


Effects of Polyvinyl Chloride Microplastics on the Reproductive System, Intestinal Structure, and Microflora in Male and Female Mice
 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Grouping Treatment
2.2. Production of Mouse Cake
2.3. Paraffin Section and HE Staining of Mouse Reproductive System
2.4. HE Section Analysis of Reproductive System in Male and Female Mice
2.5. Collection of Intestinal Microbial Samples in Mice and Paraffin Sections and HE Staining of Intestinal Tissues
2.6. HE Section Analysis of Small Intestine in Male and Female Mice
 : Indicates the width of intestinal villi measured in male and female mice (the same picture identification can be found in the article by our research group by Su et al. [16]).
: Indicates the width of intestinal villi measured in male and female mice (the same picture identification can be found in the article by our research group by Su et al. [16]).2.7. Intestine Microbiome Analysis
2.8. Analysis of Small Intestinal Microbial Sequencing Data in Male and Female Mice
2.9. Statistical Analysis
3. Results
3.1. The Effect of Plastic PVC on the Structure of Reproductive System in Male and Female Mice
3.1.1. Effects of Microplastic PVC on the Morphology of Gonadal Organs in Male Mice
3.1.2. Effect of Microplastic PVC on the Morphology of Reproductive Gland Organs in Female Mice
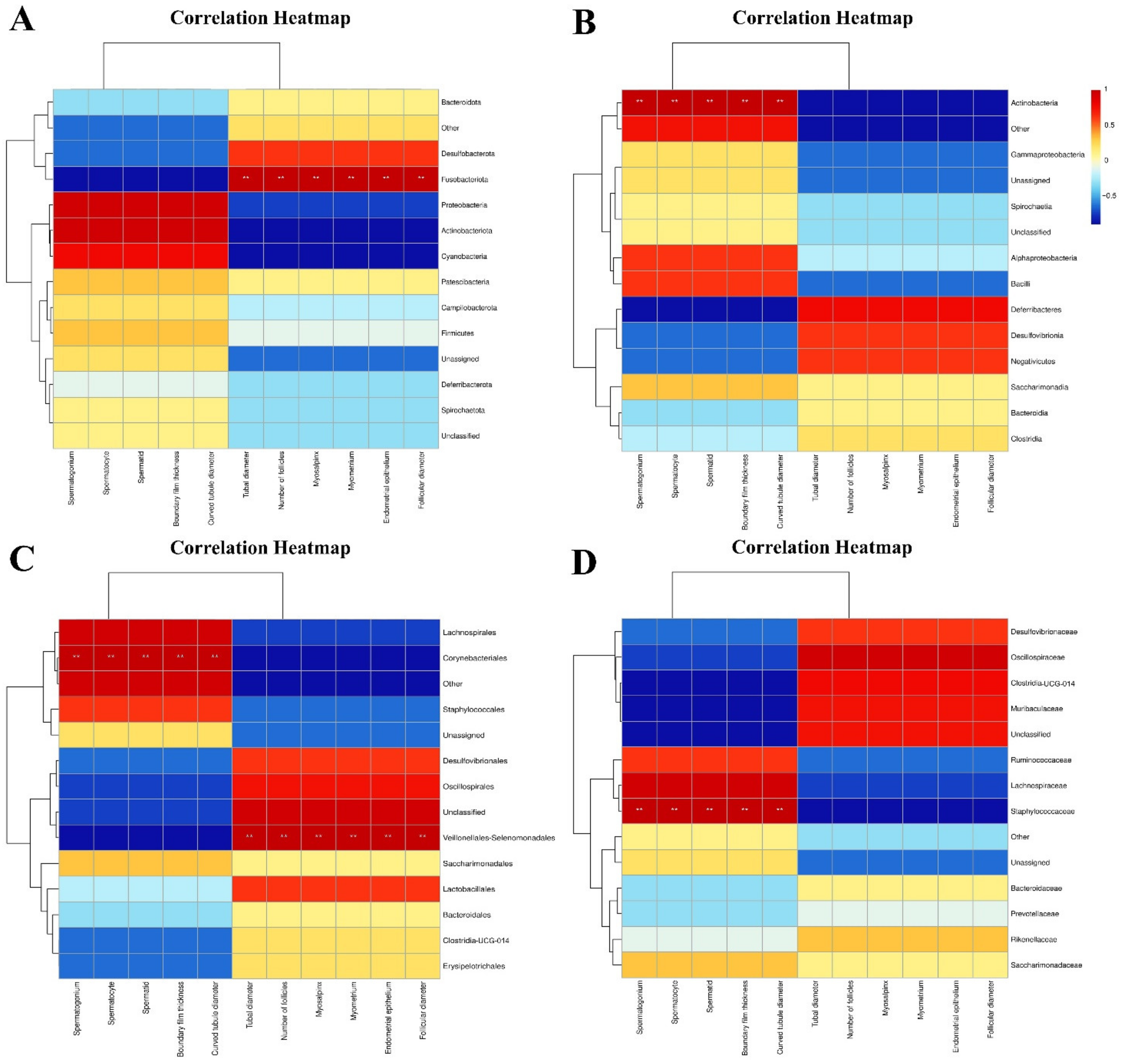
3.1.3. Association Analysis of Intestinal Microflora and Reproductive System Tissue and Organ Indexes in Male and Female Mice
3.2. Effects of Microplastic PVC on the Structure of Small Intestine in Male and Female Mice
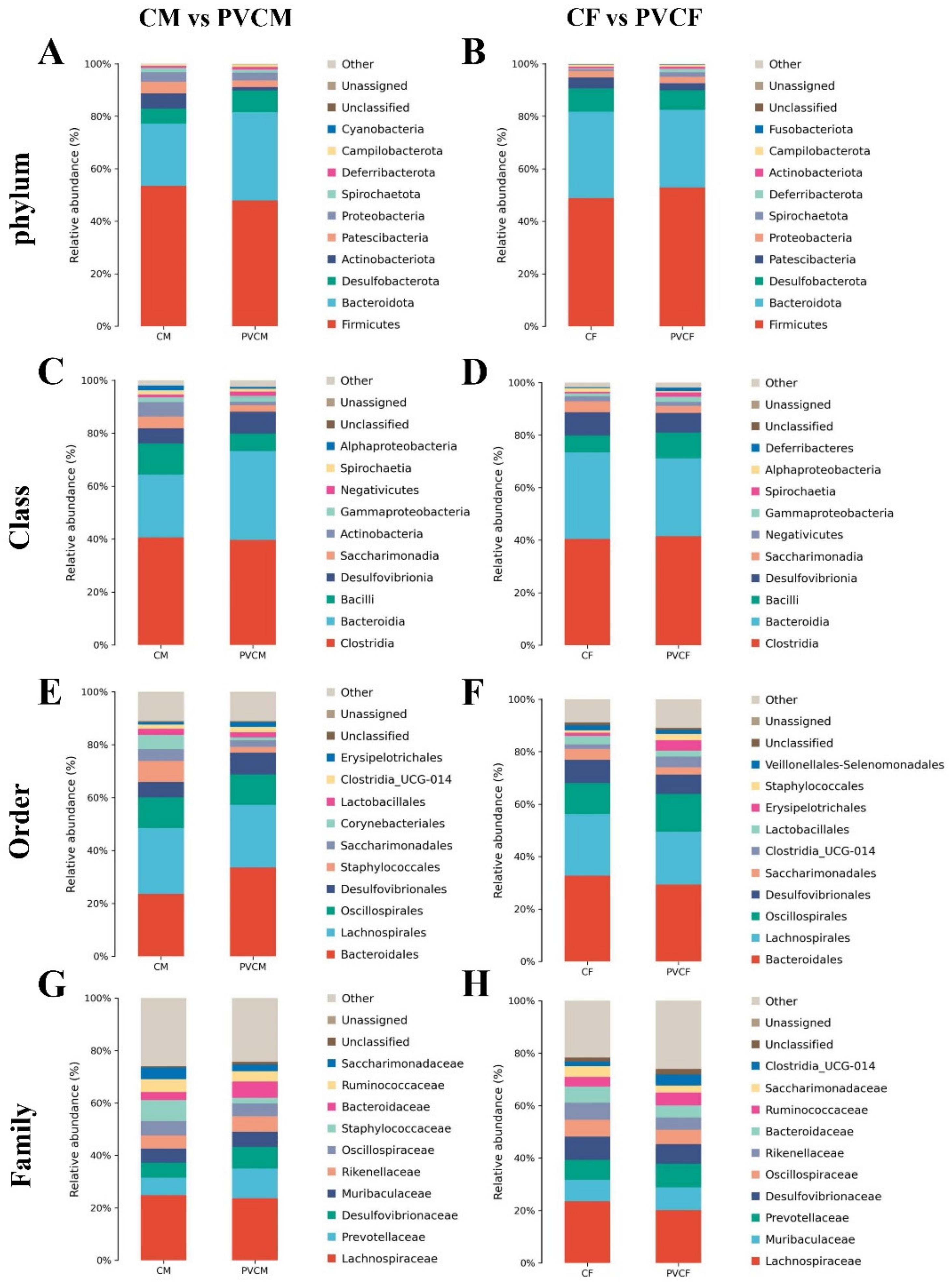
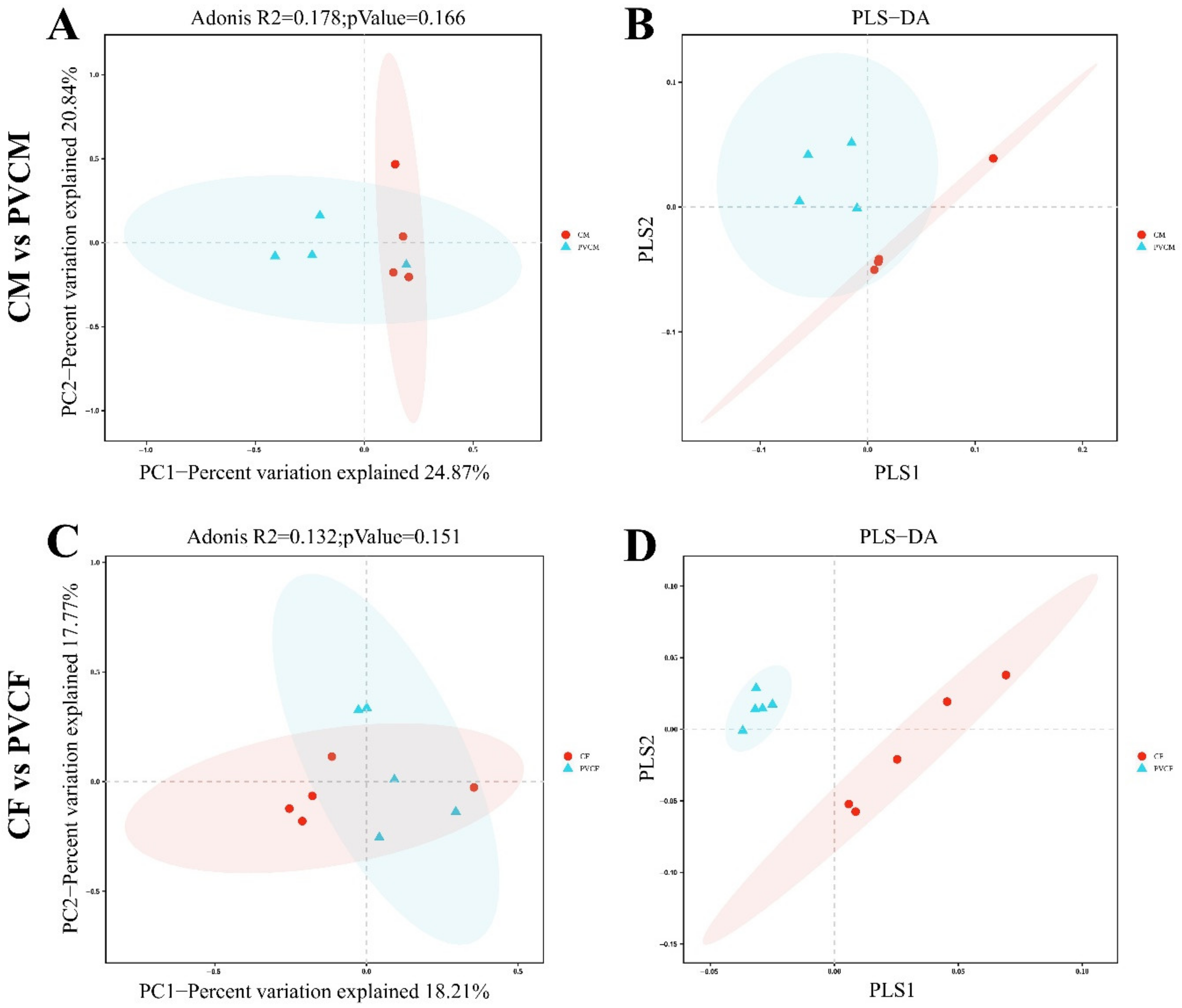
3.3. Taxonomic Analysis of Intestinal Microflora Characteristics of Male and Female Mice
3.4. Analysis of Species Composition of Intestinal Microflora in Male and Female Mice at Different Levels
3.5. Diversity Analysis of Intestinal Microbial Flora in Male and Female Mice
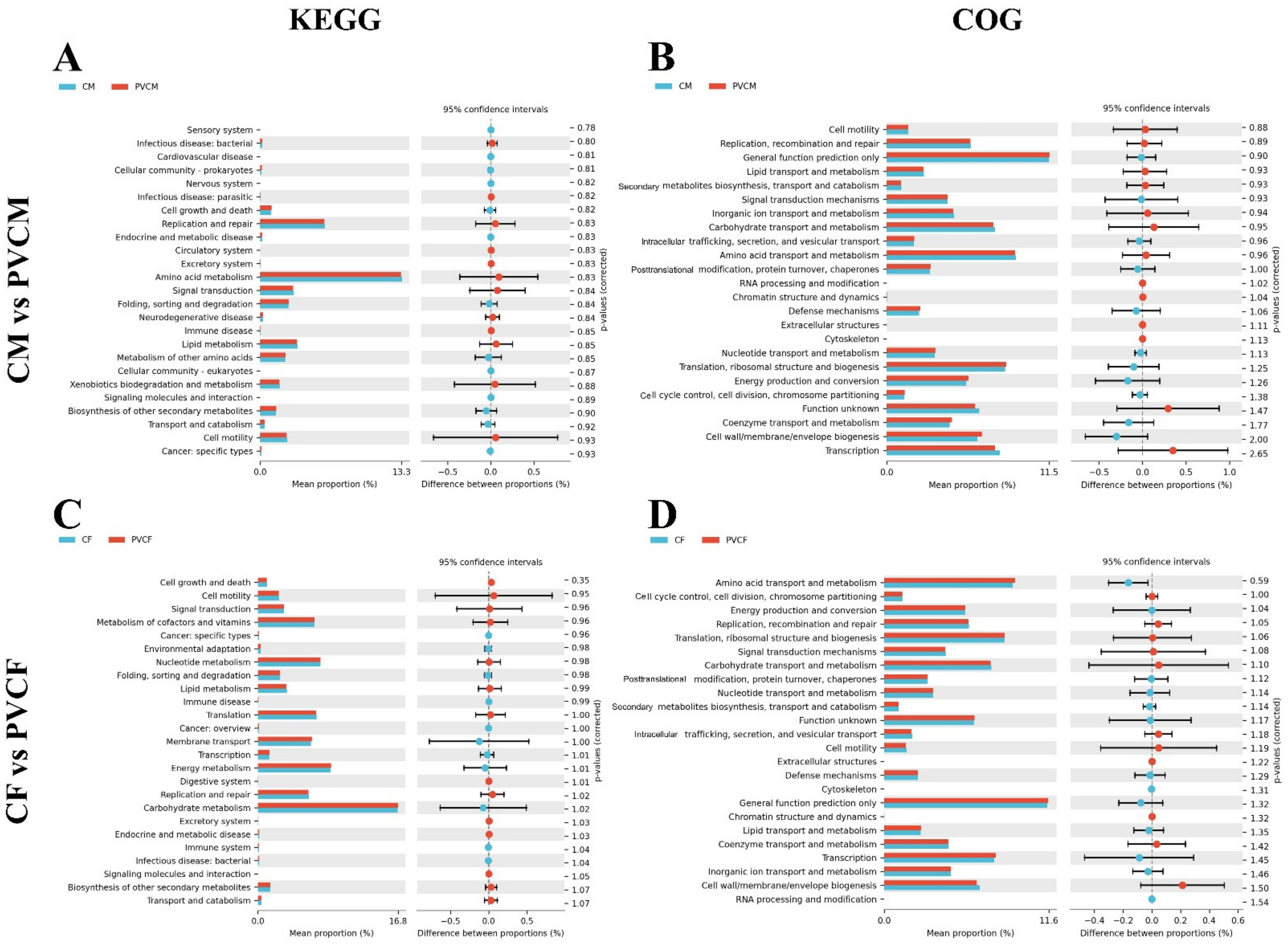
3.6. Functional Prediction Analysis of Intestinal Microflora in Male and Female Mice
4. Discussion
4.1. Effects of Microplastic PVC on the Reproductive System of Male Mice
4.2. Effects of Microplastic PVC on the Reproductive System of Female Mice
4.3. Correlation Analysis of Intestinal Microbiota and Reproductive System Tissue and Organ Indexes in Male and Female Mice
4.4. Effect and Analysis of Microplastic Polyvinyl Chloride on Small Intestine Microstructure of Male and Female Mice
4.5. Effects of Microplastic PVC on Intestinal Microflora in Male and Female Mice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Chen, L.; Shao, L.; Zhang, H.; Lu, F. Municipal solid waste (msw) landfill: A source of microplastics?—Evidence of microplastics in landfill leachate. Water Res. 2019, 159, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, N.P.; Fennell, T.R.; Johnson, L.M. Unintended human ingestion of nanoplastics and small microplastics through drinking water, beverages, and food sources. NanoImpact 2021, 21, 100302. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, R.C.; Cardoso, R.M.; Domingues, E.; Goncalves, R.V.; Lima, G.; Novaes, R.D. The emerging risk of microplastics and nanoplastics on the microstructure and function of reproductive organs in mammals: A systematic review of preclinical evidence. Life Sci. 2022, 295, 120404. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, G.; Wang, J.; Lu, L.; Li, X.; Zheng, Y.; Zhang, Z.; Ru, S. Microplastics increase the accumulation of phenanthrene in the ovaries of marine medaka (Oryzias melastigma) and its transgenerational toxicity. J. Hazard. Mater. 2022, 424, 127754. [Google Scholar] [CrossRef]
- Liu, Z.; Zhuan, Q.; Zhang, L.; Meng, L.; Fu, X.; Hou, Y. Polystyrene microplastics induced female reproductive toxicity in mice. J. Hazard. Mater. 2022, 424, 127629. [Google Scholar] [CrossRef]
- Hu, J.; Qin, X.; Zhang, J.; Zhu, Y.; Zeng, W.; Lin, Y.; Liu, X. Polystyrene microplastics disturb maternal-fetal immune balance and cause reproductive toxicity in pregnant mice. Reprod. Toxicol. 2021, 106, 42–50. [Google Scholar] [CrossRef]
- Prata, J.C.; Da, C.J.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef]
- Li, B.; Ding, Y.; Cheng, X.; Sheng, D.; Xu, Z.; Rong, Q.; Wu, Y.; Zhao, H.; Ji, X.; Zhang, Y. Polyethylene microplastics affect the distribution of gut microbiota and inflammation development in mice. Chemosphere 2020, 244, 125492. [Google Scholar] [CrossRef]
- Pedà, C.; Caccamo, L.; Fossi, M.C.; Gai, F.; Andaloro, F.; Genovese, L.; Perdichizzi, A.; Romeo, T.; Maricchiolo, G. Intestinal alterations in european sea bass Dicentrarchus labrax (Linnaeus, 1758) exposed to microplastics: Preliminary results. Environ. Pollut. 2016, 212, 251–256. [Google Scholar] [CrossRef]
- Sarker, A.; Deepo, D.M.; Nandi, R.; Rana, J.; Islam, S.; Rahman, S.; Hossain, M.N.; Islam, S.; Baroi, A.; Kim, J.-E. A review of microplastics pollution in the soil and terrestrial ecosystems: A global and bangladesh perspective. Sci. Total Environ. 2020, 733, 139296. [Google Scholar] [CrossRef] [PubMed]
- Spanogiannopoulos, P.; Bess, E.N.; Carmody, R.N.; Turnbaugh, P.J. The microbial pharmacists within us: A metagenomic view of xenobiotic metabolism. Nat. Rev. Microbiol. 2016, 14, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Lepage, P.; Leclerc, M.C.; Joossens, M.; Mondot, S.; Blottière, H.M.; Raes, J.; Ehrlich, D.; Doré, J. A metagenomic insight into our gut’s microbiome. Gut 2013, 62, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Botterell, Z.; Beaumont, N.; Dorrington, T.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability and effects of microplastics on marine zooplankton: A review. Environ. Pollut. 2019, 245, 98–110. [Google Scholar] [CrossRef]
- Ruff, W.E.; Greiling, T.M.; Kriegel, M.A. Host-microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 2020, 18, 521–538. [Google Scholar] [CrossRef]
- Su, Q.L.; Wu, J.; Tan, S.W.; Guo, X.Y.; Zou, D.Z.; Kang, K. The impact of microplastics polystyrene on the microscopic structure of mouse intestine, tight junction genes and gut microbiota. PLoS ONE 2024, 19, e304686. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhu, L.; Weng, J.; Jin, Z.; Cao, Y.; Jiang, H.; Zhang, Z. Detection and characterization of microplastics in the human testis and semen. Sci. Total Environ. 2023, 877, 162713. [Google Scholar] [CrossRef] [PubMed]
- Archana, D.; Supriya, C.; Girish, B.P.; Kishori, B.; Reddy, P.S. Alleviative effect of resveratrol on polyvinyl chloride-induced reproductive toxicity in male wistar rats. Food Chem. Toxicol. 2018, 116, 173–181. [Google Scholar] [CrossRef]
- Jin, H.; Yan, M.; Pan, C.; Liu, Z.; Sha, X.; Jiang, C.; Li, L.; Pan, M.; Li, D.; Han, X.; et al. Chronic exposure to polystyrene microplastics induced male reproductive toxicity and decreased testosterone levels via the lh-mediated lhr/camp/pka/star pathway. Part. Fibre Toxicol. 2022, 19, 13. [Google Scholar] [CrossRef]
- Jin, H.; Ma, T.; Sha, X.; Liu, Z.; Zhou, Y.; Meng, X.; Chen, Y.; Han, X.; Ding, J. Polystyrene microplastics induced male reproductive toxicity in mice. J. Hazard. Mater. 2021, 401, 123430. [Google Scholar] [CrossRef]
- Xie, X.; Deng, T.; Duan, J.; Xie, J.; Yuan, J.; Chen, M. Exposure to polystyrene microplastics causes reproductive toxicity through oxidative stress and activation of the p38 mapk signaling pathway. Ecotoxicol. Environ. Saf. 2020, 190, 110133. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.C.; Kuo, Y.T.; Leon Guo, Y.; Chen, J.R.; Tsai, S.S.; Chao, H.R.; Teng, Y.N.; Pan, M.H. The adverse effects of low-dose exposure to di(2-ethylhexyl) phthalate during adolescence on sperm function in adult rats. Environ. Toxicol. 2016, 31, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Q.; Yu, H.; Yang, L.; Sun, Y.; Xu, N.; Wang, N.; Lei, Z.; Hou, J.; Jin, Y.; et al. Polystyrene microplastics induce blood-testis barrier disruption regulated by the mapk-nrf2 signaling pathway in rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 47921–47931. [Google Scholar] [CrossRef]
- Hou, B.; Wang, F.; Liu, T.; Wang, Z. Reproductive toxicity of polystyrene microplastics: In vivo experimental study on testicular toxicity in mice. J. Hazard. Mater. 2021, 405, 124028. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Lei, Z.; Cui, L.; Hou, Y.; Yang, L.; An, R.; Wang, Q.; Li, S.; Zhang, H.; Zhang, L. Polystyrene microplastics lead to pyroptosis and apoptosis of ovarian granulosa cells via nlrp3/caspase-1 signaling pathway in rats. Ecotoxicol. Environ. Saf. 2021, 212, 112012. [Google Scholar] [CrossRef] [PubMed]
- Da, B.M.; de Albuquerque, F.O.; de Andrade, A.Z.; Cardoso, R.L.; Jordao, J.A.; Navarro, P.A. Increased concentration of 8-hydroxy-2’-deoxyguanosine in follicular fluid of infertile women with endometriosis. Cell Tissue Res. 2016, 366, 231–242. [Google Scholar]
- Wei, Z.; Wang, Y.; Wang, S.; Xie, J.; Han, Q.; Chen, M. Comparing the effects of polystyrene microplastics exposure on reproduction and fertility in male and female mice. Toxicology 2022, 465, 153059. [Google Scholar] [CrossRef]
- Ali, W.; Chen, Y.; Shah, M.G.; Buriro, R.S.; Sun, J.; Liu, Z.; Zou, H. Ferroptosis: First evidence in premature duck ovary induced by polyvinyl chloride microplastics. Sci. Total Environ. 2024, 933, 173032. [Google Scholar] [CrossRef]
- Haddadi, A.; Kessabi, K.; Boughammoura, S.; Ben Rhouma, M.; Mlouka, R.; Banni, M.; Messaoudi, I. Exposure to microplastics leads to a defective ovarian function and change in cytoskeleton protein expression in rat. Environ. Sci. Pollut. Res. Int. 2022, 29, 34594–34606. [Google Scholar] [CrossRef]
- Harris, E.A.; Stephens, K.K.; Winuthayanon, W. Extracellular vesicles and the oviduct function. Int. J. Mol. Sci. 2020, 21, 8280. [Google Scholar] [CrossRef]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1–R26. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Cao, M.; Peng, T.; Shan, H.; Lian, W.; Yu, Y.; Shui, G.; Li, R. Features, potential invasion pathways, and reproductive health risks of microplastics detected in human uterus. Environ. Sci. Technol. 2024, 58, 10482–10493. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xu, T.; Chen, T.; Liu, J.; Xu, S. Oxidative stress mediated by the tlr4/nox2 signalling axis is involved in polystyrene microplastic-induced uterine fibrosis in mice. Sci. Total Environ. 2022, 838, 155825. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Qin, Y.; Wang, M.; Xu, W.; Chen, Y.; Zheng, L.; Chen, W.; Luo, T. Microplastics from agricultural plastic mulch films: A mini-review of their impacts on the animal reproductive system. Ecotoxicol. Environ. Saf. 2022, 244, 114030. [Google Scholar] [CrossRef]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef]
- Brunkwall, L.; Orho-Melander, M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: From current human evidence to future possibilities. Diabetologia 2017, 60, 943–951. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Baj, A.; Moro, E.; Bistoletti, M.; Orlandi, V.; Crema, F.; Giaroni, C. Glutamatergic signaling along the microbiota-gut-brain axis. Int. J. Mol. Sci. 2019, 20, 1482. [Google Scholar] [CrossRef]
- Agus, A.; Clement, K.; Sokol, H. Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut 2021, 70, 1174–1182. [Google Scholar] [CrossRef]
- Pellock, S.J.; Redinbo, M.R. Glucuronides in the gut: Sugar-driven symbioses between microbe and host. J. Biol. Chem. 2017, 292, 8569–8576. [Google Scholar] [CrossRef]
- Ding, N.; Zhang, X.; Di Zhang, X.; Jing, J.; Liu, S.S.; Mu, Y.P.; Peng, L.L.; Yan, Y.J.; Xiao, G.M.; Bi, X.Y.; et al. Impairment of spermatogenesis and sperm motility by the high-fat diet-induced dysbiosis of gut microbes. Gut 2020, 69, 1608–1619. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Zhao, Y.; Liu, S.; Yuan, H.; You, T.; Xu, H. Microplastics-perturbed gut microbiota triggered the testicular disorder in male mice: Via fecal microbiota transplantation. Environ. Pollut. 2022, 309, 119789. [Google Scholar] [CrossRef] [PubMed]
- Belizario, J.E.; Napolitano, M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef] [PubMed]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, C.; Scully, P.; O’Mahony, D.; Murphy, S.; O’Brien, F.; Lyons, A.; Sherlock, G.; MacSharry, J.; Kiely, B.; Shanahan, F.; et al. Commensal-induced regulatory t cells mediate protection against pathogen-stimulated nf-kappab activation. PLoS Pathog. 2008, 4, e1000112. [Google Scholar] [CrossRef] [PubMed]
- Lyons, A.; O’Mahony, D.; O’Brien, F.; MacSharry, J.; Sheil, B.; Ceddia, M.; Russell, W.M.; Forsythe, P.; Bienenstock, J.; Kiely, B.; et al. Bacterial strain-specific induction of foxp3+ t regulatory cells is protective in murine allergy models. Clin. Exp. Allergy 2010, 40, 811–819. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P.; Izuka, E.; Menuba, I.; Jegasothy, R.; Nwagha, U. Staphylococcal infections and infertility: Mechanisms and management. Mol. Cell. Biochem. 2020, 474, 57–72. [Google Scholar] [CrossRef]
- Zhang, J.; Meng, H.; Kong, X.; Cheng, X.; Ma, T.; He, H.; Du, W.; Yang, S.; Li, S.; Zhang, L. Combined effects of polyethylene and organic contaminant on zebrafish (Danio rerio): Accumulation of 9-nitroanthracene, biomarkers and intestinal microbiota. Environ. Pollut. 2021, 277, 116767. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Sharifinia, M.; Bahmanbeigloo, Z.A.; Keshavarzifard, M.; Khanjani, M.H.; Lyons, B.P. Microplastic pollution as a grand challenge in marine research: A closer look at their adverse impacts on the immune and reproductive systems. Ecotoxicol. Environ. Saf. 2020, 204, 111109. [Google Scholar] [CrossRef]
- Subramaniam, M.D.; Kim, I.H. Clays as dietary supplements for swine: A review. J. Anim. Sci. Biotechnol. 2015, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Bonis, V.; Rossell, C.; Gehart, H. The intestinal epithelium—Fluid fate and rigid structure from crypt bottom to villus tip. Front. Cell Dev. Biol. 2021, 9, 661931. [Google Scholar] [CrossRef] [PubMed]
- Ahrendt, C.; Perez-Venegas, D.; Urbina, M.; Gonzalez, C.; Echeveste, P.; Aldana, M.; Pulgar, J.; Galbán-Malagón, C. Microplastic ingestion cause intestinal lesions in the intertidal fish girella laevifrons. Mar. Pollut. Bull. 2020, 151, 110795. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, S.; Nie, Q.; He, H.; Tan, H.; Geng, F.; Ji, H.; Hu, J.; Nie, S. Gut firmicutes: Relationship with dietary fiber and role in host homeostasis. Crit. Rev. Food Sci. Nutr. 2023, 63, 12073–12088. [Google Scholar] [CrossRef]
- Kaur, A.; Chen, T.; Green, S.J.; Mutlu, E.; Martin, B.R.; Rumpagaporn, P.; Patterson, J.A.; Keshavarzian, A.; Hamaker, B.R. Physical inaccessibility of a resistant starch shifts mouse gut microbiota to butyrogenic firmicutes. Mol. Nutr. Food Res. 2019, 63, e1801012. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lu, L.; Tu, W.; Luo, T.; Fu, Z. Impacts of polystyrene microplastic on the gut barrier, microbiota and metabolism of mice. Sci. Total Environ. 2019, 649, 308–317. [Google Scholar] [CrossRef]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding type and development drive the ingestion of microplastics by freshwater invertebrates. Sci. Rep. 2017, 7, 17006. [Google Scholar] [CrossRef]
- Zhao, H.B.; Jia, L.; Yan, Q.Q.; Deng, Q.; Wei, B. Effect of clostridium butyricum and butyrate on intestinal barrier functions: Study of a rat model of severe acute pancreatitis with intra-abdominal hypertension. Front. Physiol. 2020, 11, 561061. [Google Scholar] [CrossRef]
- Evans, N.J.; Brown, J.M.; Murray, R.D.; Getty, B.; Birtles, R.J.; Hart, C.A.; Carter, S.D. Characterization of novel bovine gastrointestinal tract treponema isolates and comparison with bovine digital dermatitis treponemes. Appl. Environ. Microbiol. 2011, 77, 138–147. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M.J. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef]
- Gibiino, G.; Lopetuso, L.R.; Scaldaferri, F.; Rizzatti, G.; Binda, C.; Gasbarrini, A. Exploring bacteroidetes: Metabolic key points and immunological tricks of our gut commensals. Dig. Liver Dis. 2018, 50, 635–639. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Cheesman, S.E.; Neal, J.T.; Mittge, E.; Seredick, B.M.; Guillemin, K. Epithelial cell proliferation in the developing zebrafish intestine is regulated by the wnt pathway and microbial signaling via myd88. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4570–4577. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar, R.D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Strukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Sun, H.; Chen, N.; Yang, X.; Xia, Y.; Wu, D. Effects induced by polyethylene microplastics oral exposure on colon mucin release, inflammation, gut microflora composition and metabolism in mice. Ecotoxicol. Environ. Saf. 2021, 220, 112340. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Chen, R.; Wang, M.; Bai, R.; Cui, X.; Liu, Y.; Wu, C.; Chen, C. Perturbation of gut microbiota plays an important role in micro/nanoplastics-induced gut barrier dysfunction. Nanoscale 2021, 13, 8806–8816. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, X.; Wang, Z.; He, F.; Huang, Z.; Chen, C.; Tang, B.; Qin, M.; Wu, Y.; Long, C.; et al. Analysis of differences in intestinal flora associated with different bmi status in colorectal cancer patients. J. Transl. Med. 2024, 22, 142. [Google Scholar] [CrossRef]
- Martin, R.; Nauta, A.J.; Ben, A.K.; Knippels, L.M.; Knol, J.; Garssen, J. Early life: Gut microbiota and immune development in infancy. Benef. Microbes 2010, 1, 367–382. [Google Scholar] [CrossRef]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene microplastics induce microbiota dysbiosis and inflammation in the gut of adult zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef]
- El Aidy, S.; Derrien, M.; Merrifield, C.A.; Levenez, F.; Doré, J.; Boekschoten, M.V.; Dekker, J.; Holmes, E.; Zoetendal, E.G.; van Baarlen, P.; et al. Gut bacteria-host metabolic interplay during conventionalisation of the mouse germfree colon. ISME J. 2013, 7, 743–755. [Google Scholar] [CrossRef] [PubMed]
 : Indicates the diameter of the tubule of the testes measured in the CM and PVCM groups. ↑: Indicates the location of spermatogonia. ★: Indicates the position of the spermatocytes. ●: Indicates the location of spermatozoa. Data are presented as mean ± SD. **, p < 0.01. Scale bar = 100 μm.
: Indicates the diameter of the tubule of the testes measured in the CM and PVCM groups. ↑: Indicates the location of spermatogonia. ★: Indicates the position of the spermatocytes. ●: Indicates the location of spermatozoa. Data are presented as mean ± SD. **, p < 0.01. Scale bar = 100 μm.
: Indicates the diameter of the tubule of the testes measured in the CM and PVCM groups. ↑: Indicates the location of spermatogonia. ★: Indicates the position of the spermatocytes. ●: Indicates the location of spermatozoa. Data are presented as mean ± SD. **, p < 0.01. Scale bar = 100 μm.
: Indicates the diameter of the tubule of the testes measured in the CM and PVCM groups. ↑: Indicates the location of spermatogonia. ★: Indicates the position of the spermatocytes. ●: Indicates the location of spermatozoa. Data are presented as mean ± SD. **, p < 0.01. Scale bar = 100 μm.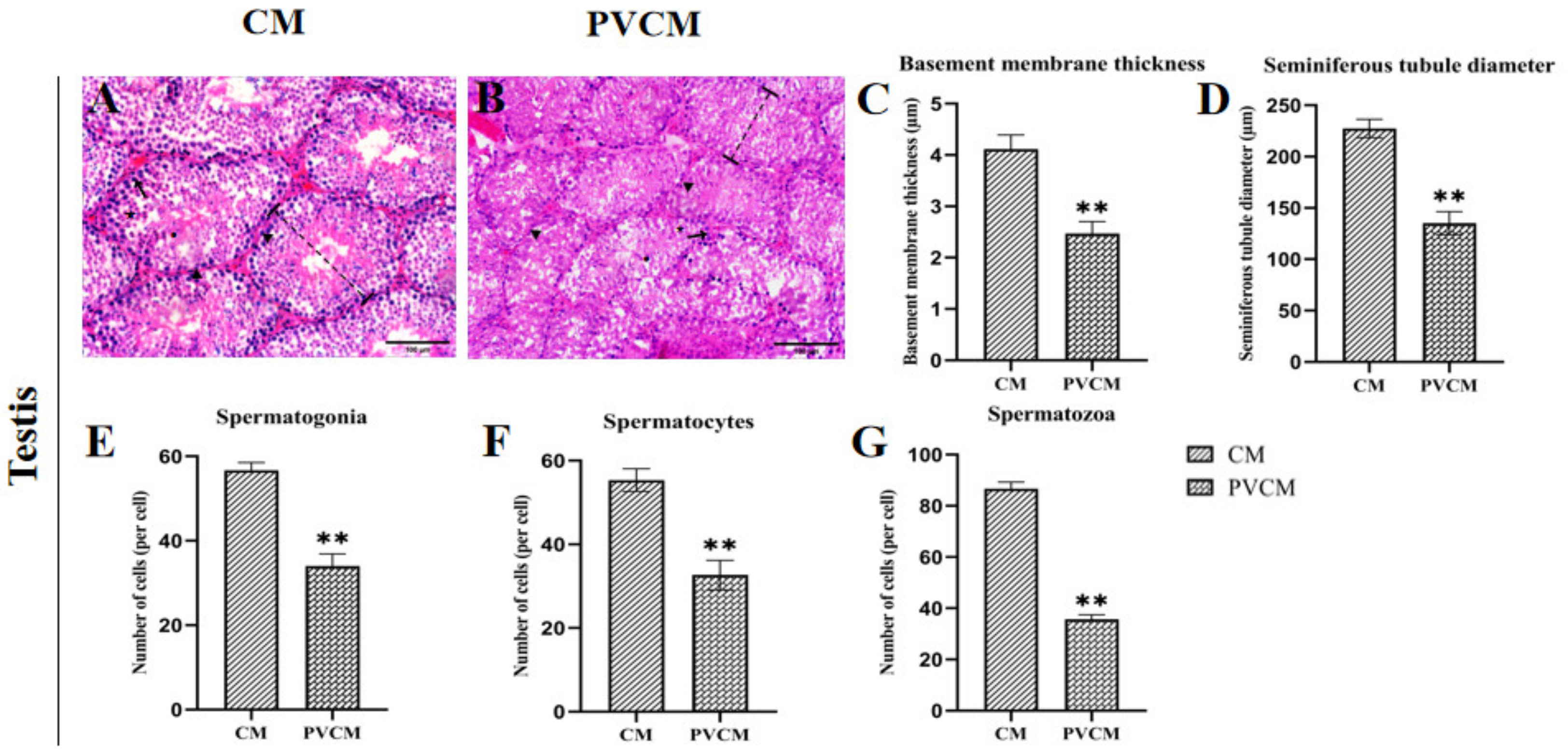
 : Indicates the follicle diameter of the ovary measured of CF and PVCF. ↑: Indicates the follicle. (C) Uterus group of CF; (D) uterine group of PVCF; (The same magnification figure (B–D) shows that the uterus is significantly reduced under PVC exposure) (E) uterine group of CF; (F) uterine group of PVCF; ▲: Indicates the position of the uterine epithelium.
: Indicates the follicle diameter of the ovary measured of CF and PVCF. ↑: Indicates the follicle. (C) Uterus group of CF; (D) uterine group of PVCF; (The same magnification figure (B–D) shows that the uterus is significantly reduced under PVC exposure) (E) uterine group of CF; (F) uterine group of PVCF; ▲: Indicates the position of the uterine epithelium.  : Indicates the thickness of the muscle layer of the uterus measured. (G) Fallopian tube group of CF; (H) Fallopian tube group of PVCF; ★: Indicates the position of fallopian tube epithelium.
: Indicates the thickness of the muscle layer of the uterus measured. (G) Fallopian tube group of CF; (H) Fallopian tube group of PVCF; ★: Indicates the position of fallopian tube epithelium.  : Indicates the muscular thickness of the fallopian tube measured. (a) Follicle diameter of CF and PVCF; (b) number of follicles of CF and PVCF; (c) myometrium thickness of CF and PVCF; (d) thickness of uterine epithelial cells of CF and PVCF; (e) Thickness of musculature of tubal tube; (f) diameter of tubal lumen; Data are presented as mean ± SD. *, p < 0.05; **, p < 0.01. Scale bar = 400 μm in (A,B); 200 μm in (C,D); 100 μm in (E–H).
: Indicates the follicle diameter of the ovary measured of CF and PVCF. ↑: Indicates the follicle. (C) Uterus group of CF; (D) uterine group of PVCF; (The same magnification figure (B–D) shows that the uterus is significantly reduced under PVC exposure) (E) uterine group of CF; (F) uterine group of PVCF; ▲: Indicates the position of the uterine epithelium. : Indicates the thickness of the muscle layer of the uterus measured. (G) Fallopian tube group of CF; (H) Fallopian tube group of PVCF; ★: Indicates the position of fallopian tube epithelium. : Indicates the muscular thickness of the fallopian tube measured. (a) Follicle diameter of CF and PVCF; (b) number of follicles of CF and PVCF; (c) myometrium thickness of CF and PVCF; (d) thickness of uterine epithelial cells of CF and PVCF; (e) Thickness of musculature of tubal tube; (f) diameter of tubal lumen; Data are presented as mean ± SD. *, p < 0.05; **, p < 0.01. Scale bar = 400 μm in (A,B); 200 μm in (C,D); 100 μm in (E–H).
: Indicates the muscular thickness of the fallopian tube measured. (a) Follicle diameter of CF and PVCF; (b) number of follicles of CF and PVCF; (c) myometrium thickness of CF and PVCF; (d) thickness of uterine epithelial cells of CF and PVCF; (e) Thickness of musculature of tubal tube; (f) diameter of tubal lumen; Data are presented as mean ± SD. *, p < 0.05; **, p < 0.01. Scale bar = 400 μm in (A,B); 200 μm in (C,D); 100 μm in (E–H).
: Indicates the follicle diameter of the ovary measured of CF and PVCF. ↑: Indicates the follicle. (C) Uterus group of CF; (D) uterine group of PVCF; (The same magnification figure (B–D) shows that the uterus is significantly reduced under PVC exposure) (E) uterine group of CF; (F) uterine group of PVCF; ▲: Indicates the position of the uterine epithelium. : Indicates the thickness of the muscle layer of the uterus measured. (G) Fallopian tube group of CF; (H) Fallopian tube group of PVCF; ★: Indicates the position of fallopian tube epithelium. : Indicates the muscular thickness of the fallopian tube measured. (a) Follicle diameter of CF and PVCF; (b) number of follicles of CF and PVCF; (c) myometrium thickness of CF and PVCF; (d) thickness of uterine epithelial cells of CF and PVCF; (e) Thickness of musculature of tubal tube; (f) diameter of tubal lumen; Data are presented as mean ± SD. *, p < 0.05; **, p < 0.01. Scale bar = 400 μm in (A,B); 200 μm in (C,D); 100 μm in (E–H).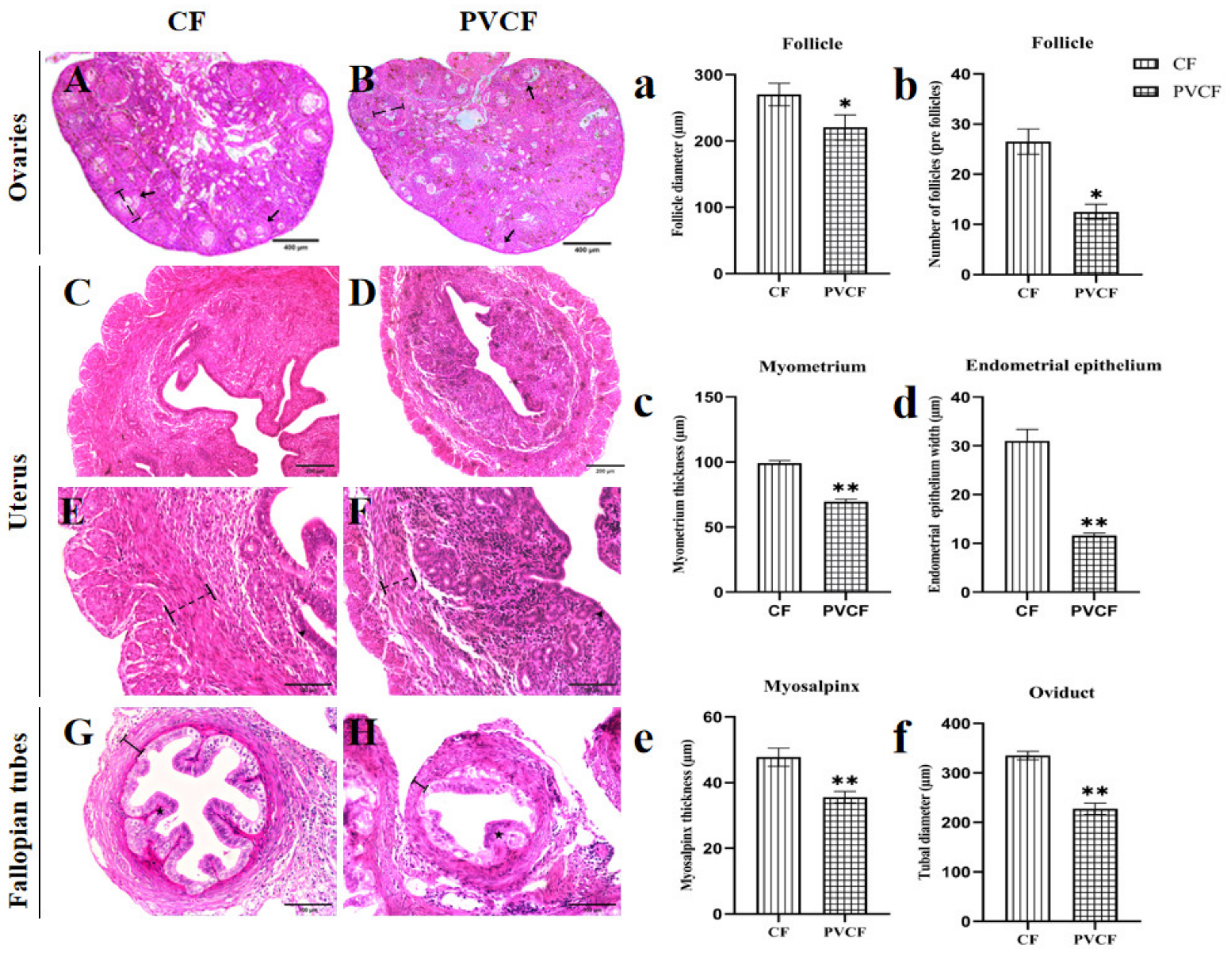
 : Indicates the width of intestinal villi measured in male and female mice. Scale bar = 100 μm.
: Indicates the width of intestinal villi measured in male and female mice. Scale bar = 100 μm.
: Indicates the width of intestinal villi measured in male and female mice. Scale bar = 100 μm.
: Indicates the width of intestinal villi measured in male and female mice. Scale bar = 100 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Object | CF | CM | PVCF | PVCM |
|---|---|---|---|---|---|
| Duodenum | Villus wide/µm | 57.66 ± 2.23 | 58.22 ± 4.15 | 66.06 ± 5.75 | 50.82 ± 4.00 |
| Villus height/µm | 814.97 ± 29.67 a | 696.08 ± 20.38 b | 648.08 ± 28.06 c | 575.75 ± 27.58 d | |
| Crypt depth/µm | 117.42 ± 4.21 ab | 101.18 ± 3.88 b | 159.91 ± 4.97 a | 128.62 ± 4.37 a | |
| Villus height/Crypt depth | 6.95 ± 0.48 | 6.89 ± 0.27 | 4.05 ± 0.12 | 4.48 ± 0.15 | |
| Jejunum | Villus wide/µm | 69.14 ± 8.77 a | 60.93 ± 4.15 a | 58.39 ± 9.72 b | 51.46 ± 4.00 b |
| Villus height/µm | 745.91 ± 34.68 b | 796.88 ± 12.42 a | 614.32 ± 28.55 d | 708.87 ± 18.48 c | |
| Crypt depth/µm | 110.12 ± 4.77 b | 104.44 ± 3.66 b | 140.81 ± 8.54 ab | 163.88 ± 6.49 a | |
| Villus height/Crypt depth | 6.79 ± 0.53 | 7.64 ± 0.35 | 4.38 ± 0.35 | 4.33 ± 0.14 | |
| Ileum | Villus wide/µm | 62.55 ± 4.71 a | 55.12 ± 4.15 ab | 46.71 ± 4.54 b | 41.17 ± 4.00 b |
| Villus height/µm | 589.58 ± 23.23 b | 715.89 ± 34.63 a | 482.72 ± 19.27 c | 575.66 ± 28.44 b | |
| Crypt depth/µm | 112.54 ± 5.23 b | 102.64 ± 4.00 b | 132.46 ± 4.40 ab | 149.61 ± 5.71 a | |
| Villus height/Crypt depth | 5.25 ± 0.40 a | 6.99 ± 0.52 a | 3.65 ± 0.23 b | 3.85 ± 0.24 b |
| Sample | Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|---|
| CF | 12.2 ± 0.45 A | 17.6 ± 0.55 a | 36.8 ± 2.17 | 55 ± 3.94 | 106.2 ± 6.10 | 32.6 ± 4.93 |
| PVCF | 11.8 ± 0.45 ab | 17.2 ± 0.84 a | 37.2 ± 2.59 | 56 ± 4.30 | 106.4 ± 9.40 | 31.2 ± 3.83 |
| CM | 10.75 ± 0.96 B | 16.25 ± 0.50 b | 35 ± 0.82 | 55 ± 3.56 | 107 ± 1.63 | 30.25 ± 2.06 |
| PVCM | 11.5 ± 0.58 b | 17 ± 1.41 ab | 36 ± 1.63 | 53.75 ± 6.08 | 105.75 ± 8.46 | 34.5 ± 4.43 |
| Sample | Feature | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|
| CF | 802.60 ± 89.16 a | 810.08 ± 91.55 a | 810.98 ± 92.02 a | 7.80 ± 0.16 | 0.99 ± 0.01 |
| CM | 750.75 ± 104.10 ab | 759.55 ± 103.86 ab | 759.21 ± 103.46 ab | 7.54 ± 0.68 | 0.98 ± 0.02 |
| PVCF | 753.40 ± 106.89 ab | 757.50 ± 109.30 ab | 757.75 ± 110.24 ab | 7.76 ± 0.33 | 0.99 ± 0.00 |
| PVCM | 686.25 ± 84.68 b | 688.47 ± 85.79 b | 687.64 ± 85.42 b | 7.49 ± 0.44 | 0.980.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.-K.-X.; Ge, S.-J.; Su, Q.-L.; Chen, J.-J.; Wu, J.; Kang, K. Effects of Polyvinyl Chloride Microplastics on the Reproductive System, Intestinal Structure, and Microflora in Male and Female Mice. Vet. Sci. 2024, 11, 488. https://doi.org/10.3390/vetsci11100488
Yang Y-K-X, Ge S-J, Su Q-L, Chen J-J, Wu J, Kang K. Effects of Polyvinyl Chloride Microplastics on the Reproductive System, Intestinal Structure, and Microflora in Male and Female Mice. Veterinary Sciences. 2024; 11(10):488. https://doi.org/10.3390/vetsci11100488
Chicago/Turabian StyleYang, Yang-Kai-Xin, Shu-Jun Ge, Qi-Ling Su, Jin-Jun Chen, Jiang Wu, and Kai Kang. 2024. "Effects of Polyvinyl Chloride Microplastics on the Reproductive System, Intestinal Structure, and Microflora in Male and Female Mice" Veterinary Sciences 11, no. 10: 488. https://doi.org/10.3390/vetsci11100488
APA StyleYang, Y.-K.-X., Ge, S.-J., Su, Q.-L., Chen, J.-J., Wu, J., & Kang, K. (2024). Effects of Polyvinyl Chloride Microplastics on the Reproductive System, Intestinal Structure, and Microflora in Male and Female Mice. Veterinary Sciences, 11(10), 488. https://doi.org/10.3390/vetsci11100488