Abstract
Cordyceps militaris is considered to be of great medicinal potential due to its remarkable pharmacological effects, safety, and edible characteristics. With the completion of the genome sequence and the advancement of efficient gene-editing technologies, coupled with the identification of gene functions in Cordyceps militaris, this fungus is poised to emerge as an outstanding strain for medicinal engineering applications. This review focuses on the development and application of genomic editing techniques, including Agrobacterium tumefaciens-mediated transformation (ATMT), PEG-mediated protoplast transformation (PMT), and CRISPR/Cas9. Through the application of these techniques, researchers can engineer the biosynthetic pathways of valuable secondary metabolites to boost yields; such metabolites include cordycepin, polysaccharides, and ergothioneine. Furthermore, by identifying and modifying genes that influence the growth, disease resistance, and tolerance to environmental stress in Cordyceps militaris, it is possible to stimulate growth, enhance desirable traits, and increase resilience to unfavorable conditions. Finally, the green sustainable industrial development of C. militaris using agricultural waste to produce high-value-added products and the future research directions of C. militaris were discussed. This review will provide future directions for the large-scale production of bioactive ingredients, molecular breeding, and sustainable development of C. militaris.
1. Introduction
Cordyceps militaris, a well-known edible and medicinal fungus, is considered to be a valuable mushroom used in traditional Chinese medicines and health supplements, which was approved by China’s Ministry of Health as a new resource of food in 2009. C. militaris primarily parasitize the pupae and larvae of Lepidoptera insects, which can penetrate the host’s body via the cuticle, spiracles, mouthparts, or other access points. The infection process of Cordyceps involves three stages: invasion, parasitism (development of the fungus before the death of the insect), and saprophytism (growth of the fungus after the host’s death) [1]. C. militaris has many bioactive constituents, like cordycepin, polysaccharide, cordyceps acid, and ergosterol, as well as multiple microelements [2,3], among which cordycepin (3′-deoxyadenosine), an adenosine analog, is one of the most valuable and extensively studied components; it boasts various pharmacological functions: antioxidant, anticancer, antitumor, antibacterial, and anti-inflammatory activity, and immune regulation [4,5,6,7,8]. Moreover, compared to Cordyceps sinensis, it grows more rapidly, requires less stringent cultivation conditions, and some of its key bioactive ingredients are even higher than Cordyceps sinensis, so C. militaris is widely used as an ideal substitute for C. sinensis and is of great research value. Traditionally, the most direct method for obtaining bioactive secondary metabolites like cordycepin from C. militaris is through extraction from wild or artificially cultivated fruit bodies [9,10]. However, due to the long growth cycle and strict environmental requirements, this process incurs high technical costs and also adds to the environmental burden of the industry.
In recent years, with the completion of the C. militaris whole-genome sequencing [11], research in molecular biology and genetics has been progressively deepening. Through genetic engineering, researchers can precisely edit the genome of C. militaris to enhance its ability to produce specific bioactive components, such as cordycepin [12,13]. In particular, CRISPR/Cas9 gene-editing technology enables the modification of multiple genes in both functional genomics research and strain breeding in C. militaris [14,15,16]. In addition, the construction of a high-production strain for secondary metabolites can be achieved by metabolic engineering and homologous/heterologous expression [17,18,19]. These technological advancements not only help to improve the biosynthetic efficiency of C. militaris but may also reveal its complex bioactive mechanisms, laying the foundation for the development of new drugs to treat various diseases.
Although C. militaris can be commercially cultivated on a large scale, it suffers from degradation problems similar to other edible fungi such as Flammulina velutipes, Lentinula edodes, and Pleurotus eryngii [20,21,22]. To date, the challenge of C. militaris degeneration remains unresolved, and its molecular mechanisms are not well understood [23,24]. Additionally, fungal diseases have become one of the bottlenecks in the scaled production of fruiting bodies, with “white mildew” disease caused by Calcarisporium cordycipiticola being the most damaging [25,26]. This not only leads to reduced yields of C. militaris but also raises concerns about pesticide contamination and product safety. Fortunately, the establishment of the first disease-resistant gene-editing breeding system based on CRISPR/Cas9 technology in C. militaris represents a seminal advance [27]. This development underscores the potential of gene-edited, disease-resistant cultivars as a cost-effective and environmentally sustainable strategy to significantly reduce losses attributed to agricultural diseases. Here, we present a review of the application of gene-editing techniques to improve the growth and metabolite production in C. militaris, and the potential of synthetic biology to optimize metabolic pathways and increase yield was discussed, alongside innovative approaches to combat industry challenges such as degradation and disease. Finally, the existing challenges and future research direction of C. militaris were discussed, aiming to promote the scientific development and technological innovation of the C. militaris industry.
2. The Genetic Manipulation Technology of Cordyceps militaris
2.1. Commonly Used and Efficient Genetic Transformation System
2.1.1. Agrobacterium tumefaciens-Mediated Transformation
The Agrobacterium tumefaciens-mediated transformation system for C. militaris was successfully established and optimized in 2011 [28], when Agrobacterium AGL-1 harboring the vector pATMT1 exhibited a transformation efficiency of 30–600 transformants per 1 × 105 conidia. Based on this, researchers utilized ATMT technology to create a mutant library of C. militaris and screened mutants using degenerate characteristics [29]. The crucial advantage of the ATMT method is the flexibility in choosing recipient cell types as starting materials for transformation. In particular, fungal spores serve as the most common material because millions of spores can be easily collected from a culture plate [30]. However, some challenges, including complicated and laborious procedures, low rates of homologous recombination, as well as a lot of transformant screening work, should be taken into consideration [31].
2.1.2. Polyethylene Glycol (PEG)-Mediated Protoplast Transformation
PEG is a kind of polymer with stable physicochemical properties, which can promote the fusion of exogenous DNA and protoplasts by interfering with the surface charge of the cell membrane or acting as an adhesive between DNA and cell membrane. This method was first successful in Saccharomyces cerevisiae [32] and was later applied to filamentous fungi. The preparation of protoplasts of C. militaris was discussed in several essays [14,16,33,34,35]. The Tns gene, which encodes a terpene synthase in C. militaris, was successfully knocked out using a combination of split-marker methods and PEG-mediated protoplast transformation techniques [33]. Moreover, the efficiency of PEG-mediated protoplast transformation in C. militaris was improved by adding the surfactant Triton X-100 used to enhance cell membrane permeability [15]. In addition, PEG-mediated protoplast transformation was markedly more efficient than ATMT transformation; the same plasmid was transformed by PEG-mediated protoplast transformation and ATMT; among the 38 transformants obtained by PEG-mediated protoplast transformation, 35 showed the predictable phenotype; however, only 7 were observed among the 70 transformants obtained by ATMT [16].
2.2. Selective Marker for the Detection of Positive Transformants
A stable and efficient genetic transformation system requires suitable screening markers, which can reduce false positives, improve screening efficiency, and avoid the loss of foreign genes in the process of purification and culture. Commonly used screening makers include auxotrophic markers and drug-resistant makers.
2.2.1. Auxotrophic Markers
An auxotrophic mutant strain refers to a mutant strain that loses the ability to synthesize a certain growth factor and can only grow in a medium with an exogenous addition of the growth factors. Auxotrophic screening restores the wildtype growth of recipient cells by transferring auxotrophic complementary genes into recipient cells. The auxotrophic markers currently used in C. militaris are pyrG (encoding orotic acid-5′-monophosphate decarboxylase) and hisB (encoding imidazole glycerophosphate dehydratase) [14,36]. So, we screen the positive transformants by adding uridine/uracil or histidine. Recently, the ΔpyrG ΔhisB dual auxotrophic mutant transformation system was constructed, and the knockout and complementation of the photoreceptor gene CmWC-1 were accomplished [36]. The newly constructed ATMT system with histidine and uridine/uracil auxotrophic markers provides a promising tool for genetic modifications in the medicinal fungus C. militaris.
2.2.2. Drug-Resistance Markers
Resistance screening refers to the transfer of resistance genes into recipient cells to make the transformants produce drug resistance, which can be screened at a certain drug concentration. Hygromycin, G418, phleomycin, nourseothricin, Basta, 5-fluorocytosine, and 5-FOA are commonly used in the genetic manipulation of fungi. The growth of conidia and mycelia was completely inhibited at a Basta concentration of 0.4 g/L and a 5-FOA concentration of 0.1 g/L, respectively [14]. After other resistant plasmids were transferred into recipient cells, related resistance was added to the plate for screening [33,37,38].
2.3. Split-Marker Approach
The split-marker approach was established to improve the frequency of homologous integration and gene knockout [33]. Deletion cassettes with different forms of the selectable marker (split or linear) have been made using single-joint PCR (SJ-PCR) and double-joint PCR (DJ-PCR). Then, the linear and split-marker deletion cassettes were constructed and introduced into C. militaris protoplasts by PEG-mediated transformation. The gene-targeting specificity and precision were enhanced when using split-marker fragments compared to linear deletion cassettes. The transformation of split-marker fragments led to a greater efficacy in achieving targeted gene disruption compared to the use of linear deletion cassettes. Moreover, the mutants could maintain resistance to glufosinate ammonium. While this approach has significantly increased the frequency of homologous recombination, the restricted selection marker choices and the enduring presence of resistance pose significant challenges in subsequent genome editing [39].
2.4. CRISPR-Cas9 Gene-Editing Technology
The clustered regularly interspaced short palindromic repeats (CRISPR) and CRISPR-associated protein 9 (Cas9) system represents a major breakthrough in the field of biology and medicine in recent years, providing an efficient and flexible method for accurately editing genes. The CRISPR editing system was first established in C. militaris in 2018, the pyrG gene was successfully edited by expressing the codon-optimized Cas9 gene with the newly discovered promoter Pcmlsm3 and terminator Tcmura3; however, the efficiency was only 11.76% [14]. The editing efficiency of Cmura5 reached 100% when an optimized ribonucleoprotein (RNP)-based method was employed [15]. Subsequently, marker-free multiplex gene-editing and a large DNA fragment deletion technique were developed by mining endogenous tRNA processing elements, introducing the autonomously replicating AMA1 plasmid and homologous templates [40]. Additionally, using the AMA1-based autonomously replicating CRISPR-Cas9 gene-editing system, the blue light receptor genes WC-1 and VVD in C. militaris were edited with efficiencies of 55.1% and 89%, respectively [16]. The application of CRISPR-Cas9 technology has accelerated the development of molecular breeding in C. militaris and improved the production efficiency of bioactive fungal strains. The gene transformation method of C. militaris is shown in Table 1 and Figure 1.

Table 1.
Genetic manipulation technologies of C. militaris.
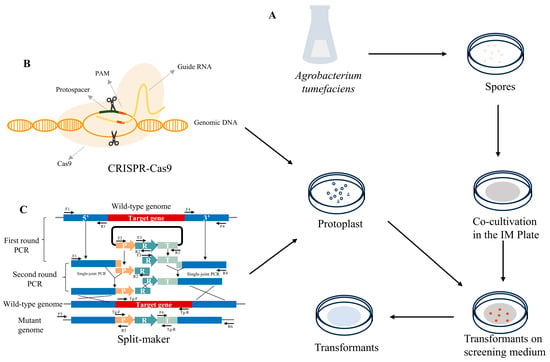
Figure 1.
Genetic manipulation technologies of C. militaris. (A) Agrobacterium-mediated transformation (ATMT) process; the plasmid was introduced into Agrobacterium and cocultivated with C. militaris spores in the IM plate (induction solid medium used for cocultivation), then the transformants (mutants) were obtained. (B) CRISPR-Cas9 editing technology. The plasmid constructed by CRISPR-Cas9 was introduced into C. militaris protoplasts through the protoplast-mediated transformation (PMT) method, then the transformants were screened on corresponding resistance plates. (C) Split-marker approach. Split-marker deletion cassettes constructed using single-joint PCR (SJ-PCR) and double-joint PCR (DJ-PCR) were introduced into C. militaris protoplasts, then the transformants were screened on corresponding resistance plates. All transformants (mutans) were confirmed by using polymerase chain reaction (PCR) or the detection of a reporter gene.
3. Increasing the Content of Secondary Metabolites of C. militaris by Genetic Engineering
C. militaris contains various active secondary metabolites, like cordycepin (COR), cordyceps polysaccharides, ergothioneine, carotenoid, and other active substances. With the publication of the whole-genome sequence of C. militaris and the development of genomics, metabolomics, transcriptomics, and other methods and technologies, the biosynthetic pathways of these metabolites have been gradually illustrated, which is conducive to increasing the content of secondary metabolites of C. militaris by genetic engineering.
3.1. The Enhancement of COR Production
To create industrial cell factories that produce high amounts of cordycepin, it is necessary to investigate the gene cluster responsible for cordycepin synthesis. Despite the identification of cordycepin in C. militaris in 1950 [42], the deciphering of its biosynthetic pathway has been protracted due to the absence of sophisticated genome analysis techniques. It was not until 2011 that the adenosine metabolic pathways in C. militaris and the genes involved in cordycepin synthesis were predominantly uncovered through genomics and transcriptomics studies. In 2017, Xia’s research team identified four highly conserved protein-coding genes by evaluating a vast number of orthologous proteins between C. militaris and Aspergillus nidulans, designated as cns1-cns4 (CCM_04436-CCM_04439), which were associated with adenosine metabolism in C. militaris [43]. The latest insights into the cordycepin biosynthesis pathways have been compiled in Figure 2. The results showed that the initial precursor of cordycepin biosynthesis pathway was glucose [44,45]. Glucose is transported into the interior of C. militaris, where it is phosphorylated to form glucose-6-phosphate (G6P), which is diverted into two pathways: a portion enters the pentose phosphate pathway, while the remainder is converted to fructose-6-phosphate (F6P) by phosphoglucose isomerase. F6P subsequently enters the tricarboxylic acid cycle (TCA) to provide energy for the growth of cells. The G6P that enters the pentose phosphate pathway is transformed into ribose-5-phosphate, which is converted into inosine monophosphate (IMP) through a series of reactions. IMP is then converted into adenylosuccinate and transformed into cordycepin under the catalysis of the cordycepin substrate channel.
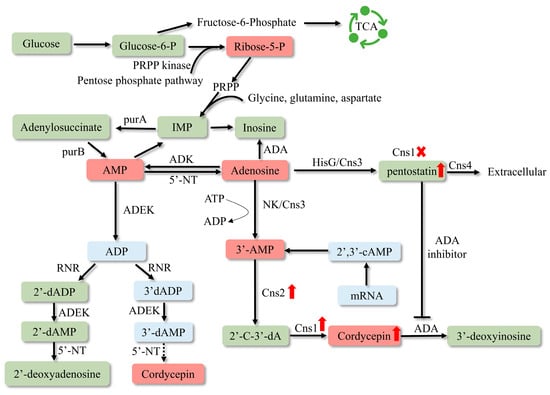
Figure 2.
Biosynthetic pathway of cordycepin. PRPP—5-phosphoribosyl 1-pyrophosphate; TCA—tricarboxylic acid cycle; IMP—inosine monophosphate; RNR—ribonucleotide reductases; ADEK—adenylate kinase; NT5E—5′-nucleotidase; 2′-C-3′-dA—2′-carbonyl-3′-deoxyadenosine; NK—N-terminal nucleoside kinase; HisG—C-terminal HisG family of ATP phosphoribosyltransferases. The deletion of cns1 causes the metabolic flow to be inclined towards pentostatin synthesis, thus pentostatin content has been enhanced.
Zhang et al. investigated the function of ribonucleotide reductases (RNRs), including the two subunits RNRL and RNRM, in the production of cordycepin (COR) by enhancing the expression of these genes in C. militaris [13]. The results showed that the concentration of cordycepin (COR) was significantly increased in C. militaris cells where the RNRM gene was overexpressed, reaching 3.750 mg/g. In contrast, the COR content in the cells overexpressing the RNRL gene only saw a minor increase to 2.600 mg/g, which was not significantly different from the wildtype level of 2.500 mg/g. It was suggested that RNRM might directly contribute to the biosynthesis of COR by hydrolyzing adenosine, although the precise mechanism by which RNRM facilitates COR synthesis remains to be elucidated. Research indicates that low-oxygen environments favor the synthesis and secretion of cordycepin, and the hypoxic environment within the insect hemocoel may also contribute to the higher cordycepin content in fruiting bodies growing on pupae [46]. Improving the low-oxygen environment by overexpressing sterol regulatory element-binding proteins (SREBPs) significantly increases cordycepin content [47]. Overexpressing enzymes involved in cordycepin biosynthesis, including the adenylosuccinate synthase, adenylosuccinate lyase, and 5′-nucleotidase genes, led to increased production in recombinant strains compared to the wildtype. Among 24 recombinant strains, the CM-adss-5 strain had the highest cordycepin production, and the extracellular and intracellular cordycepin contents were 1119.75 ± 1.61 and 65.56 ± 0.97 mg/L, 1.26 and 2.61 times those of C. militaris WT [48]. In addition, cordycepin can also be produced by heterologous expression of the key genes of cordycepin synthesis, such as overexpression of Cns1 and Cns2 in Aspergillus oryzae; through optimized fermentation conditions, a cordycepin yield of 564.64 ± 9.59 mg/L/d has been achieved [49]. As a result of overproduction of cordycepin in Saccharomyces cerevisiae by cordycepin synthase screening and metabolic engineering, the COR titer reached 725.16 mg L−1 in a 5 L bioreactor [50]. During high-level de novo biosynthesis of cordycepin by systems metabolic engineering in Yarrowia lipolytica, the production of cordycepin was achieved at 3588.59 mg/L from glucose [51,52]. By elucidating the biosynthesis and regulatory mechanisms of COR, genetically modifying the genes related to COR biosynthesis in C. militaris will become a viable approach to enhance the production of COR in this organism.
3.2. The Enhancement of Cordyceps Polysaccharides Content
The activity of polysaccharides is determined by their composition of monosaccharides, the type of glycosidic links, and the level of polymerization [53]. Cordyceps polysaccharides are mainly composed of glucans connected by glycosidic bonds, which include (1 → 3)- and (1 → 6)-β or (1 → 3)-, (1 → 4)-, and (1 → 6)-α-glucans [54,55]. Investigations into the biosynthesis of polysaccharides in various organisms, such as the overexpression of crucial polysaccharide biosynthetic genes [56], the co-expression of multiple genes [57], and the inhibition of polysaccharide synthesis pathways [58], represent all viable strategies to enhance yield. The proposed biosynthetic pathway of polysaccharides from C. militaris was shown in Figure 3.
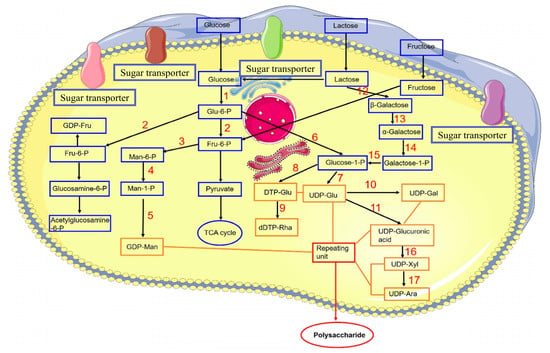
Figure 3.
Proposed biosynthetic pathway of polysaccharides from C. militaris [55]. 1. Glucokinase. 2. Phosphoglucose isomerase. 3. Phosphate mannose isomerase. 4. Phosphomannose mutase. 5. GDP-mannose pyrophosphorylase. 6. α-Phosphoglucose mutase. 7. UDP glucose pyrophosphorylase. 8. dTDP-glucose pyrophosphorylase. 9. Rhamnose synthase. 10. UDP-glucose-4-epimerase. 11. UDP-glucose dehydrogenase. 12. β-Galactosidase. 13. Galactosidase. 14. Galactokinase. 15. UDP-glucose–hexose-1-phosphate uridylyltransferase. 16. UDP-xylose synthase. 17. UDP-xylose-4-epimerase.
The pathway of fungal polysaccharide biosynthesis encompasses three stages: the synthesis of nucleotide sugar precursors, the arrangement of these precursors into repeating units, and the subsequent polymerization of these units [59]. Therefore, enhancing the catalytic efficiency of these enzymes through overexpression is anticipated to boost the yield of polysaccharides. Researchers overexpressed four genes—gk (glucokinase), pgm (phosphoglucomutase), ugp (UDP-glucose-pyrophosphosphprylase), and upgh (UDP-glucose-6-dehydrogenase)—in C. militaris, with the mutant CM-pgm producing 4.63 ± 0.23 g/L of polysaccharide, a 34.89% increase over the wildtype’s 3.43 ± 0.26 g/L. Furthermore, the combined overexpression of pgm and upgh in the engineered strain led to a polysaccharide yield of 6.11 ± 0.21 g/L, a 78.13% improvement over the wildtype. The study showed that pgm is an important regulatory gene that affects EPS biosynthesis of C. militaris [37]. Production of polysaccharides in C. militaris is intricate and multifaceted, involving a wide array of structural forms and gene clusters. Additional investigation is required to delve into the mechanisms governing polysaccharide biosynthesis and its regulation.
3.3. The Enhancement of Other Active Secondary Metabolites Production
Ergothioneine, one of the ingredients with great antioxidant and anti-inflammatory activity [60] in C. militaris, is a natural sulfur-containing thiol molecule, and has been regarded as longevity vitamins these years [61,62]. However, ergothioneine could not be synthesized by plants and animals, including humans, and the primary dietary source of ergothioneine for humans is mushroom-based foods [63]. To enhance the commercial yield of ergothioneine, engineered strains featuring different ergothioneine synthetases have been developed in model organisms such as Escherichia coli [64,65,66], Saccharomyces cerevisiae [67,68], Aspergillus oryzae [69], etc. [70,71]. However, despite the intricate optimization and high costs associated with fermentation, the ergothioneine content is not high enough to fulfill the market’s needs. The ergothioneine content in the wildtype C. militaris fruiting body ranged from 382 to 799 mg/kg of dry weight (DW), and 130 ± 11 mg/kg DW mycelia. Chen et al. identified ergothioneine synthetase genes in mushrooms and engineered them into C. militaris to establish a novel ergothioneine biosynthetic route. By integrating this route into the C. militaris genome, they achieved a significant boost in the production of ergothioneine, with the engineered strain producing up to 2.5 g/kg dry weight [72].
Carotenoids, including lutein, zeaxanthin, and cordyxanthin, are a class of pigments that possess a common terpenoid backbone, which can be used as food, dietary supplements, and cosmetic additives [73,74]. Although the biosynthetic pathway of C. militaris carotenoids is still unknown, some genes have been identified to be closely related to carotenoid synthesis. The color of C. militaris colonies can transform from white to yellow or orange when exposed to light, a phenomenon attributed to the accumulation of carotenoids. WC-1 is an essential component of all known blue light responses [75,76,77]. The absence of the CmWC-1 gene leads to abnormal color change and impaired synthesis of carotenoids [78]. Similarly, the deletion of the photoreceptor gene VVD also impaired the synthesis of carotenoids [79]. By analyzing transcriptome data of C. militaris 10 cultured under dark and light conditions, the CCM_06728 and CCM_09155 genes might be involved in the biosynthesis of carotenoids in C. militaris, and the absence of terpenoid synthase gene (Tns) in C. militaris led to a significant decrease in carotenoid production [80]. To obtain higher yield of carotenoid, it is urgent to study the biosynthetic pathway and related genes of C. militaris carotenoids in order to improve the content of carotenoids by genetic engineering.
Pentostatin is a potent inhibitor of adenosine deaminase (ADA) [81], which was approved by the FDA in 1991 and became a commercial drug for the treatment of hairy cell leukemia (under the brand name Nipent). During the majority of the growth cycle of C. militaris, pentostatin functions intracellularly by inhibiting the activity of adenosine deaminase, ensuring the synthesis of cordycepin and its low abundance presence within the cell [43]. Pentostatin shares the same precursor with cordycepin [43]; its production can be enhanced by deleting cns1 [82]. With the discovery of biosynthetic pathways of active ingredients in C. militaris, overexpression of key synthetic genes or knockout genes of competing pathways by genetic engineering technology is expected to increase the yield of active substances.
4. Promoting the Growth and Development of C. militaris by Genetic Engineering
The cultivation of C. militaris involves four key developmental stages: mycelial culture, color induction, primordia simulation, and fruiting body formation [29,83]. Using genetic engineering technology to regulate the growth and development genes of C. militaris, improve the growth rate, shorten the growth cycle, and reduce the culture cost will be conducive to the development of this industry. Recently, researchers have made significant progress in the identification and functional exploration of genes related to the traits of C. militaris (Table 2).
Hydrophobins are a unique class of small molecular secretory proteins in fungi and play a crucial role in their growth and development. C. militaris includes four hydrophobins. CmHYD1 and CmHYD2 are class II hydrophobins, while CmHYD3 and CmHYD4 are class I hydrophobins. Gene knockout, complementation, and overexpression studies have revealed that CmHYD1 positively regulates mycelial growth, hydrophobicity, and resistance to osmotic and hydrogen peroxide stress, as well as fruiting-body development, which regulates conidiospore formation by modulating the transcription levels of the circadian clock protein gene Cmfrq and the velvet factor gene Cmvosa [84]. In contrast, the knockout of the CmHYD4 gene promotes conidiospore formation and increases the density of fruiting bodies, enhancing the content of carotenoids and adenosine [85]. The first identified hydrophobin was SC3 from the Schizophyllum commune, which is essential for the formation of aerial mycelium [86]. Reports on hydrophobins in macrofungi also exist, for instance, nine hydrophobin genes were identified in Tricholoma vaccinum, which play an important role in the growth of aerial hyphae and fruiting-body development [87]. In Flammulina filiformis, Hyd9 was confirmed to play a significant role in the aerial hyphae and fruiting body formation through RNAi and overexpression [88].
Lectins are a class of proteins or glycoproteins that are not derived from the immune system and are widely found in plants, animals, and fungi [89]. Research has confirmed that lectins are closely related to the growth and development of macrofungi. For example, the application of Agrocybe aegerita lectin on the surface of normally growing mycelium can significantly promote the differentiation of mycelium and the formation of primordia in Agrocybe aegerita and Auricularia polytricha [90]. Additionally, the activity of Grifola frondosa lectin is highest in fruiting bodies and lower in mycelium and primordia [91]. Four types of lectins have been identified in C. militaris [38,92,93,94], among which the CmLec3 gene is expressed at a level five times higher in primordia than in mycelium and 1.35 times higher in fruiting bodies than in mycelium, which may be associated with the development of fruiting bodies [94]. CmLec4 may regulate various functions within C. militaris by interacting with polysaccharides and is involved in host invasion and the formation of fruiting bodies [38]. Compared to plants and animals, research on lectins in large fungi is relatively lagging, and further studies on the mechanisms of action of lectins at the genetic level and how they promote fungal development are needed.
Light is also a crucial environmental factor that significantly affects the growth, development, and metabolism of C. militaris. Seven types of photoreceptors have been identified in C. militaris, including white collar (WC-1), WC-2, Drosophila–Arabidopsis–Synechocystis–human-type cryptochromes (CRY-DASH), CRY-2, cyclobutane pyrimidine dimer photolyase (CPD), VIVID (VVD), and phytochrome (PHY) [11]. The deletion of Cry-DASH in C. militaris results in reduced conidial numbers but significantly increased levels of carotenoids and cordycepin, and the light-induced expression of Cmcry-DASH is dependent on the expression of Cmwc-1 [95]. Studies suggest that CRY-DASH can repair damaged single-stranded and double-stranded DNA and its bound CPD, but this function in C. militaris is not yet clear [96,97,98]. The regulation of growth and development of C. militaris is a complex process involving multiple genes and metabolic networks, so the regulation mechanism of growth and development needs to be further explored.
Table 2.
A list of genes that regulate the development of mycelium and fruiting body in Cordyceps militaris.
Table 2.
A list of genes that regulate the development of mycelium and fruiting body in Cordyceps militaris.
| Gene Name | Accession Number | Annotation | Function | Reference |
|---|---|---|---|---|
| Cry-DASH | CCM_00774 | Cryptochrome DASH | Fruiting-body development regulation | [95] |
| WC-1 | AGO64764 | Blue light receptor white collar 1 | Mycelium growth and fruiting-body development regulation | [78] |
| VVD | CCM_04514 | GATA transcription factor LreA | Fruiting-body development regulation | [79] |
| Lec3 | CCM_01589 | Lectin family integral membrane protein, putative | Fruiting-body development regulation | [94] |
| Lec4 | CCM_03832 | Ricin B-related lectin | Fruiting-body development regulation | [38] |
| Hyd1 | CCM_03537 | Hydrophobin 2 | Fruiting-body development regulation | [84] |
| Hyd4 | CCM_07964 | Hydrophobin | Fruiting-body development regulation | [85] |
| fhp | CCM_05119 | Flavohemoprotein | Fruiting-body development regulation | [99] |
| Snf1 | CCM_05552 | Carbon catabolite derepressing protein kinase | Fruiting-body development regulation | [100] |
| Chi1 | CCM_08231 | Class V chitinase, putative | Fruiting-body development regulation | [101] |
| Chi4 | CCM_04817 | Class V chitinase | Fruiting-body development regulation | [101] |
| crf1 | CCM_07998 | Fungal-specific transcription factor | Fruiting-body development regulation | [102] |
5. Breeding for Disease Resistance of C. militaris Using CRISPR/Cas9
The “white mildew” disease caused by Calcarisporium cordycipiticola significantly affects the quality and yield of C. militaris in industrial production [25], which invades C. militaris through the gaps in the fruiting body mycelium without forming specialized infection structures, leading to cell wall dissolution, cell deformation, loss of organelles, and ultimately cell rupture [103]. Further analysis revealed that iron is an important virulence factor for the pathogen, and the genes Cmhsp78, Cmhsp70, and Cmhyd1 in C. militaris respond rapidly during the infection process and could serve as potential resistance genes for breeding disease-resistant strains of C. militaris [26]. Building on this foundation, researchers have utilized CRISPR/Cas9 gene-editing technology to overexpress the resistance gene Cmhyd1, obtaining a C. militaris strain with significantly enhanced resistance to this disease; this is the first disease-resistant gene-editing breeding system in mushrooms [27].
Fungi have specific temperature requirements during the growth phase, and temperatures that are too high or too low affect them. For instance, the fruiting-body growth of Agaricus bisporus is severely reduced at high temperatures of 33 °C, with a notable reduction in cap diameter and biomass [104]. The growth and hyphal morphology of Tuber borchii are affected at temperatures of 28 °C or 34 °C [105], while 42 °C inhibits the growth of Ganoderma lucidum mycelium, reduces branching, and induces the accumulation of ganoderic acid within the mycelium. The optimum growth temperature of C. militaris is 20–22 °C, and exposure to 25 °C during the later stages of growth can increase the content of cordycepin and carotenoids, whereas mycelial growth is severely inhibited at 30 °C [106]; therefore, the construction of heat-resistant strains is of great significance to the development of C. militaris industry. However, the specific temperature-stress response mechanisms in C. militaris remain unclear. Studies have suggested that introducing small heat shock protein genes into organisms can enhance their thermotolerance [107,108,109], which provide a reference for studying the heat stress response mechanisms in C. militaris.
6. Resource Utilization of Agricultural Waste and Sustainable Production of C. militaris
Agricultural waste refers to the organic substances discarded during the entire process of agricultural production, including the waste generated in agricultural production, livestock, and poultry farming. The spent mushroom substrate (SMS) is the residual biomass generated after harvesting the fruit bodies of edible/medicinal fungi, which are rich in lignocellulose, protein, and other essential elements [110,111]. They are generally disposed of by landfilling, open burning, and composting with animal manure, which are prone to causing environmental issues such as soil contamination, and air and water pollution [112,113]. Hence, more effective and efficient utilization of SMS is in demand for environmental protection, waste recycling, and sustainable development of agricultural resources and environment [114,115]. Although SMS has been used to produce industrial products such as packaging and building materials, biofuels, chemicals, and enzymes [116,117], these traditional recycling methods are time-consuming, with low conversion rates and added value.
C. militaris, an edible and medicinal fungus, contains an active substance pentostatin approved by the FDA as a first-line drug for the treatment of leukemia. Currently, pentostatin is produced by large-scale fermentation of Streptomyces antibioticus [118], but the yield is low. Since C. militaris lacks lignocellulose-degrading enzymes, the researchers introduced an exogenous optimized cellulase system, including XynA (xylanase from Bacillus sp. KW1), TrCBH1 (cellobiohydrolase from Trichoderma reesei), and CtCBH1 (cellobiohydrolase from the thermophilic fungus Chaetomium thermophilum), that can effectively degrade spent mushroom substrates (SMS) into glucose and other simple fermented sugars, thus enabling C. militaris to utilize the abundant lignocellulose produce the high-value anticancer drug pentostatin (Figure 4). Compared to the wildtype, the content of pentostatin in the engineered fungus significantly increased from 23.75% (CmctC) to 96.20% (CmTX). The big increase in output could be attributed to the multifunctional XynA and high-expressed TrCBH1, and the combined action of these enzymes could potentially enhance the capacity for hydrolyzing the fungal cell wall. Consequently, the destruction of the cell wall could stimulate the synthesis of secondary metabolites, akin to the stress response. There is a precursor competition relationship between the biosynthesis of pentostatin and cordycepin. To shift the metabolic flow towards the production of pentostatin and away from the rival biosynthesis pathway of cordycepin, the cns1 gene involved in cordycepin biosynthesis was removed in CmTX mutant, leading to pentostatin titers exceeding 600 mg/L in submerged fermentation [82]. This research is the first to turn edible fungi into cell factories for anticancer drugs and successfully transform agricultural waste into high-value-added anticancer drugs.
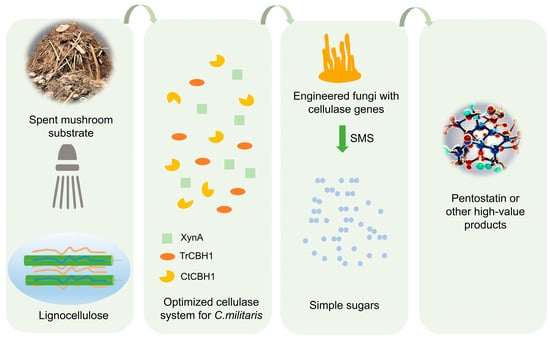
Figure 4.
Engineered C. militaris with cellulase genes convert spent mushroom substrates (SMS) to valuable products. XynA—xylanase from Bacillus sp. KW1; TrCBH1—cellobiohydrolase from Trichoderma reesei; CtCBH1—cellobiohydrolase from the thermophilic fungus Chaetomium thermophilum. The optimized cellulase system was introduced into C. militaris, which was able to degrade SMS into monosaccharides, thereby promoting its own growth and increasing the content of the target product.
7. Conclusions and Expectation
Cordyceps militaris, a fungus renowned for its diverse bioactive compounds and its significance in both culinary and medicinal practices, continues to attract significant attention in the biotechnological and pharmaceutical industries. This article offers a comprehensive overview of the progress in genomic editing techniques, including ATMT, PEG, and CRISPR/Cas9. The application of these techniques in Cordyceps militaris is also summarized, including regulating key genes in the synthesis pathway of secondary metabolites to increase yield, regulating genes affecting growth and development to promote growth, reducing culture costs, and enhancing the expression of disease-resistant genes to obtain disease-resistant strains. The development of pathogen-resistant strains of C. militaris is an area ripe for exploration, as it would significantly mitigate the risks associated with cultivation and ensure sustainable production. Additionally, through heterologous expression of the cellulase gene, Cordyceps militaris can degrade agricultural wastes to produce high-value-added drugs. This sustainable production method is crucial for achieving a balance between environmental protection and economic growth, which would not only reduce the strain on natural resources but would also present a viable solution for large-scale cultivation. Moreover, the fusion antimicrobial peptide gene of Magainin II-Cecropin B was successfully expressed in C. militaris, and the resulting recombinant antimicrobial peptide, with its broad-spectrum antibacterial properties, particularly against Staphylococcus aureus, highlights the potential of C. militaris as a versatile platform for the production of therapeutic and functional biomolecules [35]. In the future, the combination of gene editing and synthetic biology holds immense potential for the creation of novel C. militaris strains capable of producing an expanded array of bioactive substances. Furthermore, the integration of high-throughput sequencing and omics technologies will undoubtedly deepen our knowledge of the C. militaris genome and transcriptome and speed up the discovery of novel genes and regulatory networks, thereby providing a broader palette of genetic modifications to enhance the production yield and efficiency of bioactive components. In conclusion, these advancements are poised to cater to the increasing demands of the pharmaceutical, health, and industrial sectors. Continued research on and development of C. militaris will undoubtedly lead to groundbreaking discoveries and applications in various fields, further solidifying its status as a valuable resource in the biomedical and industrial sectors.
Author Contributions
Y.H. conceptualized this paper and wrote it. Y.W. participated in revising the manuscript. J.S. participated in revising the figures and table. M.M. and Y.X. collected the references. B.Z. reviewed this paper. All authors have read and agreed to the published version of the manuscript.
Funding
The Key Special Projects of the National Key Research and Development Plan (2021YFA1301302), Project of the Department of Science and Technology of the Jiangxi Province (20213AAG02020), the National Natural Science Foundation of China (32200606), Self-made Experimental Instruments and Equipment Project of Shenzhen Technology University (JSZZ202301021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Buenz, E.J.; Bauer, B.A.; Osmundson, T.W.; Motley, T.J. The traditional Chinese medicine Cordyceps sinensis and its effects on apoptotic homeostasis. J. Ethnopharmacol. 2005, 96, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.D. Biotechnological production and applications of Cordyceps militaris, a valued traditional Chinese medicine. Crit. Rev. Biotechnol. 2015, 35, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.T.; Yang, Y.C.; Li, Y.H.; Mau, J.L.; Wasser, S.P. Chemical Composition and Nutritional and Medicinal Value of Fruit Bodies and Submerged Cultured Mycelia of Culinary-Medicinal Higher Basidiomycetes Mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Cordyceps Medicinal Fungus and Its Nutraceutical and Therapeutic Potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, G.; Yan, W.; Lv, Y.; Wang, X.; Jin, G.; Cui, M.; Lin, Z.; Ren, X. Cordycepin Inhibits Cancer Cell Proliferation and Angiogenesis through a DEK Interaction via ERK Signaling in Cholangiocarcinoma. J. Pharmacol. Exp. Ther. 2020, 373, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Tania, M. Cordycepin and kinase inhibition in cancer. Drug Discov. Today 2023, 28, 103481. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.P.; Qian, Z.Q.; Wu, H. Enhancing cordycepin production in liquid static cultivation of Cordyceps militaris by adding vegetable oils as the secondary carbon source. Bioresour. Technol. 2018, 268, 60–67. [Google Scholar] [CrossRef]
- Li, X.; Liu, Q.; Li, W.J.; Li, Q.P.; Qian, Z.M.; Liu, X.Z.; Dong, C.H. A breakthrough in the artificial cultivation of Chinese cordyceps on a large-scale and its impact on science, the economy, and industry. Crit. Rev. Biotechnol. 2019, 39, 181–191. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, Z.Q.; Xue, Y.P.; Baker, P.J.; Wu, H.; Xu, F.; Teng, Y.; Brathwaite, M.E.; Zheng, Y.G. Biosynthetic Pathway Analysis for Improving the Cordycepin and Cordycepic Acid Production in Hirsutella sinensis. Appl. Biochem. Biotechnol. 2016, 179, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Y.X.; Tong, X.X.; Yokoyama, W.; Cao, J.; Wang, F.; Peng, C.; Guo, J.L. Overexpression of ribonucleotide reductase small subunit, RNRM, increases cordycepin biosynthesis in transformed Cordyceps militaris. Chin. J. Nat. Med. 2020, 18, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.X.; Wei, T.; Ye, Z.W.; Yun, F.; Kang, L.Z.; Tang, H.B.; Guo, L.Q.; Lin, J.F. Efficient CRISPR-Cas9 Gene Disruption System in Edible-Medicinal Mushroom Cordyceps militaris. Front. Microbiol. 2018, 9, 1157. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Xiao, M.; Chai, S.; Zhu, Z.; Wang, Y.; Zhou, Z. Efficient genome editing in filamentous fungi via an improved CRISPR-Cas9 ribonucleoprotein method facilitated by chemical reagents. Microb. Biotechnol. 2021, 14, 2343–2355. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.L.; Wang, X.P.; Liu, M.Q.; Wang, F.; Liu, Q.Z.; Dong, C.H. Efficient CRISPR/Cas9 system based on autonomously replicating plasmid with an AMA1 sequence and precisely targeted gene deletion in the edible fungus, Cordyceps militaris. Microb. Biotechnol. 2022, 15, 2594–2606. [Google Scholar] [CrossRef]
- Kang, N.; Lee, H.H.; Park, I.; Seo, Y.S. Development of High Cordycepin-Producing Cordyceps militaris Strains. Mycobiology 2017, 45, 31–38. [Google Scholar] [CrossRef]
- Kunhorm, P.; Chueaphromsri, P.; Chaicharoenaudomrung, N.; Noisa, P. Enhancement of cordycepin production from Cordyceps militaris culture by epigenetic modification. Biotechnol. Lett. 2022, 44, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.Y.; Yang, H.; Wang, C.; Liu, H.H.; Lu, X.Y.; Tian, Y. Microbial synthesis of cordycepin, current systems and future perspectives. Trends Food Sci. Technol. 2023, 132, 162–170. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, K.H.; Im, C.H.; Ali, A.; Lee, C.Y.; Kong, W.S.; Ryu, J.S. Identification of Degenerate Nuclei and Development of a SCAR Marker for Flammulina velutipes. PLoS ONE 2014, 9, e107207. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, S.W.; Ryu, J.S.; Lee, C.Y.; Ro, H.S. Isolation of a Variant Strain of Pleurotus eryngii and the Development of Specific DNA Markers to Identify the Variant Strain. Mycobiology 2014, 42, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.P.; Bian, Y.B.; Wang, J.J.; Wang, G.Z.; Ma, X.L.; Xu, Z.Y. Biological and Molecular Characteristics of a Novel Partitivirus Infecting the Edible Fungus Lentinula edodes. Plant Dis. 2017, 101, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Deng, C.H.; Zhang, L.Y.; Hu, K.I. Molecular analysis and biochemical characteristics of degenerated strains of Cordyceps militaris. Arch. Microbiol. 2017, 199, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Yin, J.; Zhang, B.; Li, Z.; Zhao, S.; Gui, Z. Genome-wide analysis of DNA methylation in subcultured Cordyceps militaris. Arch. Microbiol. 2019, 201, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.Z.; Dong, C.H.; Liu, X.Z.; Liu, J.K.; Hyde, K.D. Calcarisporium cordycipiticola sp nov., an important fungal pathogen of Cordyceps militaris. Phytotaxa 2016, 268, 135–144. [Google Scholar] [CrossRef]
- Liu, Q.; Dong, C.H. Dual Transcriptomics Reveals Interspecific Interactions between the Mycoparasite Calcarisporium cordycipiticola and Its Host Cordyceps militaris. Microbiol. Spectr. 2023, 11, e04800-22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Meng, G.; Wang, M.; Li, X.; Liu, M.; Wang, F.; Yang, Y.; Dong, C. Safe-Harbor-Targeted CRISPR/Cas9 System and Cmhyd1 Overexpression Enhances Disease Resistance in Cordyceps militaris. J. Agric. Food Chem. 2023, 71, 15249–15260. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.L.; Huang, C.H.; Cao, L.; Xie, C.H.; Han, R.H. Agrobacterium tumefaciens mediated transformation as a tool for insertional mutagenesis in medicinal fungus Cordyceps militaris. Fungal Biol. 2011, 115, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.L.; Qiu, X.H.; Han, R.C. Identification of the Genes Involved in the Fruiting Body Production and Cordycepin Formation of Cordyceps militaris Fungus. Mycobiology 2015, 43, 37–42. [Google Scholar] [CrossRef]
- Thai, H.D.; Nguyen, B.T.; Nguyen, V.M.; Nguyen, Q.H.; Tran, V.T. Development of a new Agrobacterium-mediated transformation system based on a dual auxotrophic approach in the filamentous fungus Aspergillus oryzae. World J. Microbiol. Biotechnol. 2021, 37, 92. [Google Scholar] [CrossRef]
- Koukaki, M.; Giannoutsou, E.; Karagouni, A.; Diallinas, G. A novel improved method for Aspergillus nidulans transformation. J. Microbiol. Methods 2003, 55, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Hinnen, A.; Hicks, J.B.; Fink, G.R. Transformation of yeast. Proc. Natl. Acad. Sci. USA 1978, 75, 1929–1933. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Ye, Z.; Yun, F.; Lin, J.; Guo, L.; Chen, B.; Mu, Z. Targeted Gene Deletion in Cordyceps militaris Using the Split-Marker Approach. Mol. Biotechnol. 2018, 60, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.W.; Ye, Z.W.; Yu, Y.H.; Lin, J.F.; Guo, L.Q.; Chen, B.X.; Tang, H.B.; Wei, T.; Chen, L.T.; Yun, F. The efficient genetic transformation of Cordycepsmilitaris by using mononuclear protoplasts. Sci. Hortic. 2019, 243, 307–313. [Google Scholar] [CrossRef]
- Zhang, M.; Shan, Y.L.; Gao, H.T.; Wang, B.; Liu, X.; Dong, Y.Y.; Liu, X.M.; Yao, N.; Zhou, Y.G.; Li, X.W.; et al. Expression of a recombinant hybrid antimicrobial peptide magainin II-cecropin B in the mycelium of the medicinal fungus Cordyceps militaris and its validation in mice. Microb. Cell Factories 2018, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.H.; Shang, Y.T.; Wang, L.H.; Tian, X.Q.; Tran, V.T.; Yao, L.H.; Zeng, B.; Hu, Z.H. Construction of a new Agrobacterium tumefaciens-mediated transformation system based on a dual auxotrophic approach in Cordyceps militaris. J. Microbiol. Biotechnol. 2024, 34, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Chen, P.; Yang, S.; Zhang, H. Homologous overexpression of genes in Cordyceps militaris improves the production of polysaccharides. Food Res. Int. 2021, 147, 110452. [Google Scholar] [CrossRef]
- Ono, A.; Suzuki, T.; Takeshima, Y.; Kashiwa, T.; Motoyama, T.; Choi, J.H.; Sato, C.; Konno, N.; Miyakawa, H.; Ogata, M.; et al. CmLec4, a lectin from the fungus Cordyceps militaris, controls host infection and fruiting body formation. Int. J. Biol. Macromol. 2022, 215, 303–311. [Google Scholar] [CrossRef]
- Zeng, J.; Zhou, Y.; Lyu, M.; Huang, X.; Xie, M.; Huang, M.; Chen, B.X.; Wei, T. Cordyceps militaris: A novel mushroom platform for metabolic engineering. Biotechnol. Adv. 2024, 74, 108396. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.X.; Xue, L.N.; Wei, T.; Wang, N.; Zhong, J.R.; Ye, Z.W.; Guo, L.Q.; Lin, J.F. Multiplex gene precise editing and large DNA fragment deletion by the CRISPR-Cas9-TRAMA system in edible mushroom Cordyceps militaris. Microb. Biotechnol. 2022, 15, 2982–2991. [Google Scholar] [CrossRef]
- Rachmawati, R.; Kinoshita, H.; Nihira, T. Establishment of transformation system in Cordyceps militaris by using integration vector with benomyl resistance gene. In Proceedings of the 3rd International Conference on Sustainable Future for Human Security (SUSTAIN), Kyoto Univ, Kyoto, Japan, 3–5 November 2012; pp. 142–149. [Google Scholar]
- Cunningham, K.G.; Manson, W.; Spring, F.S.; Hutchinson, S.A. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) Link. Nature 1950, 166, 949. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489.e4. [Google Scholar] [CrossRef]
- Vongsangnak, W.; Raethong, N.; Mujchariyakul, W.; Nguyen, N.N.; Leong, H.W.; Laoteng, K. Genome-scale metabolic network of Cordyceps militaris useful for comparative analysis of entomopathogenic fungi. Gene 2017, 626, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.-d.; Wang, W.; Zhong, J.-J. Enhancement of cordycepin production in submerged cultures of Cordyceps militaris by addition of ferrous sulfate. Biochem. Eng. J. 2012, 60, 30–35. [Google Scholar] [CrossRef]
- Kato, T.; Nishimura, K.; Suparmin, A.; Ikeo, K.; Park, E.Y. Effects of Cordycepin in Cordyceps militaris during Its Infection to Silkworm Larvae. Microorganisms 2021, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Z.S.; Bao, D.P.; Li, B.; Yin, X.; Wu, Y.Y.; Chen, H.Y.; Tang, G.R.; Li, N.Y.; Zou, G. Improving Hypoxia Adaption Causes Distinct Effects on Growth and Bioactive Compounds Synthesis in an Entomopathogenic Fungus Cordyceps militaris. Front. Microbiol. 2021, 12, 698436. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, P.; Xu, L.; Xu, D.; Hu, W.; Cheng, Y.; Yang, S. Construction of Cordycepin High-Production Strain and Optimization of Culture Conditions. Curr. Microbiol. 2022, 80, 12. [Google Scholar] [CrossRef] [PubMed]
- Jeennor, S.; Anantayanon, J.; Panchanawaporn, S.; Chutrakul, C.; Vongsangnak, W.; Laoteng, K. Efficient de novo production of bioactive cordycepin by Aspergillus oryzae using a food-grade expression platform. Microb. Cell Fact. 2023, 22, 253. [Google Scholar] [CrossRef]
- Wang, H.; Fu, X.; Zuo, X.; Zhang, C.; Lu, W. Overproduction of cordycepin in Saccharomyces cerevisiae by cordycepin synthase screening and metabolic engineering. AIChE J. 2024, 70, e18361. [Google Scholar] [CrossRef]
- Duan, X.Y.; Tian, Y.; Song, Z.Q.; Song, L.P.; Lin, W.B.; Wang, C.; Yang, H.; Lu, X.Y.; Ji, X.J.; Liu, H.H. High-level de novo biosynthesis of cordycepin by systems metabolic engineering in Yarrowia lipolytica. Bioresour. Technol. 2022, 363, 127862. [Google Scholar] [CrossRef]
- Song, Z.Q.; Lin, W.B.; Duan, X.Y.; Song, L.P.; Wang, C.; Yang, H.; Lu, X.Y.; Ji, X.J.; Tian, Y.; Liu, H.H. Increased Cordycepin Production in Yarrowia lipolytica Using Combinatorial Metabolic Engineering Strategies. Acs Synth. Biol. 2023, 12, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Methacanon, P.; Madla, S.; Kirtikara, K.; Prasitsil, M. Structural elucidation of bioactive fungi-derived polymers. Carbohydr. Polym. 2005, 60, 199–203. [Google Scholar] [CrossRef]
- Ujita, M.; Inoue, R.; Makino, Y.; Katsuno, Y.; Okumura, H. Binding specificity of the recombinant cytoplasmic domain of Cordyceps militaris β-1,3-glucan synthase catalytic subunit. Biosci. Biotechnol. Biochem. 2011, 75, 171–174. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wu, N.; Ge, X.; Yin, X.; Yang, L.; Chen, L.; Shao, R.; Xu, W. A review on polysaccharide biosynthesis in Cordyceps militaris. Int. J. Biol. Macromol. 2024, 260, 129336. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Bai, Y.; Dai, R.; Guo, X.; Liu, Z.H.; Yuan, S. Improved Polysaccharide Production by Homologous Co-overexpression of Phosphoglucomutase and UDP Glucose Pyrophosphorylase Genes in the Mushroom Coprinopsis cinerea. J. Agric. Food Chem. 2018, 66, 4702–4709. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zhong, Y.; Ding, L.; Liu, X.; Xu, S.; Guo, D.; Blennow, A.; Xue, J. Biosynthesis, structure and functionality of starch granules in maize inbred lines with different kernel dehydration rate. Food Chem. 2022, 368, 130796. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Zhao, F.; Han, S.; Zhang, Y. Enhanced rhamnolipids production in Pseudomonas aeruginosa SG by selectively blocking metabolic bypasses of glycosyl and fatty acid precursors. Biotechnol. Lett. 2020, 42, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, F.; Xu, Z.; Ding, Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules 2017, 22, 955. [Google Scholar] [CrossRef] [PubMed]
- Cheah, I.K.; Halliwell, B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2012, 1822, 784–793. [Google Scholar] [CrossRef]
- Beelman, R.B.; Kalaras, M.D.; Phillips, A.T.; Richie, J.P., Jr. Is ergothioneine a ‘longevity vitamin’ limited in the American diet? J. Nutr. Sci. 2020, 9, e52. [Google Scholar] [CrossRef]
- Ames, B.N. Prolonging healthy aging: Longevity vitamins and proteins. Proc. Natl. Acad. Sci. USA 2018, 115, 10836–10844. [Google Scholar] [CrossRef] [PubMed]
- Tsiantas, K.; Tsiaka, T.; Koutrotsios, G.; Siapi, E.; Zervakis, G.I.; Kalogeropoulos, N.; Zoumpoulakis, P. On the Identification and Quantification of Ergothioneine and Lovastatin in Various Mushroom Species: Assets and Challenges of Different Analytical Approaches. Molecules 2021, 26, 1832. [Google Scholar] [CrossRef] [PubMed]
- Kamide, T.; Takusagawa, S.; Tanaka, N.; Ogasawara, Y.; Kawano, Y.; Ohtsu, I.; Satoh, Y.; Dairi, T. High Production of Ergothioneine in Escherichia coli using the Sulfoxide Synthase from Methylobacterium strains. J. Agric. Food Chem. 2020, 68, 6390–6394. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Kawano, Y.; Satoh, Y.; Dairi, T.; Ohtsu, I. Gram-scale fermentative production of ergothioneine driven by overproduction of cysteine in Escherichia coli. Sci. Rep. 2019, 9, 1895. [Google Scholar] [CrossRef] [PubMed]
- Osawa, R.; Kamide, T.; Satoh, Y.; Kawano, Y.; Ohtsu, I.; Dairi, T. Heterologous and High Production of Ergothioneine in Escherichia coli. J. Agric. Food Chem. 2018, 66, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, S.A.; Darbani, B.; Zugaj, K.E.; Prabhala, B.K.; Biron, M.B.; Randelovic, M.; Medina, J.B.; Kell, D.B.; Borodina, I. Engineering the Yeast Saccharomyces cerevisiae for the Production of L- (+) -Ergothioneine. Front. Bioeng. Biotechnol. 2019, 7, 262. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Pan, H.Y.; Guo, L.Q.; Lin, J.F.; Liao, H.L.; Li, H.Y. Successful biosynthesis of natural antioxidant ergothioneine in Saccharomyces cerevisiae required only two genes from Grifola frondosa. Microb. Cell Fact. 2020, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Takusagawa, S.; Satoh, Y.; Ohtsu, I.; Dairi, T. Ergothioneine production with Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2019, 83, 181–184. [Google Scholar] [CrossRef]
- van der Hoek, S.A.; Rusnák, M.; Jacobsen, I.H.; Martínez, J.L.; Kell, D.B.; Borodina, I. Engineering ergothioneine production in Yarrowia lipolytica. FEBS Lett. 2022, 596, 1356–1364. [Google Scholar] [CrossRef]
- Pluskal, T.; Ueno, M.; Yanagida, M. Genetic and metabolomic dissection of the ergothioneine and selenoneine biosynthetic pathway in the fission yeast, S. pombe, and construction of an overproduction system. PLoS ONE 2014, 9, e97774. [Google Scholar] [CrossRef]
- Chen, B.X.; Xue, L.N.; Wei, T.; Ye, Z.W.; Li, X.H.; Guo, L.Q.; Lin, J.F. Enhancement of ergothioneine production by discovering and regulating its metabolic pathway in Cordyceps militaris. Microb. Cell Fact. 2022, 21, 169. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and torularhodin: “new” fungal carotenoids for industry? Microb. Cell Factories 2018, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.-H.; Hao, Y.-F.; Li, Y.-M.; Liang, Y.-J.; Jiang, J.-G. Inhibiting Lycopene Cyclases to Accumulate Lycopene in High β-Carotene-Accumulating Dunaliella bardawil. Food Bioprocess. Technol. 2016, 9, 1002–1009. [Google Scholar] [CrossRef]
- Linden, H.; Ballario, P.; Macino, G. Blue light regulation in Neurospora crassa. Fungal Genet. Biol. 1997, 22, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Talora, C.; Franchi, L.; Linden, H.; Ballario, P.; Macino, G. Role of a white collar-1-white collar-2 complex in blue-light signal transduction. Embo J. 1999, 18, 4961–4968. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Verma, S.; Corrochano, L.M. A glimpse into the basis of vision in the kingdom Mycota. Fungal Genet. Biol. 2010, 47, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Guo, M.M.; Yang, H.J.; Guo, S.P.; Dong, C.H. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, F.; Yang, Y.; Wang, Y.; Dong, C. CmVVD is involved in fruiting body development and carotenoid production and the transcriptional linkage among three blue-light receptors in edible fungus Cordyceps militaris. Environ. Microbiol. 2020, 22, 466–482. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.W.; Zhao, Y.; Tang, H.B.; Ye, Z.W.; Wei, T.; Lin, J.F.; Guo, L.Q. Transcriptome Analysis of Cordyceps militaris Reveals Genes Associated with Carotenoid Synthesis and Identification of the Function of the Cmtns Gene. Front. Microbiol. 2019, 10, 2105. [Google Scholar] [CrossRef]
- Rottenberg, M.E.; Masocha, W.; Ferella, M.; Petitto-Assis, F.; Goto, H.; Kristensson, K.; McCaffrey, R.; Wigzell, H. Treatment of African trypanosomiasis with cordycepin and adenosine deaminase inhibitors in a mouse model. J. Infect. Dis. 2005, 192, 1658–1665. [Google Scholar] [CrossRef]
- Zou, G.; Li, B.; Wang, Y.; Yin, X.; Gong, M.; Shang, J.J.; Wei, Y.J.; Li, X.L.; Bao, D.P. Efficient conversion of spent mushroom substrate into a high value-added anticancer drug pentostatin with engineered Cordyceps militaris. Green. Chem. 2021, 23, 10030–10038. [Google Scholar] [CrossRef]
- Lian, T.T.; Yang, T.; Liu, G.J.; Sun, J.D.; Dong, C.H. Reliable reference gene selection for Cordyceps militaris gene expression studies under different developmental stages and media. Fems Microbiol. Lett. 2014, 356, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, F.; Liu, M.; Dong, C. Hydrophobin CmHYD1 Is Involved in Conidiation, Infection and Primordium Formation, and Regulated by GATA Transcription Factor CmAreA in Edible Fungus, Cordyceps militaris. J. Fungi 2021, 7, 674. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, M.; Dong, C. Hydrophobin Gene Cmhyd4 Negatively Regulates Fruiting Body Development in Edible Fungi Cordyceps militaris. Int. J. Mol. Sci. 2023, 24, 4586. [Google Scholar] [CrossRef] [PubMed]
- vanWetter, M.A.; Schuren, F.H.J.; Schuurs, T.A.; Wessels, J.G.H. Targeted mutation of the SC3 hydrophobin gene of Schizophyllum commune affects formation of aerial hyphae. Fems Microbiol. Lett. 1996, 140, 265–269. [Google Scholar] [CrossRef]
- Sammer, D.; Krause, K.; Gube, M.; Wagner, K.; Kothe, E. Hydrophobins in the Life Cycle of the Ectomycorrhizal Basidiomycete Tricholoma vaccinum. PLoS ONE 2016, 11, e0167773. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X.; Chen, R.L.; Yan, J.J.; Long, Y.; Tong, Z.J.; Song, H.B.; Xie, B.G. A hydrophobin gene, Hyd9, plays an important role in the formation of aerial hyphae and primordia in Flammulina filiformis. Gene 2019, 706, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.D.; Ainouz, I.L.; Deoliveira, J.T.A.; Cavada, B.S. Plant-lectins, chemical and biological aspects. Mem. Do Inst. Oswaldo Cruz 1991, 86, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhao, C.G.; Tong, X.; Qi, Y.P. A lectin with mycelia differentiation and antiphytovirus activities from the edible mushroom Agrocybe aegerita. J. Biochem. Mol. Biol. 2003, 36, 214–222. [Google Scholar] [CrossRef]
- Nagata, Y.; Yamashita, M.; Honda, H.; Akabane, J.; Uehara, K.; Saito, A.; Sumisa, F.; Nishibori, K.; Oodaira, Y. Characterization, occurrence, and molecular cloning of a lectin from Grifola frondosa Jacalin-related lectin of fungal origin. Biosci. Biotechnol. Biochem. 2005, 69, 2374–2380. [Google Scholar] [CrossRef]
- Jung, E.C.; Kim, K.D.; Bae, C.H.; Kim, J.C.; Kim, D.K.; Kim, H.H. A mushroom lectin from ascomycete Cordyceps militaris. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2007, 1770, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Wang, H.; Ng, T.B. A haemagglutinin from the medicinal fungus Cordyceps militaris. Biosci. Rep. 2009, 29, 321–327. [Google Scholar] [CrossRef]
- Bao, D.P.; Ma, Y.W.; Gong, M.; Li, Y.; Gao, Y.N.; Yang, R.H.; Yang, R.F.; Mao, W.J.; Wang, Y. Sequence analysis and heterologous expression of lectin-like gene CMLec3 from the medicinal fungus Cordyceps militaris. Mycoscience 2018, 59, 119–123. [Google Scholar] [CrossRef]
- Wang, F.; Song, X.H.; Dong, X.M.; Zhang, J.J.; Dong, C.H. DASH-type cryptochromes regulate fruiting body development and secondary metabolism differently than CmWC-1 in the fungus Cordyceps militaris. Appl. Microbiol. Biotechnol. 2017, 101, 4645–4657. [Google Scholar] [CrossRef]
- Selby, C.P.; Sancar, A. A cryptochrome/photolyase class of enzymes with single-stranded DNA-specific photolyase activity. Proc. Natl. Acad. Sci. USA 2006, 103, 17696–17700. [Google Scholar] [CrossRef]
- Tagua, V.G.; Pausch, M.; Eckel, M.; Gutiérrez, G.; Miralles-Durán, A.; Sanz, C.; Eslava, A.P.; Pokorny, R.; Corrochano, L.M.; Batschauer, A. Fungal cryptochrome with DNA repair activity reveals an early stage in cryptochrome evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 15130–15135. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Fungal Light Sensing at the Bench and Beyond. In Advances in Genetics; Friedmann, T., Dunlap, J.C., Goodwin, S.F., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 96, pp. 1–51. [Google Scholar]
- Lou, H.W.; Zhao, Y.; Chen, B.X.; Yu, Y.H.; Tang, H.B.; Ye, Z.W.; Lin, J.F.; Guo, L.Q. Cmfhp Gene Mediates Fruiting Body Development and Carotenoid Production in Cordyceps militaris. Biomolecules 2020, 10, 410. [Google Scholar] [CrossRef]
- Wāng, Y.; Wang, R.; Wáng, Y.; Li, Y.; Yang, R.H.; Gong, M.; Shang, J.J.; Zhang, J.S.; Mao, W.J.; Zou, G.; et al. Diverse function and regulation of CmSnf1 in entomopathogenic fungus Cordyceps militaris. Fungal Genet. Biol. 2020, 142, 103415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Yin, Y.Y.; Cui, Y.; Zhang, Y.X.; Liu, B.Y.; Ma, Y.C.; Liu, Y.N.; Liu, G.Q. Chitinase Is Involved in the Fruiting Body Development of Medicinal Fungus Cordyceps militaris. Life 2023, 13, 764. [Google Scholar] [CrossRef]
- He, R.; Zhang, L.; Lan, J.; Mei, S.; Li, Y. Cmcrf1, a Putative Zn2Cys6 Fungal Transcription Factor, Is Involved in Conidiation, Carotenoid Production, and Fruiting Body Development in Cordyceps militaris. Biology 2022, 11, 1535. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, Y.Y.; Zhang, X.L.; Li, K.; Li, X.; Wang, F.; Xu, F.X.; Dong, C.H. Infection Process and Genome Assembly Provide Insights into the Pathogenic Mechanism of Destructive Mycoparasite Calcarisporium cordycipiticola with Host Specificity. J. Fungi 2021, 7, 918. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Kong, X.; Lu, Z.; Xiao, M.; Chen, M.; Zhu, L.; Shen, Y.; Hu, X.; Song, S. Para-aminobenzoic acid (PABA) synthase enhances thermotolerance of mushroom Agaricus bisporus. PLoS ONE 2014, 9, e91298. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, P.; Iotti, M.; Zeppa, S.D.; Lancellotti, E.; Amicucci, A.; Zambonelli, A. Morphological and functional changes in mycelium and mycorrhizas of Tuber borchii due to heat stress. Fungal Ecol. 2017, 29, 20–29. [Google Scholar] [CrossRef]
- Jiaojiao, Z.; Fen, W.; Kuanbo, L.; Qing, L.; Ying, Y.; Caihong, D. Heat and light stresses affect metabolite production in the fruit body of the medicinal mushroom Cordyceps militaris. Appl. Microbiol. Biotechnol. 2018, 102, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Sanmiya, K.; Suzuki, K.; Egawa, Y.; Shono, M. Mitochondrial small heat-shock protein enhances thermotolerance in tobacco plants. FEBS Lett. 2004, 557, 265–268. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, S.; Luo, Y.; Ma, C.; Gong, Y.; Zhou, Y.; Gao, S.; Huang, Z.; Yan, L.; Hu, Y.; et al. The heat shock protein 40 LeDnaJ regulates stress resistance and indole-3-acetic acid biosynthesis in Lentinula edodes. Fungal Genet. Biol. 2018, 118, 37–44. [Google Scholar] [CrossRef]
- Ling, Y.Y.; Ling, Z.L.; Zhao, R.L. Construction of a heat-resistant strain of Lentinus edodes by fungal Hsp20 protein overexpression and genetic transformation. Front. Microbiol. 2022, 13, 1009885. [Google Scholar] [CrossRef]
- Leong, Y.K.; Ma, T.W.; Chang, J.S.; Yang, F.C. Recent advances and future directions on the valorization of spent mushroom substrate (SMS): A review. Bioresour. Technol. 2022, 344, 126157. [Google Scholar] [CrossRef]
- Martín, C.; Zervakis, G.I.; Xiong, S.; Koutrotsios, G.; Strætkvern, K.O. Spent substrate from mushroom cultivation: Exploitation potential toward various applications and value-added products. Bioengineered 2023, 14, 2252138. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, M.; Chen, J.; Li, S.; Shao, Y.; Yang, J.; Li, J. Characteristics of bio-oil produced by the pyrolysis of mixed oil shale semi-coke and spent mushroom substrate. Fuel 2017, 200, 218–224. [Google Scholar] [CrossRef]
- Lam, S.S.; Lee, X.Y.; Nam, W.L.; Phang, X.Y.; Liew, R.K.; Yek, P.N.; Ho, Y.L.; Ma, N.L.; Rosli, M.H. Microwave vacuum pyrolysis conversion of waste mushroom substrate into biochar for use as growth medium in mushroom cultivation. J. Chem. Technol. Biotechnol. 2019, 94, 1406–1415. [Google Scholar] [CrossRef]
- Pérez-Chávez, A.M.; Mayer, L.; Albertó, E. Mushroom cultivation and biogas production: A sustainable reuse of organic resources. Energy Sustain. Dev. 2019, 50, 50–60. [Google Scholar] [CrossRef]
- Wan Mahari, W.A.; Peng, W.; Nam, W.L.; Yang, H.; Lee, X.Y.; Lee, Y.K.; Liew, R.K.; Ma, N.L.; Mohammad, A.; Sonne, C.; et al. A review on valorization of oyster mushroom and waste generated in the mushroom cultivation industry. J. Hazard. Mater. 2020, 400, 123156. [Google Scholar] [CrossRef] [PubMed]
- Jasiūnas, L.; Pedersen, T.H.; Toor, S.S.; Rosendahl, L.A. Biocrude production via supercritical hydrothermal co-liquefaction of spent mushroom compost and aspen wood sawdust. Renew. Energy 2017, 111, 392–398. [Google Scholar] [CrossRef]
- Ryden, P.; Efthymiou, M.N.; Tindyebwa, T.A.M.; Elliston, A.; Wilson, D.R.; Waldron, K.W.; Malakar, P.K. Bioethanol production from spent mushroom compost derived from chaff of millet and sorghum. Biotechnol. Biofuels 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Raethong, N.; Thananusak, R.; Cheawchanlertfa, P.; Prabhakaran, P.; Rattanaporn, K.; Laoteng, K.; Koffas, M.; Vongsangnak, W. Functional genomics and systems biology of Cordyceps species for biotechnological applications. Curr. Opin. Biotechnol. 2023, 81, 102939. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).