabe, M.; Ohkawa, E.; Tsugeno, Y.; Suzuki, S.; Itoh, K.; Ida, Y.; Hikage, F.; Ohguro, H. Fatty acid metabolism is involved in both retinal physiology and the pathology of retinal vascular diseases. Prostaglandins Leukot. Essent. Fat Acids 2022, 183, 102473. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Sato, T.; Tsugeno, Y.; Higashide, M.; Furuhashi, M.; Ohguro, H. TGF-β-3 Induces Different Effects from TGF-β-1 and -2 on Cellular Metabolism and the Spatial Properties of the Human Trabecular Meshwork Cells. Int. J. Mol. Sci. 2023, 24, 4181. [Google Scholar] [CrossRef] [PubMed]
- Ichioka, H.; Hirohashi, Y.; Sato, T.; Furuhashi, M.; Watanabe, M.; Ida, Y.; Hikage, F.; Torigoe, T.; Ohguro, H. G-Protein-Coupled Receptors Mediate Modulations of Cell Viability and Drug Sensitivity by Aberrantly Expressed Recoverin 3 within A549 Cells. Int. J. Mol. Sci. 2023, 24, 771. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Shao, Z.; Wang, K.; Zhang, S.; Yuan, J.; Liao, X.; Wu, C.; Zou, Y.; Ha, Y.; Shen, Z.; Guo, J.; et al. Ingenuity pathway analysis of differentially expressed genes involved in signaling pathways and molecular networks in RhoE gene-edited cardiomyocytes. Int. J. Mol. Med. 2020, 46, 1225–1238. [Google Scholar] [CrossRef]
- Alimadadi, A.; Aryal, S.; Manandhar, I.; Joe, B.; Cheng, X. Identification of Upstream Transcriptional Regulators of Ischemic Cardiomyopathy Using Cardiac RNA-Seq Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 3472. [Google Scholar] [CrossRef]
- Ida, Y.; Hikage, F.; Itoh, K.; Ida, H.; Ohguro, H. Prostaglandin F2α agonist-induced suppression of 3T3-L1 cell adipogenesis affects spatial formation of extra-cellular matrix. Sci. Rep. 2020, 10, 7958. [Google Scholar] [CrossRef]
- Furuhashi, M.; Tuncman, G.; Görgün, C.Z.; Makowski, L.; Atsumi, G.; Vaillancourt, E.; Kono, K.; Babaev, V.R.; Fazio, S.; Linton, M.F.; et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature 2007, 447, 959–965. [Google Scholar] [CrossRef]
- Nickla, D.L.; Wallman, J. The multifunctional choroid. Prog. Retin. Eye Res. 2010, 29, 144–168. [Google Scholar] [CrossRef]
- Lakkaraju, A.; Umapathy, A.; Tan, L.X.; Daniele, L.; Philp, N.J.; Boesze-Battaglia, K.; Williams, D.S. The cell biology of the retinal pigment epithelium. Prog. Retin. Eye Res. 2020, 78, 100846. [Google Scholar] [CrossRef]
- Skeie, J.M.; Mahajan, V.B. Proteomic landscape of the human choroid-retinal pigment epithelial complex. JAMA Ophthalmol. 2014, 132, 1271–1281. [Google Scholar] [CrossRef]
- Soundara Pandi, S.P.; Chen, M.; Guduric-Fuchs, J.; Xu, H.; Simpson, D.A. Extremely complex populations of small RNAs in the mouse retina and RPE/choroid. Investig. Ophthalmol. Vis. Sci. 2013, 54, 8140–8151. [Google Scholar] [CrossRef]
- Voigt, A.P.; Mulfaul, K.; Mullin, N.K.; Flamme-Wiese, M.J.; Giacalone, J.C.; Stone, E.M.; Tucker, B.A.; Scheetz, T.E.; Mullins, R.F. Single-cell transcriptomics of the human retinal pigment epithelium and choroid in health and macular degeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 24100–24107. [Google Scholar] [CrossRef]
- Wang, S.; Liu, S.; Mao, J.; Wen, D. Effect of retinoic acid on the tight junctions of the retinal pigment epithelium-choroid complex of guinea pigs with lens-induced myopia in vivo. Int. J. Mol. Med. 2014, 33, 825–832. [Google Scholar] [CrossRef]
- Ibuki, M.; Lee, D.; Shinojima, A.; Miwa, Y.; Tsubota, K.; Kurihara, T. Rice Bran and Vitamin B6 Suppress Pathological Neovascularization in a Murine Model of Age-Related Macular Degeneration as Novel HIF Inhibitors. Int. J. Mol. Sci. 2020, 21, 8940. [Google Scholar] [CrossRef] [PubMed]
- Pietras, A.; Johnsson, A.S.; Påhlman, S. The HIF-2α-driven pseudo-hypoxic phenotype in tumor aggressiveness, differentiation, and vascularization. Curr. Top. Microbiol. Immunol. 2010, 345, 1–20. [Google Scholar] [CrossRef]
- Påhlman, S.; Mohlin, S. Hypoxia and hypoxia-inducible factors in neuroblastoma. Cell Tissue Res. 2018, 372, 269–275. [Google Scholar] [CrossRef]
- Azimi, I. The interplay between HIF-1 and calcium signalling in cancer. Int. J. Biochem. Cell Biol. 2018, 97, 73–77. [Google Scholar] [CrossRef]
- Suzuki, K.; Lavaroni, S.; Mori, A.; Okajima, F.; Kimura, S.; Katoh, R.; Kawaoi, A.; Kohn, L.D. Thyroid transcription factor 1 is calcium modulated and coordinately regulates genes involved in calcium homeostasis in C cells. Mol. Cell. Biol. 1998, 18, 7410–7422. [Google Scholar] [CrossRef]
- Xi, S.; Wang, H.; Chen, J.; Gan, T.; Zhao, L. LncRNA GAS5 Attenuates Cardiac Electrical Remodeling Induced by Rapid Pacing via the miR-27a-3p/HOXa10 Pathway. Int. J. Mol. Sci. 2023, 24, 12093. [Google Scholar] [CrossRef]
- Wanet, A.; Bassal, M.A.; Patel, S.B.; Marchi, F.; Mariani, S.A.; Ahmed, N.; Zhang, H.; Borchiellini, M.; Chen, S.; Zhang, J.; et al. E-cadherin is regulated by GATA-2 and marks the early commitment of mouse hematopoietic progenitors to the basophil and mast cell fates. Sci. Immunol. 2021, 6, 56. [Google Scholar] [CrossRef]
- Nurminen, V.; Seuter, S.; Carlberg, C. Primary Vitamin D Target Genes of Human Monocytes. Front. Physiol. 2019, 10, 194. [Google Scholar] [CrossRef]
- Miller, B.A. TRPM2 in Cancer. Cell Calcium 2019, 80, 8–17. [Google Scholar] [CrossRef]
- Ipposhi, K.; Tomokiyo, A.; Ono, T.; Yamashita, K.; Alhasan, M.A.; Hasegawa, D.; Hamano, S.; Yoshida, S.; Sugii, H.; Itoyama, T.; et al. Secreted Frizzled-Related Protein 1 Promotes Odontoblastic Differentiation and Reparative Dentin Formation in Dental Pulp Cells. Cells 2021, 10, 2491. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Farage, E.; Sugimoto, M.; Anand-Apte, B. A novel transgenic zebrafish model for blood-brain and blood-retinal barrier development. BMC Dev. Biol. 2010, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Z.; Saxena, V.; Sharma, M.K.; Thisse, C.; Thisse, B.; Denovan-Wright, E.M.; Wright, J.M. The fabp4 gene of zebrafish (Danio rerio)–genomic homology with the mammalian FABP4 and divergence from the zebrafish fabp3 in developmental expression. Febs J. 2007, 274, 1621–1633. [Google Scholar] [CrossRef]
- Bisgrove, D.A.; Godbout, R. Differential expression of AP-2alpha and AP-2beta in the developing chick retina: Repression of R-FABP promoter activity by AP-2. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1999, 214, 195–206. [Google Scholar] [CrossRef]
- Godbout, R.; Marusyk, H.; Bisgrove, D.; Dabbagh, L.; Poppema, S. Localization of a fatty acid binding protein and its transcript in the developing chick retina. Exp. Eye Res. 1995, 60, 645–657. [Google Scholar] [CrossRef]
- Huang, H.W.; Ryoo, H.D. Drosophila fabp is required for light-dependent Rhodopsin-1 clearance and photoreceptor survival. PLoS Genet. 2021, 17, e1009551. [Google Scholar] [CrossRef]
- Fu, Z.; Kern, T.S.; Hellström, A.; Smith, L.E.H. Fatty acid oxidation and photoreceptor metabolic needs. J. Lipid Res. 2021, 62, 100035. [Google Scholar] [CrossRef]
- Busik, J.V. Lipid metabolism dysregulation in diabetic retinopathy. J. Lipid Res. 2021, 62, 100017. [Google Scholar] [CrossRef]
- Escandon, P.; Vasini, B.; Whelchel, A.E.; Nicholas, S.E.; Matlock, H.G.; Ma, J.X.; Karamichos, D. The role of peroxisome proliferator-activated receptors in healthy and diseased eyes. Exp. Eye Res. 2021, 208, 108617. [Google Scholar] [CrossRef]
- Floresta, G.; Pistarà, V.; Amata, E.; Dichiara, M.; Marrazzo, A.; Prezzavento, O.; Rescifina, A. Adipocyte fatty acid binding protein 4 (FABP4) inhibitors. A comprehensive systematic review. Eur. J. Med. Chem. 2017, 138, 854–873. [Google Scholar] [CrossRef]
- Floresta, G.; Patamia, V.; Zagni, C.; Rescifina, A. Adipocyte fatty acid binding protein 4 (FABP4) inhibitors. An update from 2017 to early 2022. Eur. J. Med. Chem. 2022, 240, 114604. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulation | Down-Regulation | ||
|---|---|---|---|
| Molecules | p-Value | Molecules | p-Value |
| KIF1A | 6.301 | CHRNA1 | −5.838 |
| CDHR1 | 6.156 | TNFSF18 | −5.603 |
| CSTA | 5.410 | IDO1 | −5.018 |
| LCN1 | 5.271 | LOC105377155 | −4.997 |
| HSPA6 | 4.990 | LOC102724434 | −4.661 |
| LINC01705 | 4.837 | ENSG00000289492 | −4.658 |
| PIP5K1B | 4.719 | CFI | −4.610 |
| LINC02568 | 4.679 | AKAP13-AS1 | −4.605 |
| ZBBX | 4.607 | MYO16 | −4.555 |
| AGA-DT | 4.593 | RP11_885L141 | −4.502 |
| Name | p-Value of Range |
|---|---|
| Cellular Movement | 1.58 × 10−6–1.98 × 10−21 |
| Cellular Development | 1.47 × 10−6–4.12 × 10−19 |
| Cellular Function and Maintenance | 5.00 × 10−7–4.12 × 10−19 |
| Cellular Growth and Proliferation | 1.24 × 10−6–4.12 × 10−19 |
| Cell-to-Cell Signaling and Interaction | 1.58 × 10−6–6.61 × 10−16 |
| Name | p-Value |
|---|---|
| rhodopsin-like receptors | 2.34 × 10−8 |
| G-alpha (i) signaling events | 3.08 × 10−6 |
| cAMP-response element-binding protein (CREB) signaling in neurons | 6.25 × 10−6 |
| S100 family signaling pathway | 7.71 × 10−6 |
| visual phototransduction and adrenergic receptor signaling | 1.51 × 10−5 |
| Name | Score |
|---|---|
| developmental disorder | 43 |
| cell morphology | 41 |
| cell-to-cell signaling and interaction | 41 |
| cell death and survival | 37 |
| neurological disease | 34 |
| Regulators | Disease and Function | Consistency Score |
|---|---|---|
| ADIPOQ, IL25, RNF31 | acute disease, immune response of leukocytes | 7.542 |
| CASP4, CEBPA, CLEC11A, RARB | acute disease, hypertensive reaction | 2.296 |
| ADIPOQ | polarization of myeloid cells | −6 |
| ADIPOQ | polarization of phagocytes | −6 |
| RARB | secretion of neurotransmitter | −8.083 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohguro, H.; Watanabe, M.; Sato, T.; Nishikiori, N.; Umetsu, A.; Higashide, M.; Ogawa, T.; Furuhashi, M. FABP4 Is an Indispensable Factor for Regulating Cellular Metabolic Functions of the Human Retinal Choroid. Bioengineering 2024, 11, 584. https://doi.org/10.3390/bioengineering11060584
Ohguro H, Watanabe M, Sato T, Nishikiori N, Umetsu A, Higashide M, Ogawa T, Furuhashi M. FABP4 Is an Indispensable Factor for Regulating Cellular Metabolic Functions of the Human Retinal Choroid. Bioengineering. 2024; 11(6):584. https://doi.org/10.3390/bioengineering11060584
Chicago/Turabian StyleOhguro, Hiroshi, Megumi Watanabe, Tatsuya Sato, Nami Nishikiori, Araya Umetsu, Megumi Higashide, Toshifumi Ogawa, and Masato Furuhashi. 2024. "FABP4 Is an Indispensable Factor for Regulating Cellular Metabolic Functions of the Human Retinal Choroid" Bioengineering 11, no. 6: 584. https://doi.org/10.3390/bioengineering11060584
APA StyleOhguro, H., Watanabe, M., Sato, T., Nishikiori, N., Umetsu, A., Higashide, M., Ogawa, T., & Furuhashi, M. (2024). FABP4 Is an Indispensable Factor for Regulating Cellular Metabolic Functions of the Human Retinal Choroid. Bioengineering, 11(6), 584. https://doi.org/10.3390/bioengineering11060584