


Impact of Equine Ocular Surface Squamous Neoplasia on Interactions between Ocular Transcriptome and Microbiome

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Horses
2.2. Sample Collection and Processing
2.3. RNA Sequencing
2.4. Microbial 16S Sequencing
2.5. Interactome Analysis of Transcriptome and Microbiome Data
3. Results
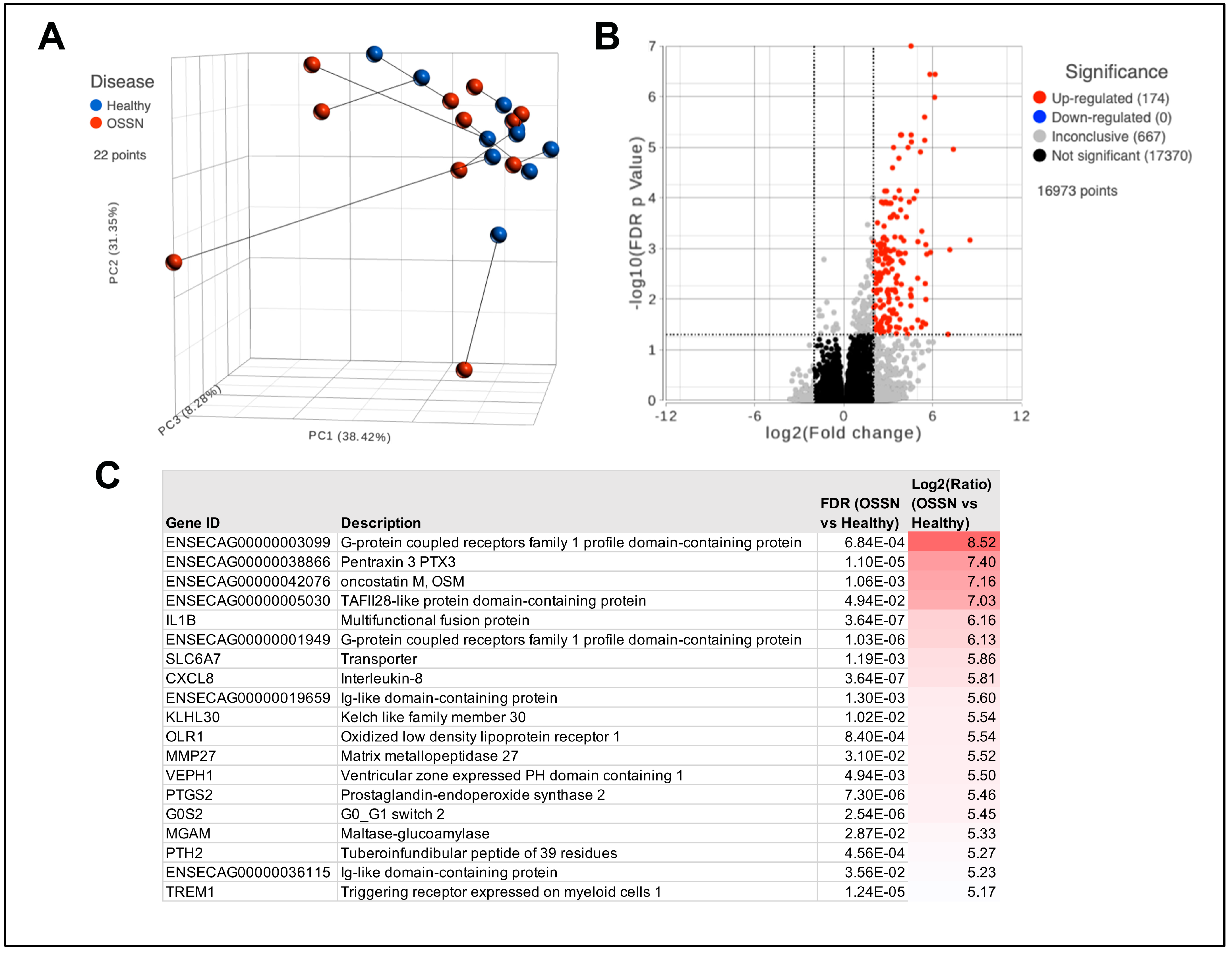
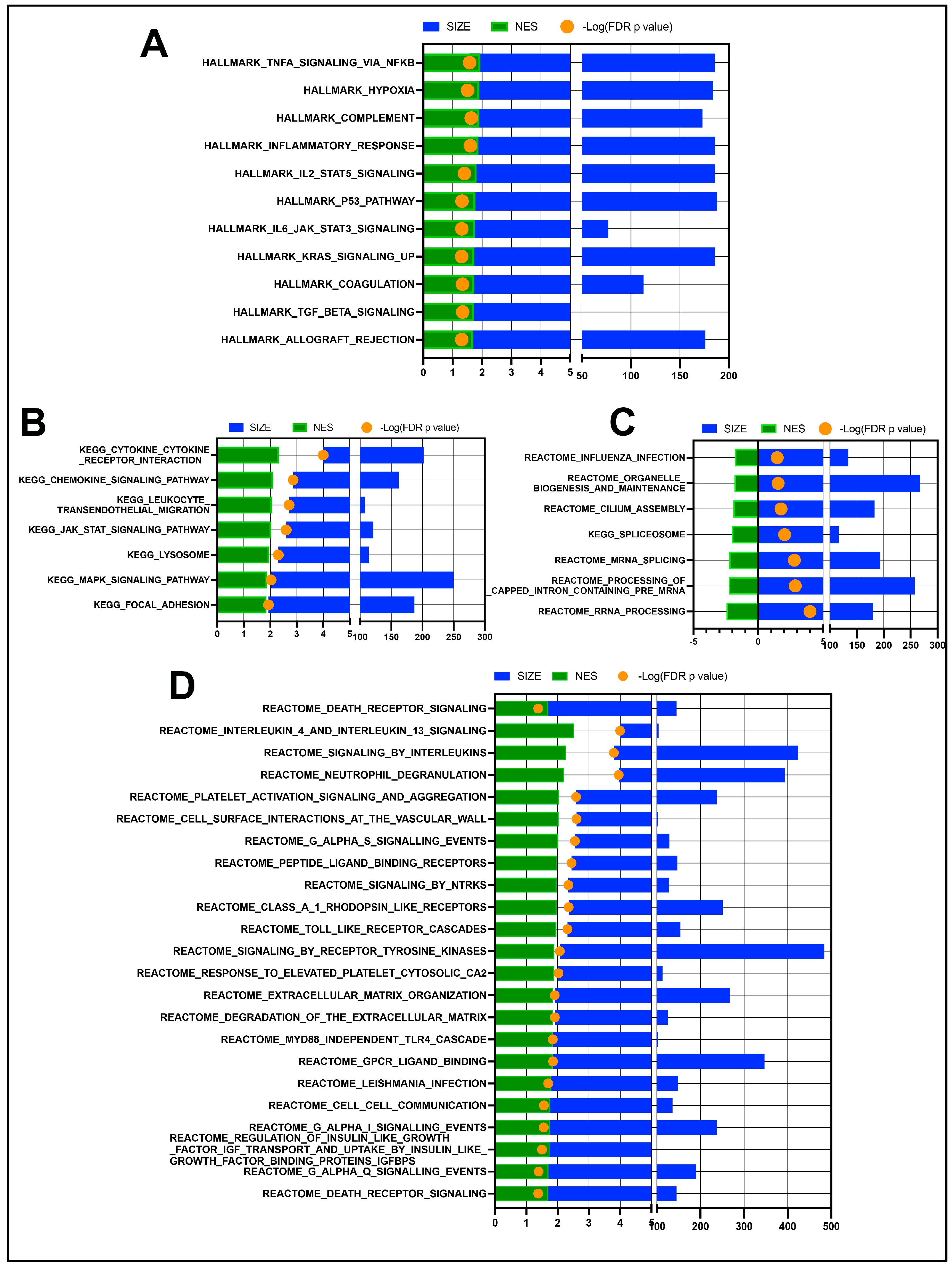
3.1. Transcriptomic Differences in OSSN Effected Surface Ocular Cells
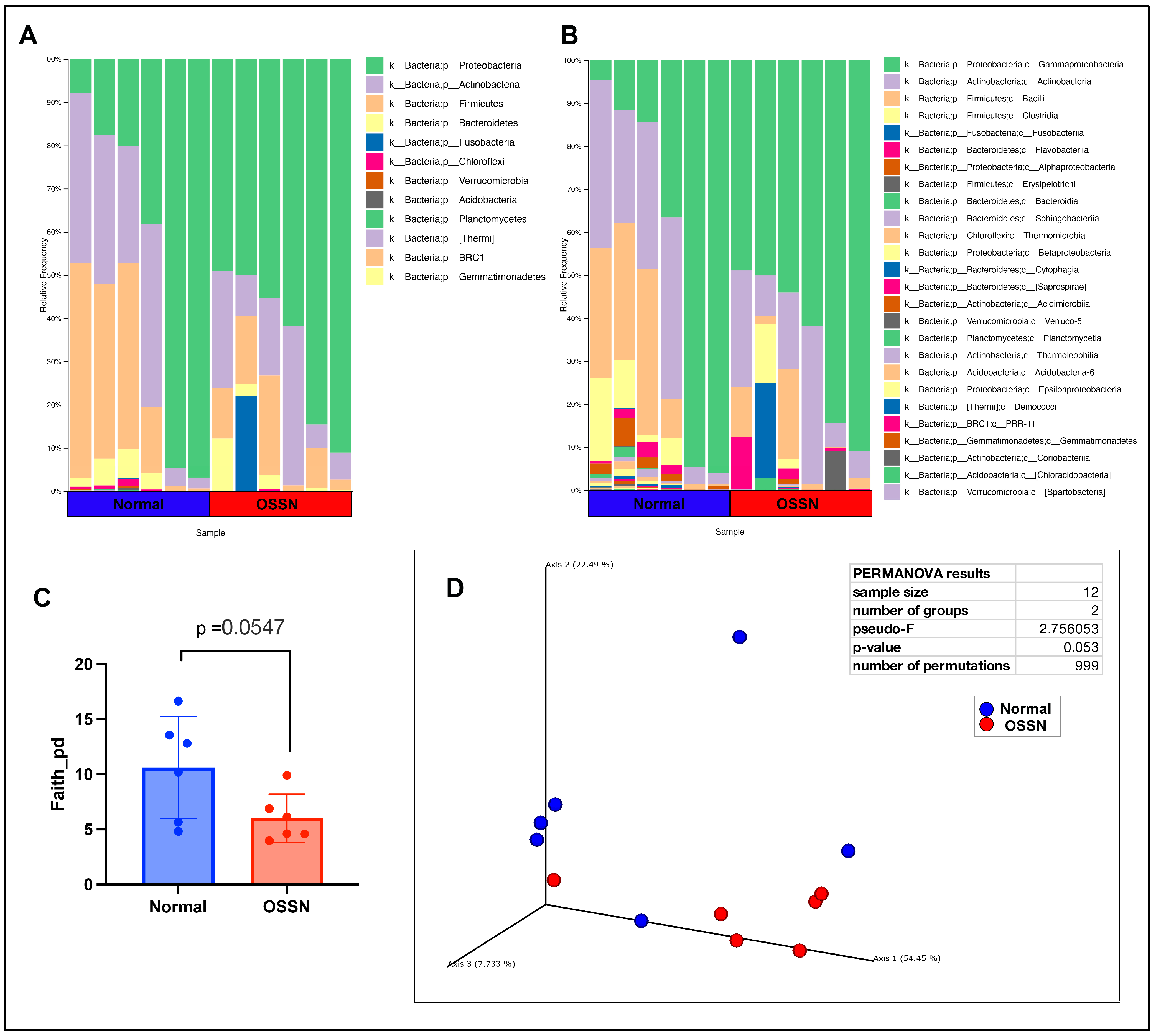
3.2. Ocular Microbiome of OSSN-Affected Horses
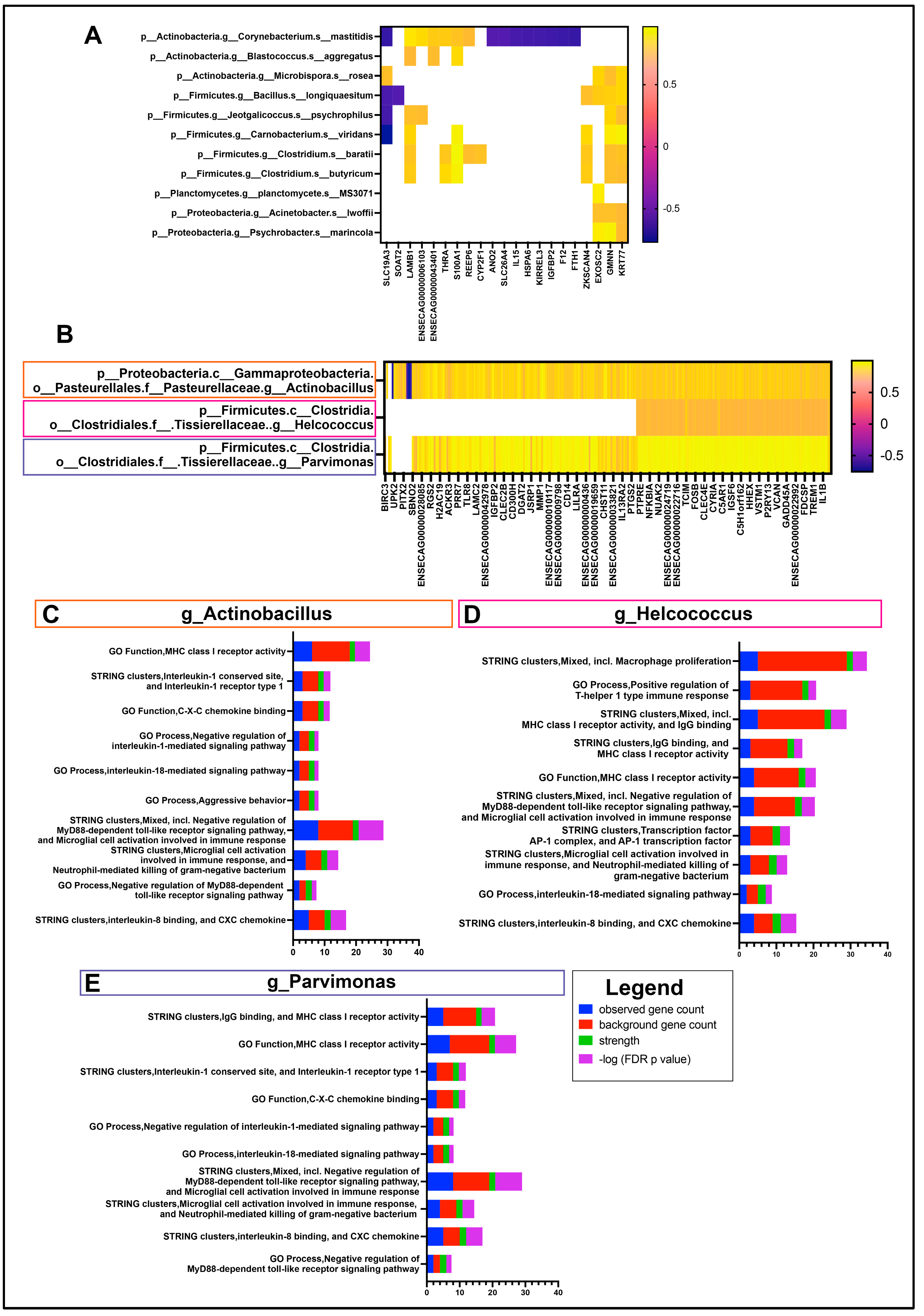
3.3. Interactome of OSSN
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kafarnik, C.; Rawlings, M.; Dubielzig, R.R. Corneal stromal invasive squamous cell carcinoma: A retrospective morphological description in 10 horses. Vet. Ophthalmol. 2009, 12, 6–12. [Google Scholar] [CrossRef]
- Wilcock, B.P. The eye and ear. Pathol. Domest. Anim. 1993, 441–529. [Google Scholar]
- Blodi, F.C.; Ramsey, F.K. Ocular tumors in domestic animals. Am. J. Ophthalmol. 1967, 64, 627–633. [Google Scholar] [CrossRef] [PubMed]
- McInnis, C.L.; Giuliano, E.A.; Johnson, P.J.; Turk, J.R. Immuno-histochemical evaluation of cyclooxygenase expression in corneal squamous cell carcinoma in horses. Am. J. Vet. Res. 2007, 68, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Dugan, S.J.; Curtis, C.R.; Roberts, S.M.; Severin, G.A. Epidemiologic study of ocular/adnexal squamous cell carcinoma in horses. J. Am. Vet. Med. Assoc. 1991, 198, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Dugan, S.J.; Roberts, S.M.; Curtis, C.R.; Severin, G.A. Prognostic factors and survival of horses with ocular/adnexal squamous cell carcinoma: 147 cases (1978–1988). J. Am. Vet. Med. Assoc. 1991, 198, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Sykora, S.; Brandt, S. Papillomavirus infection and squamous cell carcinoma in horses. Vet. J. 2017, 223, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Rassnick, K.M.; Njaa, B.L. Cyclooxygenase-2 immunoreactivity in equine ocular squamous-cell carcinoma. J. Vet. Diagn. Investig. 2007, 19, 436–439. [Google Scholar] [CrossRef]
- Crausaz, M.; Launois, T.; Smith-Fleming, K.; McCoy, A.M.; Knickelbein, K.E.; Bellone, R.R. DDB2 genetic risk factor for ocular squamous cell carcinoma identified in three additional horse breeds. Genes 2020, 11, 1460. [Google Scholar] [CrossRef]
- Bellone, R.R. Genetics of equine ocular disease. Vet. Clin. N. Am. Equine Pract. 2020, 36, 303–322. [Google Scholar] [CrossRef]
- Bellone, R.R.; Liu, J.; Petersen, J.L.; Mack, M.; Singer-Berk, M.; Drögemüller, C.; Malvick, J.; Wallner, B.; Brem, G.; Penedo, M.C. A missense mutation in damage-specific DNA binding protein 2 is a genetic risk factor for limbal squamous cell carcinoma in horses. Int. J. Cancer 2017, 141, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Knickelbein, K.E.; Lassaline, M.E.; Singer-Berk, M.; Reilly, C.M.; Clode, A.B.; Famula, T.R.; Michau, T.M.; Bellone, R.R. A missense mutation in damage-specific DNA binding protein 2 is a genetic risk factor for ocular squamous cell carcinoma in Belgian horses. Equine Vet. J. 2020, 52, 34–40. [Google Scholar] [CrossRef] [PubMed]
- King, T.C.; Priehs, D.R.; Gum, G.G.; Miller, T.R. Therapeutic management of ocular squamous cell carcinoma in the horse: 43 cases (1979–1989). Equine Vet. J. 1991, 23, 449–452. [Google Scholar] [CrossRef]
- Mosunic, C.B.; Moore, P.A.; Carmicheal, K.P.; Chandler, M.J.; Vidyashankar, A.; Zhao, Y.; Roberts, R.E.; Dietrich, U.M. Effects of treatment with and without adjuvant radiation therapy on recurrence of ocular and adnexal squamous cell carcinoma in horses: 157 cases (1985–2002). J. Am. Vet. Med. Assoc. 2004, 225, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- McCalla, T.L.; Moore, C.P.; Collier, L.L. Immunotherapy of periocular squamous cell carcinoma with metastasis in a pony. J. Am. Vet. Med. Assoc. 1992, 200, 1678–1681. [Google Scholar] [CrossRef]
- Théon, A.P.; Pascoe, J.R.; Meagher, D.M. Perioperative intratumoral administration of cisplatin for treatment of cutaneous tumors in equidae. J. Am. Vet. Med. Assoc. 1994, 205, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- Théon, A.P.; Pascoe, J.R. Iridium-192 interstitial brachytherapy for equine periocular tumours: Treatment results and prognostic factors in 115 horses. Equine Vet. J. 1995, 27, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Théon, A.P.; Pascoe, J.R.; Madigan, J.E.; Carlson, G.; Metzger, L. Comparison of intratumoral administration of cisplatin versus bleomycin for treatment of periocular squamous cell carcinomas in horses. Am. J. Vet. Res. 1997, 58, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, D.A.; Burt, J.K. Combined treatment of ocular squamous cell carcinoma in a horse, using radiofrequency hyperthermia and interstitial 198Au implants. J. Am. Vet. Med. Assoc. 1990, 196, 1831–1833. [Google Scholar] [CrossRef] [PubMed]
- English, R.V.; Nasisse, M.P.; Davidson, M.G. Carbon dioxide laser ablation for treatment of limbal squamous cell carcinoma in horses. J. Am. Vet. Med. Assoc. 1990, 196, 439–442. [Google Scholar] [CrossRef]
- Walker, M.A.; Schumacher, J.; Schmitz, D.G.; McMullen, W.C.; Ruoff, W.W.; Crabill, M.R.; Hawkins, J.F.; Hogan, P.M.; McClure, S.R.; Vacek, J.R. Cobalt 60 radiotherapy for treatment of squamous cell carcinoma of the nasal cavity and paranasal sinuses in three horses. J. Am. Vet. Med. Assoc. 1998, 212, 848–851. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, T.; Ruppin, M.; Wheeless, M.; Williams, S.; Reddell, P. Use of the intratumoural anticancer drug tigilanol tiglate in two horses. Front. Vet. Sci. 2020, 7, 639. [Google Scholar] [CrossRef] [PubMed]
- Estell, K. Periocular neoplasia in the horse. Vet. Clin. N. Am. Equine Pract. 2017, 33, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Elce, Y.A.; Orsini, J.A.; Blikslager, A.T. Expression of cyclooxy-genase-1 and -2 in naturally occurring squamous cell carcinomas in horses. Am. J. Vet. Res. 2007, 68, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Howarth, S.; Lucke, V.M.; Pearson, H. Squamous cell carcinoma of the equine external genitalia: A review and assessment of penile amputation and urethrostomy as a surgical treatment. Equine Vet. J. 1991, 23, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, D.V.H. Equine ocular squamous cell carcinoma. Clin. Tech. Equine Pract. 2005, 4, 87–94. [Google Scholar] [CrossRef]
- Wotman, K.; Chow, L.; Martabano, B.; Pezzanite, L.M.; Dow, S. Novel ocular immunotherapy induces tumor regression in an equine model of ocular surface squamous neoplasia. Cancer Immunol. Immunother. 2023, 72, 1185–1198. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hui, L.; Chen, Y. Tumor microenvironment: Sanctuary of the devil. Cancer Lett. 2015, 368, 7–13. [Google Scholar] [CrossRef]
- Reichman, H.; Karo-Atar, D.; Munitz, A. Emerging Roles for Eosinophils in the Tumor Microenvironment. Trends Cancer 2016, 2, 664–675. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Jobin, C. The microbiome and cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.; Backhed, F.; Levin, B.R.; McFall-Ngai, M.J.; McLean, A.R. Evolution, human-microbe interactions, and life history plasticity. Lancet 2017, 390, 521–530. [Google Scholar] [CrossRef]
- Davenport, E.R.; Sanders, J.G.; Song, S.J.; Amato, K.R.; Clark, A.G.; Knight, R. The human microbiome in evolution. BMC Biol. 2017, 15, 127. [Google Scholar] [CrossRef] [PubMed]
- Nené, N.R.; Reisel, D.; Leimbach, A.; Franchi, D.; Jones, A.; Evans, I. Association between the cervicovaginal microbiome, BRCA1 mutation status, and risk of ovarian cancer: A case-control study. Lancet Oncol. 2019, 20, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wang, Z.; Li, C.; Lv, K.; Tian, G.; Tang, M. Bacterial biomarkers capable of identifying recurrence or metastasis carry disease severity information for lung cancer. Front. Microbiol. 2022, 13, 1007831. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, J.; Willcox, M.D. The Ocular Microbiome: Molecular Characterization of a Unique and Low Microbial Environment. Curr. Eye Res. 2019, 44, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wei, J. Identification of Pathogen Signatures in Prostate Cancer Using RNA-seq. PLoS ONE 2015, 10, e0128955. [Google Scholar] [CrossRef] [PubMed]
- Pevsner-Fischer, M.; Tuganbaev, T.; Meijer, M.; Zhang, S.H.; Zeng, Z.R.; Chen, M.H. Role of the microbiome in non-gastrointestinal cancers. World J. Clin. Oncol. 2016, 7, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Zackular, J.P.; Rogers, M.A.; Ruffin, M.T.; Schloss, P.D. The human gut microbiome as a screening tool for colorectal cancer. Cancer Prev. Res. 2014, 7, 1112–1121. [Google Scholar] [CrossRef]
- Erb-Downward, J.R.; Thompson, D.L.; Han, M.K.; Freeman, C.M.; McCloskey, L.; Schmidt, L.A. Analysis of the lung microbiome in the “healthy” smoker and in COPD. PLoS ONE 2011, 6, e16384. [Google Scholar] [CrossRef] [PubMed]
- Hieken, T.J.; Chen, J.; Hoskin, T.L.; Walther-Antonio, M.; Johnson, S.; Ramaker, S. The Microbiome of Aseptically Collected Human Breast Tissue in Benign and Malignant Disease. Sci. Rep. 2016, 6, 30751. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The Microbiota of Breast Tissue and Its Association with Breast Cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef] [PubMed]
- Xuan, C.; Shamonki, J.M.; Chung, A.; Dinome, M.L.; Chung, M.; Sieling, P.A. Microbial dysbiosis is associated with human breast cancer. PLoS ONE 2014, 9, e83744. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.J.; Ingle, J.N.; Tang, X.; Chia, N.; Jeraldo, P.R.; Walther-Antonio, M.R.; Kandimalla, K.K.; Johnson, S.; Yao, J.Z.; Harrington, S.C.; et al. A comprehensive analysis of breast cancer microbiota and host gene expression. PLoS ONE 2017, 30, e0188873. [Google Scholar] [CrossRef] [PubMed]
- Nino, J.L.G.; LaCourse, K.D.; Kempchinsky, A.G.; Baryliames, A.; Barber, B.; Futran, N.; Houlton, J.; Sather, C.; Sicinska, E.; Taylor, A.; et al. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature 2022, 611, 810–817. [Google Scholar] [CrossRef]
- Chen, F.; Yang, J.; Guo, Y.; Su, D.; Sheng, Y.; Wu, Y. Integrating bulk and single-cell RNA sequencing data reveals the relationship between intratumor microbiome signature and host metabolic heterogeneity in breast cancer. Front. Immunol. 2023, 14, 1140995. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.Å.; Knight, R.T.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2022. [Google Scholar]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING V10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Ishibashi, T. Hyalocytes: Essential cells of the vitreous cavity in vitroretinal pathphysiology? Retina 2011, 31, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Boneva, S.; Rosmus, D.; Agostini, H.; Schlunch, G.; Weighofer, P.; Schlecht, A.; Lange, C. Deciphering the molecular signature of human hyalocytes in relation to other innate immune cell populations. Investig. Ophthalmol. Vis. Sci. 2022, 63, 9. [Google Scholar] [CrossRef] [PubMed]
- Gichuhi, S.; Ohnuma, S.; Sagoo, M.S.; Burton, M. Pathophysiology of ocular surface squamous neoplasia. Exp. Eye Res. 2014, 129, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.S.; Vora, G.K.; Gupta, P.K. Anterior segment imaging in ocular surface squamous neoplasia. J. Ophthalmol. 2016, 2016, 5435092. [Google Scholar] [CrossRef] [PubMed]
- Julius, P.; Siyumbwa, S.N.; Maate, F.; Moonga, P.; Kang, G.; Kaile, T.; West, J.T.; Wood, C.; Angeletti, P.C. Yes-associated protein-1 overexpression in ocular surface squamous neoplasia; a potential diagnostic marker and therapeutic target. Front. Oncol. 2023, 13, 1213426. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.M.; Del Piero, F.; Habecker, P.L.; Langohr, I.M. A retrospective histologic study of 140 cases of clinically significant equine ocular disorders. J. Vet. Diag Invest. 2020, 32, 382–388. [Google Scholar] [CrossRef]
- Zhou, P.; Zhou, J. The primary cilium as a therapeutic target in ocular diseases. Front. Pharmacol. 2020, 11, 977. [Google Scholar] [CrossRef]
- LaFrentz, S.; Abarca, E.; Mohammed, H.H.; Cuming, R.; Arias, C.R. Characterization of the normal equine conjunctival bacterial community using culture-independent methods. Vet. Ophthalmol. 2020, 23, 480–488. [Google Scholar] [CrossRef]
- Scott, E.M.; Arnold, C.; Dowell, S.; Suchodolski, J.S. Evaluation of the bacterial ocular surface microbiome in clinically normal horses before and after treatment with topical neomycin-polymyxin-bacitracin. PLoS ONE 2019, 14, e0214877. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.L.; Meason-Smith, C.; Arnold, C.; Suchodolski, J.S.; Scott, E.M. Evaluation of the ocular surface mycobiota in clinically normal horses. PLoS ONE 2021, 16, e0246537. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, R.; Lara, F.; Barros, T.M.; Mardones, E.; Cuadra, F.; Thomson, P. Ocular Microbiome in a Group of Clinically Healthy Horses. Animals 2022, 12, 943. [Google Scholar] [CrossRef]
- Julien, M.E.; Shih, J.B.; Correa Lopes, B.; Vallone, L.V.; Suchodolski, J.S.; Pilla, R. Alterations of the bacterial ocular surface microbiome are found in both eyes of horses with unilateral ulcerative keratitis. PLoS ONE 2023, 18, e0291028. [Google Scholar] [CrossRef] [PubMed]
- Delbeke, H.; Younas, S.; Casteels, I.; Joossens, M. Current knowledge on the human eye microbiome: A systematic review of available amplicon and metagenomic sequencing data. Acta Ophthalmologica 2020, 99, 16–25. [Google Scholar] [CrossRef]
- Gafen, H.B.; Liu, C.-C.; Ineck, N.E.; Scully, C.M.; Mironovich, M.A.; Guarneri, L.; Taylor, C.M.; Luo, M.; Leis, M.L.; Scott, E.M. Relative and Quantitative Characterization of the Bovine Bacterial Ocular Surface Microbiome in the Context of Suspected Ocular Squamous Cell Carcinoma. Animals 2023, 13, 1976. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, X.; Zhou, Y.; Fu, K.; Lau, H.C.-H.; Chun, T.W.-Y.; Cheung, A.H.-K.; Coker, O.O.; Wei, H.; Wu, W.K.-K.; et al. Parvimonas micra promotes colorectal tumorigenesis and is associated with prognosis of colorectal cancer patients. Oncogene 2002, 41, 4200–4210. [Google Scholar] [CrossRef]
- Bergsten, E.; Mestivier, D.; Donnadieu, F.; Pedron, T.; Barau, C.; Meda, L.T.; Mettouchi, A.; Lemichez, E.; Gorgette, O.; Chamaillard, M.; et al. Parvimonas micra, an oral pathobiont associated with colorectal cancer, epigenetically reprograms human colonocytes. Gut Microbes 2023, 15, 2265138. [Google Scholar] [CrossRef]
- Szoboszlay, M.; Schramm, L.; Pinzauti, D.; Scerri, J.; Sandionigi, A.; Biazzo, M. Nanopore Is Preferable over Illumina for 16S Amplicon Sequencing of the Gut Microbiota When Species-Level Taxonomic Classification, Accurate Estimation of Richness, or Focus on Rare Taxa Is Required. Microorganisms 2023, 11, 804. [Google Scholar] [CrossRef]
- Takan, I.; Karakülah, G.; Louka, A.; Pavlopoulou, A. “In the light of evolution”: Keratins as exceptional tumor biomarkers. PeerJ 2023, 11, e15099. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Huang, H.; Wang, B.; Jiang, S.; Guo, H.; Zhu, L.; Wu, S.; Liu, J.; Wang, L.; Lan, X.; et al. Comprehensive 3D epigenomic maps define limbal stem/progenitor cell function and identity. Nat. Commun. 2022, 13, 1293. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, J.; Willcox, M.; Wemheuer, B.; Wilcsek, G.; Coroneo, M.; Thomas, T. Biogeography of the human ocular microbiota. Ocul. Surf. 2019, 17, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Herzog, E.L.; Kreuzer, M.; Zinkernagel, M.S.; Zysset-Burri, D.C. Challenges and insights in the exploration fo the low abundance human ocular surface microbiome. Front. Cell Infect. Microbiol. 2023, 13, 1232147. [Google Scholar] [CrossRef]
- Auten, C.R.; Urbanz, J.L.; Dees, D.D. Comparison of bacterial culture results collected via direct corneal ulcer vs conjunctival fornix sampling in canine eyes with presumed bacterial ulcerative keratitis. Vet. Opthalmol. 2019, 23, 135–140. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chow, L.; Flaherty, E.; Pezzanite, L.; Williams, M.; Dow, S.; Wotman, K. Impact of Equine Ocular Surface Squamous Neoplasia on Interactions between Ocular Transcriptome and Microbiome. Vet. Sci. 2024, 11, 167. https://doi.org/10.3390/vetsci11040167
Chow L, Flaherty E, Pezzanite L, Williams M, Dow S, Wotman K. Impact of Equine Ocular Surface Squamous Neoplasia on Interactions between Ocular Transcriptome and Microbiome. Veterinary Sciences. 2024; 11(4):167. https://doi.org/10.3390/vetsci11040167
Chicago/Turabian StyleChow, Lyndah, Edward Flaherty, Lynn Pezzanite, Maggie Williams, Steven Dow, and Kathryn Wotman. 2024. "Impact of Equine Ocular Surface Squamous Neoplasia on Interactions between Ocular Transcriptome and Microbiome" Veterinary Sciences 11, no. 4: 167. https://doi.org/10.3390/vetsci11040167
APA StyleChow, L., Flaherty, E., Pezzanite, L., Williams, M., Dow, S., & Wotman, K. (2024). Impact of Equine Ocular Surface Squamous Neoplasia on Interactions between Ocular Transcriptome and Microbiome. Veterinary Sciences, 11(4), 167. https://doi.org/10.3390/vetsci11040167