


The Novel Role of the NLRP3 Inflammasome in Mycotoxin-Induced Toxicological Mechanisms

Abstract
Simple Summary
Abstract
1. Introduction
2. Mycotoxicosis
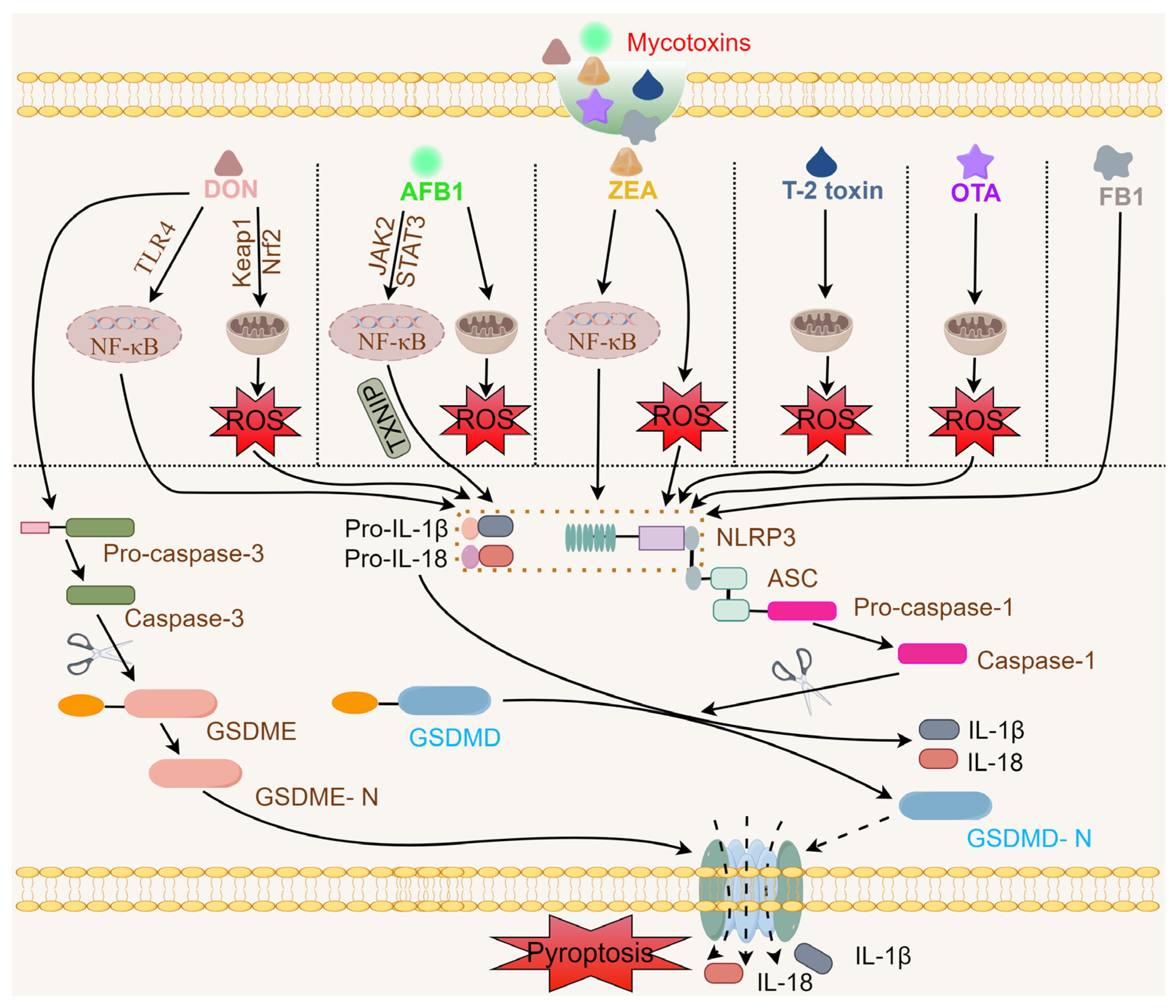
3. Mycotoxins Activate the NLRP3 Inflammasome
3.1. DON and the NLRP3 Inflammasome
3.2. AFB1 and the NLRP3 Inflammasome
3.3. ZEA and the NLRP3 Inflammasome
3.4. T-2 Toxin and the NLRP3 Inflammasome
3.5. OTA and the NLRP3 Inflammasome
3.6. FB1 and the NLRP3 Inflammasome
4. Targeting the NLRP3 Inflammasome for Mycotoxin Exposure
5. Conclusions and Future Perspectives
- (1)
- As mentioned above, mycotoxins trigger NLRP3 inflammasome activation via the canonical activation pathway. Although evidence suggests that exogenous stimuli such as DON, AFB1, ZEA, OTA, T-2 toxin, FB1, or patulin can trigger NLRP3 inflammasome activation, whether these mycotoxins act directly on NLRP3 remains unclear. Previous studies have investigated alterations in the expression of factors involved in generation of NLRP3 inflammasomes in response to mycotoxin exposure, but the exact mechanisms and signaling pathways by which mycotoxins induce NLRP3 inflammasome activation have remained elusive.
- (2)
- Most of the research on NLRP3 inflammasome activation has focused on DON, AFB1, and ZEA, whereas studies of OTA, T-2 toxin, FB1, mycophenolic acid, and patulin are scarce; alternaria, HT-2 toxins, citrinin, enniatins, ergot alkaloids, and nivalenol have not been studied at all. Based on the existing research, it is currently not possible to ascribe a set of common factors to the activation of NLRP3 inflammasomes triggered by all 500 mycotoxins. Therefore, a wide range of mycotoxins as a subject of future study is necessary to improve understanding of the general rules for mycotoxin-induced NLRP3 inflammasome activation.
- (3)
- In recent years, more than 20 pharmacological inhibitors of NLRP3 inflammasomes have been reported, some of which exhibit promising therapeutic potential for treating NLRP3-related diseases in the clinic. Undeniably, all studies have clearly indicated that MCC950, BAY 11-7082, curcumin, and luteolin ameliorate tissue damage and inflammatory responses in models of mycotoxin exposure. Theoretically, any molecule or signal involved in the activation of NLRP3 inflammasomes, such as active caspase-1, GSDMD cleavage, proteins involved in inflammasome assembly, and inflammatory cytokines, can potentially inhibit the NLRP3 inflammasome. These are the two major mechanisms of small molecule inhibitors: they directly interact with the NLRP3 protein and bind to the ATP-binding motifs of the NACHT domain of NLRP3, subsequently inhibiting ATPase activity. In addition to the inhibitors mentioned above, other kinds of NLRP3 inflammasome inhibitors should be designed to investigate their effects on mycotoxin exposure. It will be worth examining whether the occurrence of chemical reactions between these inhibitors and mycotoxins reduces their toxic effects. For example, the carbon-carbon double bonds in BAY 11-7082 can react with the amino group in FB1 via Michael addition, and the sulfonyl group in MCC950 can react with the carboxyl group in FB1 via amide bond-forming reactions.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdallah, M.F.; Gado, M.; Abdelsadek, D.; Zahran, F.; El-Salhey, N.N.; Mehrez, O.; Abdel-Hay, S.; Mohamed, S.M.; De Ruyck, K.; Yang, S.; et al. Mycotoxin contamination in the Arab world: Highlighting the main knowledge gaps and the current legislation. Mycotoxin Res. 2024, 40, 19–44. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, K.; Long, M.; Yang, S.; Zhang, Y. An update on immunotoxicity and mechanisms of action of six environmental mycotoxins. Food Chem. Toxicol. 2022, 163, 112895. [Google Scholar] [CrossRef] [PubMed]
- Raduly, Z.; Szabo, A.; Mezes, M.; Balatoni, I.; Price, R.G.; Dockrell, M.E.; Pocsi, I.; Csernoch, L. New perspectives in application of kidney biomarkers in mycotoxin induced nephrotoxicity, with a particular focus on domestic pigs. Front. Microbiol. 2023, 14, 1085818. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mao, X.; Liu, K.; Sun, J.; Li, B.; Malyar, R.M.; Liu, D.; Pan, C.; Gan, F.; Liu, Y.; et al. Ochratoxin A induces nephrotoxicity in vitro and in vivo via pyroptosis. Arch. Toxicol. 2021, 95, 1489–1502. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, L.; Shi, L.; Yao, K.; Sun, X.; Yang, G.; Jiang, L.; Zhang, C.; Wang, N.; Zhang, H.; et al. Zearalenone induces NLRP3-dependent pyroptosis via activation of NF-kappaB modulated by autophagy in INS-1 cells. Toxicology 2019, 428, 152304. [Google Scholar] [CrossRef]
- Su, D.; Jiang, W.; Yuan, Q.; Guo, L.; Liu, Q.; Zhang, M.; Kang, C.; Xiao, C.; Yang, C.; Li, L.; et al. Chronic exposure to aflatoxin B1 increases hippocampal microglial pyroptosis and vulnerability to stress in mice. Ecotoxicol. Env. Saf. 2023, 258, 114991. [Google Scholar] [CrossRef] [PubMed]
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Stinnett, G.S.; Kuo, C.H.; Ono, S.J. Impact of inflammasomes on the ocular surface. Curr. Opin. Allergy Clin. Immunol. 2024, 10–1097. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, X.; Liu, W.; Hu, Q.; Yang, H. Targeting Pyroptosis through Lipopolysaccharide-Triggered Noncanonical Pathway for Safe and Efficient Cancer Immunotherapy. Nano Lett. 2023, 23, 8725–8733. [Google Scholar] [CrossRef]
- Evavold, C.L.; Hafner-Bratkovic, I.; Devant, P.; D’Andrea, J.M.; Ngwa, E.M.; Borsic, E.; Doench, J.G.; LaFleur, M.W.; Sharpe, A.H.; Thiagarajah, J.R.; et al. Control of gasdermin D oligomerization and pyroptosis by the Ragulator-Rag-mTORC1 pathway. Cell 2021, 184, 4495–4511.e4419. [Google Scholar] [CrossRef]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, J.; Zhou, X.; Lu, Y.; Cui, W.; Miao, L. Mini-Review: GSDME-Mediated Pyroptosis in Diabetic Nephropathy. Front. Pharmacol. 2021, 12, 780790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, W.; Li, W.; Zhao, Y. NLRP3 Inflammasome: Checkpoint Connecting Innate and Adaptive Immunity in Autoimmune Diseases. Front. Immunol. 2021, 12, 732933. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhou, Y.; Ning, X.; Li, S.; Xue, D.; Wei, C.; Zhu, Z.; Sheng, L.; Lu, B.; Li, Y.; et al. Directly targeting ASC by lonidamine alleviates inflammasome-driven diseases. J. Neuroinflammation 2022, 19, 315. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajslova, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Env. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed]
- Arce-Lopez, B.; Coton, M.; Coton, E.; Hymery, N. Occurrence of the two major regulated mycotoxins, ochratoxin A and fumonisin B1, in cereal and cereal-based products in Europe and toxicological effects: A review. Env. Toxicol. Pharmacol. 2024, 109, 104489. [Google Scholar] [CrossRef]
- Song, C.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Neurotoxic mechanisms of mycotoxins: Focus on aflatoxin B1 and T-2 toxin. Env. Pollut. 2024, 356, 124359. [Google Scholar] [CrossRef]
- Agahi, F.; Juan, C.; Font, G.; Juan-Garcia, A. Neurotoxicity of zearalenone’s metabolites and beauvericin mycotoxins via apoptosis and cell cycle disruption. Toxicology 2021, 456, 152784. [Google Scholar] [CrossRef]
- Alonso-Garrido, M.; Frangiamone, M.; Font, G.; Cimbalo, A.; Manyes, L. In vitro blood brain barrier exposure to mycotoxins and carotenoids pumpkin extract alters mitochondrial gene expression and oxidative stress. Food Chem. Toxicol. 2021, 153, 112261. [Google Scholar] [CrossRef]
- Tonshin, A.A.; Teplova, V.V.; Andersson, M.A.; Salkinoja-Salonen, M.S. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by affecting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology 2010, 276, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, Y.; Abraham, N.; Li, X.Z.; Kimber, M.; Zhou, T. The Ribosome-Binding Mode of Trichothecene Mycotoxins Rationalizes Their Structure-Activity Relationships. Int. J. Mol. Sci. 2021, 22, 1604. [Google Scholar] [CrossRef] [PubMed]
- Janevska, S.; Ferling, I.; Jojic, K.; Rautschek, J.; Hoefgen, S.; Proctor, R.H.; Hillmann, F.; Valiante, V. Self-Protection against the Sphingolipid Biosynthesis Inhibitor Fumonisin B(1) Is Conferred by a FUM Cluster-Encoded Ceramide Synthase. mBio 2020, 11, 1110–1128. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, D.; Fayjaloun, S.; Nassar, M.; Sahakian, J.; Aad, P.Y. Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals. Toxins 2019, 11, 515. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.L.; Dowe, R.J.; Geronimo, I.H.; Bender, W. Evidence for an indirect mechanism of aflatoxin B1 inhibition of rat liver nuclear RNA polymerase II activity in vivo. Carcinogenesis 1986, 7, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Li, L.; Makarova, A.V.; Burgers, P.M.; Stone, M.P.; Lloyd, R.S. Error-prone replication bypass of the primary aflatoxin B1 DNA adduct, AFB1-N7-Gua. J. Biol. Chem. 2014, 289, 18497–18506. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, K.I.; Kinoshita, M.; Furusawa, H.; Sato, K.; Honma, M. Epigenetic effect of the mycotoxin fumonisin B1 on DNA methylation. Mutagenesis 2021, 36, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Manyes, L. Toxicity of mycotoxins in vivo on vertebrate organisms: A review. Food Chem. Toxicol. 2020, 137, 111161. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal Transduct. Target. Ther. 2021, 6, 128. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef] [PubMed]
- Kodi, T.; Sankhe, R.; Gopinathan, A.; Nandakumar, K.; Kishore, A. New Insights on NLRP3 Inflammasome: Mechanisms of Activation, Inhibition, and Epigenetic Regulation. J. Neuroimmune Pharmacol. 2024, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Perregaux, D.G.; McNiff, P.; Laliberte, R.; Hawryluk, N.; Peurano, H.; Stam, E.; Eggler, J.; Griffiths, R.; Dombroski, M.A.; Gabel, C.A. Identification and characterization of a novel class of interleukin-1 post-translational processing inhibitors. J. Pharmacol. Exp. Ther. 2001, 299, 187–197. [Google Scholar] [PubMed]
- Fu, J.; Wu, H. Structural Mechanisms of NLRP3 Inflammasome Assembly and Activation. Annu. Rev. Immunol. 2023, 41, 301–316. [Google Scholar] [CrossRef]
- Downs, K.P.; Nguyen, H.; Dorfleutner, A.; Stehlik, C. An overview of the non-canonical inflammasome. Mol. Asp. Med. 2020, 76, 100924. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef]
- Lupfer, C.R.; Anand, P.K.; Liu, Z.; Stokes, K.L.; Vogel, P.; Lamkanfi, M.; Kanneganti, T.D. Reactive oxygen species regulate caspase-11 expression and activation of the non-canonical NLRP3 inflammasome during enteric pathogen infection. PLoS Pathog. 2014, 10, e1004410. [Google Scholar] [CrossRef]
- Zhu, F.; Ma, J.; Li, W.; Liu, Q.; Qin, X.; Qian, Y.; Wang, C.; Zhang, Y.; Li, Y.; Jiang, D.; et al. The orphan receptor Nur77 binds cytoplasmic LPS to activate the non-canonical NLRP3 inflammasome. Immunity 2023, 56, 753–767.E8. [Google Scholar] [CrossRef]
- Jalilzadeh-Amin, G.; Dalir-Naghadeh, B.; Ahmadnejad-Asl-Gavgani, M.; Fallah, A.A.; Mousavi Khaneghah, A. Prevalence and Concentration of Mycotoxins in Animal Feed in the Middle East and North Africa (MENA): A Systematic Review and Meta-Analysis. Toxins 2023, 15, 214. [Google Scholar] [CrossRef]
- Franchino, C.; Vita, V.; Iammarino, M.; De Pace, R. Monitoring of Animal Feed Contamination by Mycotoxins: Results of Five Years of Official Control by an Accredited Italian Laboratory. Microorganisms 2024, 12, 173. [Google Scholar] [CrossRef]
- Munoz-Solano, B.; Gonzalez-Penas, E. Co-Occurrence of Mycotoxins in Feed for Cattle, Pigs, Poultry, and Sheep in Navarra, a Region of Northern Spain. Toxins 2023, 15, 172. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Park, S.Y.; Kim, Y.H.; Kim, S.J.; Han, J.H. Occurrence of mycotoxins in swine feed from South Korea. J. Adv. Vet. Anim. Res. 2024, 11, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, B.; Wang, L.; Li, X.; Nawaz, M.Y.; Saleemi, M.K.; Khatoon, A.; Yongping, X. Recalling the reported toxicity assessment of deoxynivalenol, mitigating strategies and its toxicity mechanisms: Comprehensive review. Chem. Biol. Interact. 2024, 387, 110799. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Peng, Z.; Chen, L.; Nussler, A.K.; Liu, L.; Yang, W. Deoxynivalenol, gut microbiota and immunotoxicity: A potential approach? Food Chem. Toxicol. 2018, 112, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.H.; Shin, S.; Park, J.; Lee, B.R.; Lee, S.I. Pyroptosis-Mediated Damage Mechanism by Deoxynivalenol in Porcine Small Intestinal Epithelial Cells. Toxins 2023, 15, 300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, F.; Wang, Y.; Zhang, K.; Yang, X.; Wang, X. Tanshinone IIA protects intestinal epithelial cells from deoxynivalenol-induced pyroptosis. Ecotoxicol. Env. Saf. 2024, 269, 115743. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zhang, S.; He, W.; Liu, S.; Mao, X.; Yin, L.; Yue, D.; Zhang, P.; Huang, K.; Chen, X. Crucial Function of Caveolin-1 in Deoxynivalenol-Induced Enterotoxicity by Activating ROS-Dependent NLRP3 Inflammasome-Mediated Pyroptosis. J. Agric. Food Chem. 2022, 70, 12968–12981. [Google Scholar] [CrossRef]
- Ge, L.; Lin, Z.; Le, G.; Hou, L.; Mao, X.; Liu, S.; Liu, D.; Gan, F.; Huang, K. Nontoxic-dose deoxynivalenol aggravates lipopolysaccharides-induced inflammation and tight junction disorder in IPEC-J2 cells through activation of NF-kappaB and LC3B. Food Chem. Toxicol. 2020, 145, 111712. [Google Scholar] [CrossRef]
- Ge, L.; Liu, D.; Mao, X.; Liu, S.; Guo, J.; Hou, L.; Chen, X.; Huang, K. Low Dose of Deoxynivalenol Aggravates Intestinal Inflammation and Barrier Dysfunction Induced by Enterotoxigenic Escherichia coli Infection through Activating Macroautophagy/NLRP3 Inflammasomes. J. Agric. Food Chem. 2022, 70, 3009–3022. [Google Scholar] [CrossRef]
- Song, J.L.; Sun, Y.J.; Liu, G.Q.; Zhang, G.L. Deoxynivalenol and zearalenone: Different mycotoxins with different toxic effects in donkey (Equus asinus) endometrial epithelial cells. Theriogenology 2022, 179, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Molagoda, I.M.N.; Lee, S.; Jayasooriya, R.; Jin, C.Y.; Choi, Y.H.; Kim, G.Y. Deoxynivalenol enhances IL-1ss expression in BV2 microglial cells through activation of the NF-?B pathway and the ASC/NLRP3 inflammasome. EXCLI J. 2019, 18, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.; Liu, W.; Xu, Y.; Zhang, B.; Meng, L.; Yin, R.; Chen, H. New insights into micro-algal astaxanthin’s effect on deoxynivalenol-induced spleen lymphocytes pyroptosis in Cyprinus carpio: Involving mitophagy and mtROS-NF-kappaB-dependent NLRP3 inflammasome. Fish. Shellfish. Immunol. 2024, 144, 109259. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, Z.; Wu, N.; Li, J.; Xi, N.; Xu, M.; Wu, F.; Fu, Q.; Yan, G.; Liu, Y.; et al. Chlorogenic acid attenuates deoxynivalenol-induced apoptosis and pyroptosis in human keratinocytes via activating Nrf2/HO-1 and inhibiting MAPK/NF-kappaB/NLRP3 pathways. Biomed. Pharmacother. 2024, 170, 116003. [Google Scholar] [CrossRef]
- Rogers, C.; Fernandes-Alnemri, T.; Mayes, L.; Alnemri, D.; Cingolani, G.; Alnemri, E.S. Cleavage of DFNA5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat. Commun. 2017, 8, 14128. [Google Scholar] [CrossRef]
- Mao, X.; Li, J.; Xie, X.; Chen, S.; Huang, Q.; Mu, P.; Jiang, J.; Deng, Y. Deoxynivalenol induces caspase-3/GSDME-dependent pyroptosis and inflammation in mouse liver and HepaRG cells. Arch. Toxicol. 2022, 96, 3091–3112. [Google Scholar] [CrossRef]
- Cai, G.; Guerrero-Netro, H.M.; Bian, J.; Oswald, I.P.; Price, C.; Alassane-Kpembi, I. Real-life exposure to Fusarium toxins deoxynivalenol and zearalenone triggers apoptosis and activates NLRP3 inflammasome in bovine primary theca cells. Mycotoxin Res. 2023, 39, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Zhang, P.; Du, H.; Ge, L.; Liu, S.; Huang, K.; Chen, X. The combined effect of deoxynivalenol and Fumonisin B(1) on small intestinal inflammation mediated by pyroptosis in vivo and in vitro. Toxicol. Lett. 2023, 372, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, Q.; He, W.; Ge, L.; Huang, K. Deoxynivalenol aggravates the immunosuppression in piglets and PAMs under the condition of PEDV infection through inhibiting TLR4/NLRP3 signaling pathway. Ecotoxicol. Env. Saf. 2022, 231, 113209. [Google Scholar] [CrossRef]
- Liu, D.; Wang, Q.; He, W.; Chen, X.; Wei, Z.; Huang, K. Two-way immune effects of deoxynivalenol in weaned piglets and porcine alveolar macrophages: Due mainly to its exposure dosage. Chemosphere 2020, 249, 126464. [Google Scholar] [CrossRef]
- Song, J.L.; Zhang, G.L. Deoxynivalenol and Zearalenone: Different Mycotoxins with Different Toxic Effects in the Sertoli Cells of Equus asinus. Cells 2021, 10, 1898. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Li, Q. Aflatoxin B(1) in poultry liver: Toxic mechanism. Toxicon 2023, 233, 107262. [Google Scholar] [CrossRef]
- Dai, C.; Tian, E.; Hao, Z.; Tang, S.; Wang, Z.; Sharma, G.; Jiang, H.; Shen, J. Aflatoxin B1 Toxicity and Protective Effects of Curcumin: Molecular Mechanisms and Clinical Implications. Antioxidants 2022, 11, 2031. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Zhan, D.L.; Chen, Y.Y.; Wang, W.H.; He, C.Y.; Lin, Y.; Lin, Y.C.; Lin, Z.N. Aflatoxin B1 enhances pyroptosis of hepatocytes and activation of Kupffer cells to promote liver inflammatory injury via dephosphorylation of cyclooxygenase-2: An in vitro, ex vivo and in vivo study. Arch. Toxicol. 2019, 93, 3305–3320. [Google Scholar] [CrossRef]
- Lv, H.; Liu, J.; He, Y.; Xia, S.; Qiao, C.; Xu, C. The Ameliorative Role of Lico A on Aflatoxin B(1)-Triggered Hepatotoxicity Partially by Activating Nrf2 Signal Pathway. J. Agric. Food Chem. 2024, 72, 2741–2755. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Yang, Y.; Zhao, D.; Liu, R.; Li, S.; Zhang, X. Proteomic analysis of ITPR2 as a new therapeutic target for curcumin protection against AFB1-induced pyroptosis. Ecotoxicol. Env. Saf. 2023, 260, 115073. [Google Scholar] [CrossRef]
- Wang, Q.; Jia, F.; Guo, C.; Wang, Y.; Zhang, X.; Cui, Y.; Song, M.; Cao, Z.; Li, Y. PINK1/Parkin-mediated mitophagy as a protective mechanism against AFB(1)-induced liver injury in mice. Food Chem. Toxicol. 2022, 164, 113043. [Google Scholar] [CrossRef]
- Luo, T.; Zhou, X.; Qin, M.; Lin, Y.; Lin, J.; Chen, G.; Liu, A.; Ouyang, D.; Chen, D.; Pan, H. Corilagin Restrains NLRP3 Inflammasome Activation and Pyroptosis through the ROS/TXNIP/NLRP3 Pathway to Prevent Inflammation. Oxid. Med. Cell Longev. 2022, 2022, 1652244. [Google Scholar] [CrossRef]
- Zhong, Z.; Liang, S.; Sanchez-Lopez, E.; He, F.; Shalapour, S.; Lin, X.J.; Wong, J.; Ding, S.; Seki, E.; Schnabl, B.; et al. New mitochondrial DNA synthesis enables NLRP3 inflammasome activation. Nature 2018, 560, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; He, H.; Ma, M.; Zhou, R. Apilimod activates the NLRP3 inflammasome through lysosome-mediated mitochondrial damage. Front. Immunol. 2023, 14, 1128700. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Chen, H.; Wang, J.; Tsim, K.W.K.; Wang, Y.; Shen, X.; Lei, H.; Liu, Y. Aflatoxin B(1)-induced liver pyroptosis is mediated by disturbing the gut microbial metabolites: The roles of pipecolic acid and norepinephrine. J. Hazard. Mater. 2024, 474, 134822. [Google Scholar] [CrossRef]
- Chen, H.; Ye, L.; Wang, Y.; Chen, J.; Wang, J.; Li, X.; Lei, H.; Liu, Y. Aflatoxin B(1) exposure causes splenic pyroptosis by disturbing the gut microbiota-immune axis. Food Funct. 2024, 15, 3615–3628. [Google Scholar] [CrossRef]
- Jin, S.; Yang, H.; Wang, Y.; Pang, Q.; Jiao, Y.; Shan, A.; Feng, X. Dietary Curcumin Alleviated Aflatoxin B1-Induced Acute Liver Damage in Ducks by Regulating NLRP3-Caspase-1 Signaling Pathways. Foods 2021, 10, 3086. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, Q.; Zhang, X.; Yang, X.; Shi, Y.; Li, Y.; Song, M. Curcumin Alleviates Aflatoxin B(1)-Induced Liver Pyroptosis and Fibrosis by Regulating the JAK2/NLRP3 Signaling Pathway in Ducks. Foods 2023, 12, 1006. [Google Scholar] [CrossRef]
- Gao, X.; Jiang, L.; Xu, J.; Liu, W.; Li, S.; Huang, W.; Zhao, H.; Yang, Z.; Yu, X.; Wei, Z. Aflatoxin B1-activated heterophil extracellular traps result in the immunotoxicity to liver and kidney in chickens. Dev. Comp. Immunol. 2022, 128, 104325. [Google Scholar] [CrossRef]
- Li, S.; Liu, R.; Xia, S.; Wei, G.; Ishfaq, M.; Zhang, Y.; Zhang, X. Protective role of curcumin on aflatoxin B1-induced TLR4/RIPK pathway mediated-necroptosis and inflammation in chicken liver. Ecotoxicol. Env. Saf. 2022, 233, 113319. [Google Scholar] [CrossRef]
- Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-kappaB Signaling Pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef]
- Zhang, F.L.; Ma, H.H.; Dong, P.Y.; Yan, Y.C.; Chen, Y.; Yang, G.M.; Shen, W.; Zhang, X.F. Bacillus licheniformis ameliorates Aflatoxin B1-induced testicular damage by improving the gut-metabolism-testis axis. J. Hazard. Mater. 2024, 468, 133836. [Google Scholar] [CrossRef]
- Pan, H.; Hu, T.; He, Y.; Zhong, G.; Wu, S.; Jiang, X.; Rao, G.; You, Y.; Ruan, Z.; Tang, Z.; et al. Curcumin attenuates aflatoxin B1-induced ileum injury in ducks by inhibiting NLRP3 inflammasome and regulating TLR4/NF-kappaB signaling pathway. Mycotoxin Res. 2024, 40, 255–268. [Google Scholar] [CrossRef]
- Zhang, J.; Su, D.; Liu, Q.; Yuan, Q.; Ouyang, Z.; Wei, Y.; Xiao, C.; Li, L.; Yang, C.; Jiang, W.; et al. Gasdermin D-mediated microglial pyroptosis exacerbates neurotoxicity of aflatoxins B1 and M1 in mouse primary microglia and neuronal cultures. Neurotoxicology 2022, 91, 305–320. [Google Scholar] [CrossRef]
- Yan, H.; Ge, J.; Gao, H.; Pan, Y.; Hao, Y.; Li, J. Melatonin attenuates AFB1-induced cardiotoxicity via the NLRP3 signalling pathway. J. Int. Med. Res. 2020, 48, 300060520952656. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Huangfu, B.; Xu, T.; Xu, W.; Asakiya, C.; Huang, K.; He, X. Research Progress of Safety of Zearalenone: A Review. Toxins 2022, 14, 386. [Google Scholar] [CrossRef]
- Wang, T.; Ye, Y.; Ji, J.; Yang, X.; Xu, J.; Wang, J.S.; Han, X.; Zhang, T.; Sun, X. Diet composition affects long-term zearalenone exposure on the gut-blood-liver axis metabolic dysfunction in mice. Ecotoxicol. Env. Saf. 2022, 236, 113466. [Google Scholar] [CrossRef]
- Ding, C.; Fan, W.; Shen, T.; Huang, K.; Song, S.; Yan, L. Zearalenone can relieve dextran sulfate sodium-induced inflammatory reaction. J. Biochem. Mol. Toxicol. 2018, 33, e22236. [Google Scholar] [CrossRef]
- Yan, W.K.; Liu, Y.N.; Song, S.S.; Kang, J.W.; Zhang, Y.; Lu, L.; Wei, S.W.; Xu, Q.X.; Zhang, W.Q.; Liu, X.Z.; et al. Zearalenone affects the growth of endometriosis via estrogen signaling and inflammatory pathways. Ecotoxicol. Env. Saf. 2022, 241, 113826. [Google Scholar] [CrossRef]
- Fan, W.; Lv, Y.; Ren, S.; Shao, M.; Shen, T.; Huang, K.; Zhou, J.; Yan, L.; Song, S. Zearalenone (ZEA)-induced intestinal inflammation is mediated by the NLRP3 inflammasome. Chemosphere 2018, 190, 272–279. [Google Scholar] [CrossRef]
- Guan, H.; Ma, W.; Wu, Q.; Cai, J.; Zhang, Z. Exploring the Toxic Effects of ZEA on IPEC-J2 Cells from the Inflammatory Response and Apoptosis. Animals 2023, 13, 2731. [Google Scholar] [CrossRef]
- Lee, P.Y.; Liu, C.C.; Wang, S.C.; Chen, K.Y.; Lin, T.C.; Liu, P.L.; Chiu, C.C.; Chen, I.C.; Lai, Y.H.; Cheng, W.C.; et al. Mycotoxin Zearalenone Attenuates Innate Immune Responses and Suppresses NLRP3 Inflammasome Activation in LPS-Activated Macrophages. Toxins 2021, 13, 593. [Google Scholar] [CrossRef]
- Li, S.J.; Zhang, G.; Xue, B.; Ding, Q.; Han, L.; Huang, J.C.; Wu, F.; Li, C.; Yang, C. Toxicity and detoxification of T-2 toxin in poultry. Food Chem. Toxicol. 2022, 169, 113392. [Google Scholar] [CrossRef]
- Zhang, S.; Song, W.; Hua, Z.; Du, J.; Lucena, R.B.; Wang, X.; Zhang, C.; Yang, X. Overview of T-2 Toxin Enterotoxicity: From Toxic Mechanisms and Detoxification to Future Perspectives. J. Agric. Food Chem. 2024, 72, 3314–3324. [Google Scholar] [CrossRef]
- Kankkunen, P.; Valimaki, E.; Rintahaka, J.; Palomaki, J.; Nyman, T.; Alenius, H.; Wolff, H.; Matikainen, S. Trichothecene mycotoxins activate NLRP3 inflammasome through a P2X7 receptor and Src tyrosine kinase dependent pathway. Hum. Immunol. 2014, 75, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, P.; Cui, Y.; Song, M.; Zhang, X.; Zhang, C.; Jiang, Y.; Li, Y. T-2 Toxin Caused Mice Testicular Inflammation Injury via ROS-Mediated NLRP3 Inflammasome Activation. J. Agric. Food Chem. 2022, 70, 14043–14051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, B.; Huo, S.; Du, J.; Zhang, J.; Song, M.; Cui, Y.; Li, Y. T-2 Toxin Induces Kidney Fibrosis via the mtROS-NLRP3-Wnt/beta-Catenin Axis. J. Agric. Food Chem. 2022, 70, 13765–13777. [Google Scholar] [CrossRef] [PubMed]
- Mwabulili, F.; Xie, Y.; Li, Q.; Sun, S.; Yang, Y.; Ma, W. Research progress of ochratoxin a bio-detoxification. Toxicon 2023, 222, 107005. [Google Scholar] [CrossRef] [PubMed]
- Khoi, C.S.; Chen, J.H.; Lin, T.Y.; Chiang, C.K.; Hung, K.Y. Ochratoxin A-Induced Nephrotoxicity: Up-to-Date Evidence. Int. J. Mol. Sci. 2021, 22, 1237. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Liu, K.; Liu, S.; Yue, D.; Zhang, P.; Mao, X.; He, W.; Huang, K.; Chen, X. Taurine alleviates ochratoxin A-induced pyroptosis in PK-15 cells by inhibiting oxidative stress. J. Biochem. Mol. Toxicol. 2023, 37, e23249. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, S.; Lan, H.; Zheng, X. Ochratoxin A (OTA) causes intestinal aging damage through the NLRP3 signaling pathway mediated by calcium overload and oxidative stress. Env. Sci. Pollut. Res. Int. 2024, 31, 27864–27882. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wei, Z.; Wang, Y.; Long, M.; Wu, W.; Kuca, K. Fumonisin B(1): Mechanisms of toxicity and biological detoxification progress in animals. Food Chem. Toxicol. 2021, 149, 111977. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wen, J.; Tang, Y.; Shi, J.; Mu, G.; Yan, R.; Cai, J.; Long, M. Research Progress on Fumonisin B1 Contamination and Toxicity: A Review. Molecules 2021, 26, 5238. [Google Scholar] [CrossRef]
- Li, Q.; Dong, Z.; Lian, W.; Cui, J.; Wang, J.; Shen, H.; Liu, W.; Yang, J.; Zhang, X.; Cui, H. Ochratoxin A causes mitochondrial dysfunction, apoptotic and autophagic cell death and also induces mitochondrial biogenesis in human gastric epithelium cells. Arch. Toxicol. 2019, 93, 1141–1155. [Google Scholar] [CrossRef]
- Ruan, D.; Wang, W.C.; Lin, C.X.; Fouad, A.M.; Chen, W.; Xia, W.G.; Wang, S.; Luo, X.; Zhang, W.H.; Yan, S.J.; et al. Effects of curcumin on performance, antioxidation, intestinal barrier and mitochondrial function in ducks fed corn contaminated with ochratoxin A. Animal 2019, 13, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, Z.; Yue, D.; Qu, J.; Zhang, P.; Zhang, S.; Huang, K.; Zou, Y.; Wang, C.; Chen, X. Mild endoplasmic reticulum stress alleviates FB1-triggered intestinal pyroptosis via the Sec62-PERK pathway. Cell Biol. Toxicol. 2024, 40, 33. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Liu, S.; Ge, L.; Du, H.; Yue, D.; Hou, L.; Huang, K.; Chen, X. mTOR-Mediated Autophagy Regulates Fumonisin B(1)-Induced Intestinal Inflammation via Pyroptosis In Vivo and In Vitro. J. Agric. Food Chem. 2022, 70, 9187–9200. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Wang, S.; Jiang, L.; Jiao, Y.; Sun, X.; Li, J.; Yang, L.; Hou, Y.; Wang, N.; Yao, X.; et al. Patulin induces pyroptosis through the autophagic-inflammasomal pathway in liver. Food Chem. Toxicol. 2021, 147, 111867. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, Q.; He, Y.; Chen, S.; Li, T. Mycophenolic acid enhanced lipopolysaccharide-induced interleukin-18 release in THP-1 cells via activation of the NLRP3 inflammasome. Immunopharmacol. Immunotoxicol. 2019, 41, 521–526. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Pharmacological Inhibition of the NLRP3 Inflammasome: Structure, Molecular Activation, and Inhibitor-NLRP3 Interaction. Pharmacol. Rev. 2023, 75, 487–520. [Google Scholar] [CrossRef] [PubMed]
- Coll, R.C.; Hill, J.R.; Day, C.J.; Zamoshnikova, A.; Boucher, D.; Massey, N.L.; Chitty, J.L.; Fraser, J.A.; Jennings, M.P.; Robertson, A.A.B.; et al. MCC950 directly targets the NLRP3 ATP-hydrolysis motif for inflammasome inhibition. Nat. Chem. Biol. 2019, 15, 556–559. [Google Scholar] [CrossRef]
- Li, H.; Guan, Y.; Liang, B.; Ding, P.; Hou, X.; Wei, W.; Ma, Y. Therapeutic potential of MCC950, a specific inhibitor of NLRP3 inflammasome. Eur. J. Pharmacol. 2022, 928, 175091. [Google Scholar] [CrossRef]
- Sun, Y.; Su, J.; Liu, Z.; Liu, D.; Gan, F.; Chen, X.; Huang, K. Aflatoxin B(1) Promotes Influenza Replication and Increases Virus Related Lung Damage via Activation of TLR4 Signaling. Front. Immunol. 2018, 9, 2297. [Google Scholar] [CrossRef]
- Hou, L.; Gan, F.; Zhou, X.; Zhou, Y.; Qian, G.; Liu, Z.; Huang, K. Immunotoxicity of ochratoxin A and aflatoxin B1 in combination is associated with the nuclear factor kappa B signaling pathway in 3D4/21 cells. Chemosphere 2018, 199, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, R.; Chang, Q.; Dong, Z.; Yang, H.; Xu, C. Lactobacillus bulgaricus or Lactobacillus rhamnosus Suppresses NF-kappaB Signaling Pathway and Protects against AFB(1)-Induced Hepatitis: A Novel Potential Preventive Strategy for Aflatoxicosis? Toxins 2019, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Liu, M.; Zhou, X.; Wang, M.; Cao, K.; Jin, S.; Shan, A.; Feng, X. Curcumin mitigates aflatoxin B1-induced liver injury via regulating the NLRP3 inflammasome and Nrf2 signaling pathway. Food Chem. Toxicol. 2022, 161, 112823. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Wang, Y.; Rajput, S.A.; Shaukat, A.; Yang, P.; Farooq, M.Z.; Cheng, Q.; Ali, M.; Mi, X.; An, Y.; et al. Ameliorative Effects of Luteolin and Activated Charcoal on Growth Performance, Immunity Function, and Antioxidant Capacity in Broiler Chickens Exposed to Deoxynivalenol. Toxins 2023, 15, 478. [Google Scholar] [CrossRef]
- Rajput, S.A.; Shaukat, A.; Wu, K.; Rajput, I.R.; Baloch, D.M.; Akhtar, R.W.; Raza, M.A.; Najda, A.; Rafal, P.; Albrakati, A.; et al. Luteolin Alleviates AflatoxinB(1)-Induced Apoptosis and Oxidative Stress in the Liver of Mice through Activation of Nrf2 Signaling Pathway. Antioxidants 2021, 10, 1268. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.S.A.; Abadi, A.M.; El-Kott, A.F.; Mohamed, G.; Alrashdi, B.M.; Eid, R.A.; Salem, E.T. Protective efficacy of luteolin against aflatoxinB1-induced toxicity, oxidative damage, and apoptosis in the rat liver. Env. Sci. Pollut. Res. Int. 2023, 30, 52358–52368. [Google Scholar] [CrossRef] [PubMed]
- Cariddi, L.N.; Sabini, M.C.; Escobar, F.M.; Montironi, I.; Manas, F.; Iglesias, D.; Comini, L.R.; Sabini, L.I.; Dalcero, A.M. Polyphenols as possible bioprotectors against cytotoxicity and DNA damage induced by ochratoxin A. Env. Toxicol. Pharmacol. 2015, 39, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Wohlert, A.; Palkovicsne Pezsa, N.; Moritz, A.V.; Jerzsele, A.; Farkas, O.; Paszti-Gere, E. Luteolin and Chrysin Could Prevent E. coli Lipopolysaccharide-Ochratoxin A Combination-Caused Inflammation and Oxidative Stress in In Vitro Porcine Intestinal Model. Animals 2022, 12, 2747. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Cheng, C.; Li, X.; Zhou, S.; Hua, J.; Huang, J.; Li, Y.; Yang, K.; Zhang, P.; Zhang, Y.; et al. Luteolin alleviates ochratoxin A induced oxidative stress by regulating Nrf2 and HIF-1alpha pathways in NRK-52E rat kidney cells. Food Chem. Toxicol. 2020, 141, 111436. [Google Scholar] [CrossRef]
- Wen, D.; Han, W.; Chen, Q.; Qi, G.; Gao, M.; Guo, P.; Liu, Y.; Wu, Z.; Fu, S.; Lu, Q.; et al. Integrating network pharmacology and experimental validation to explore the mechanisms of luteolin in alleviating fumonisin B1-induced intestinal inflammatory injury. Toxicon 2024, 237, 107531. [Google Scholar] [CrossRef]
- Tsilioni, I.; Theoharides, T.C. Ochratoxin A stimulates release of IL-1beta, IL-18 and CXCL8 from cultured human microglia. Toxicology 2024, 502, 153738. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, I.; Murn, M.; Betbeder, A.M.; Guilcher, J.; Creppy, E.E. Effect of piroxicam on the nephrotoxicity induced by ochratoxin A in rats. Toxicology 1995, 95, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Fricke, R.F.; Jorge, J. Beneficial effect of dexamethasone in decreasing the lethality of acute T-2 toxicosis. Gen. Pharmacol. 1991, 22, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Y.; Zhang, X.; Zhang, K.; Chen, F.; Fan, J.; Wang, X.; Yang, X. Maintaining the Mitochondrial Quality Control System Was a Key Event of Tanshinone IIA against Deoxynivalenol-Induced Intestinal Toxicity. Antioxidants 2024, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Biasizzo, M.; Kopitar-Jerala, N. Interplay Between NLRP3 Inflammasome and Autophagy. Front. Immunol. 2020, 11, 591803. [Google Scholar] [CrossRef] [PubMed]
- Nurmi, K.; Kareinen, I.; Virkanen, J.; Rajamaki, K.; Kouri, V.P.; Vaali, K.; Levonen, A.L.; Fyhrquist, N.; Matikainen, S.; Kovanen, P.T.; et al. Hemin and Cobalt Protoporphyrin Inhibit NLRP3 Inflammasome Activation by Enhancing Autophagy: A Novel Mechanism of Inflammasome Regulation. J. Innate Immun. 2017, 9, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Spalinger, M.R.; Lang, S.; Gottier, C.; Dai, X.; Rawlings, D.J.; Chan, A.C.; Rogler, G.; Scharl, M. PTPN22 regulates NLRP3-mediated IL1B secretion in an autophagy-dependent manner. Autophagy 2017, 13, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Jiang, C.; Chen, H.; Chai, Y. Rapamycin Attenuates High Glucose-Induced Inflammation Through Modulation of mTOR/NF-kappaB Pathways in Macrophages. Front. Pharmacol. 2019, 10, 1292. [Google Scholar] [CrossRef]
- Zhang, X.; Du, J.; Li, B.; Huo, S.; Zhang, J.; Cui, Y.; Song, M.; Shao, B.; Li, Y. PINK1/Parkin-mediated mitophagy mitigates T-2 toxin-induced nephrotoxicity. Food Chem. Toxicol. 2022, 164, 113078. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Li, H.; Ge, L.; Liu, S.; Hou, L.; Yue, D.; Du, H.; Pan, C.; Gan, F.; Liu, Y.; et al. Selenomethionine alleviated Ochratoxin A induced pyroptosis and renal fibrotic factors expressions in MDCK cells. J. Biochem. Mol. Toxicol. 2022, 36, e22933. [Google Scholar] [CrossRef]
- Li, Y.; Chen, S.; Yu, Z.; Yao, J.; Jia, Y.; Liao, C.; Chen, J.; Wei, Y.; Guo, R.; He, L.; et al. A Novel Bacillus Velezensis for Efficient Degradation of Zearalenone. Foods 2024, 13, 530. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Guo, J.; Jia, Y.; Liao, C.; He, L.; Li, J.; Wei, Y.; Chen, S.; Chen, J.; Shang, K.; et al. A Bacillus subtilis Strain ZJ20 with AFB1 Detoxification Ability: A Comprehensive Analysis. Biology 2023, 12, 1195. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Ding, K.; Liu, S.; Li, Y.; Li, W.; Li, Y.; Cao, P.; Liu, Y.; Sun, E. Screening and identification of a Bacillus cereus strain able to degradate deoxynivalenol. Food Sci. 2016, 37, 5. [Google Scholar]

{kind=link}
| Inhibitors | Mycotoxins | Inhibitory Mechanism | Events | Ref. |
|---|---|---|---|---|
| MCC950 | OTA, T-2 toxin, FB1, and DON | Binds to Walker B motif of NATCH domain to inhibit ATPase activity and close active conformation. | ↓: NLRP3, pro-caspase-1, caspase-1, GSDMD, ASC, IL-18, pro-IL-1β, IL-1β, TNF-α, IL-6, NLRP3 inflammasome, and pyroptosis. | [4,48,54,58,92,93,103] |
| BAY 11-7082 | DON, AFB1, OTA | Binds with NATCH and leucine-rich repeat domain and inhibits ATPase. | ↓: NLRP3, pro-IL-1β, caspase-1, NF-κB signaling pathway, inflammatory responses, and cytokines. | [49,110,111,112] |
| Curcumin | AFB1 | Prevents K+ efflux, inhibits microtubule-driven recruitment of ASC on mitochondria to NLRP3 on the endoplasmic reticulum. | ↓: NLRP3 inflammasome, NLRP3/caspase-1 signaling pathways, TLR4/NF-κB signaling pathway, JAK2/NLRP3 signaling pathway, OS, inflammatory response, ITPR2, caspase-12/caspase-3 pathway, intestinal injury, fibrosis, and pyroptosis; ↑: Nrf2-ARE signaling pathway. | [66,74,77,79,113] |
| Luteolin | DON, AFB1, FB1, and OTA | Inhibits NLRP3 inflammasome activation by disrupting the interaction between NLRP3 and ASC. | ↓: apoptosis, OS, NF-κB, ERK signaling pathways, IL-6, IL-1β, extracellular H2O2, intracellular ROS, DNA damage, toxicity, and inflammatory injury; ↑: antioxidant enzymes, Nrf2 and HIF-1α pathways. | [114,115,116,117,118,119,120] |
| Piroxicam | OTA | Suppresses NLRP3 activation via reversible inhibition of volume-regulated anion (chloride) channels and the NF-κB pathway. | ↓: nephrotoxicity. | [122] |
| Tanshinone IIA | DON | Inhibits mitochondrial ROS release. | ↓: NLRP3, caspase-1, GSDMD, IL-1β, IL-18, and cell injury; improved mitochondrial function via mitochondrial quality control. | [47,124] |
| Licochalcone A | AFB1 | Binds to NEK7 and disrupts NEK7-NLRP3 interaction. | ↓: TLR4-NF-κB/MAPK, NLRP3/caspase-1/GSDMD signaling pathway, oxidative insults, inflammation, pyroptosis, and hepatotoxicity. | [65] |
| Rapamycin | FB1 | Downregulates NF-κB signaling pathway. | ↓: intestinal inflammatory injury and pyroptosis-related genes. | [103] |
| MitoQ | T-2 toxin | Reduces mitochondrial DNA damage. | ↓: mtROS, NLRP3-inflammasome, W/β signaling, structural and functional damage, and fibrosis | [93] |
| Bacillus licheniformis | AFB1 | Inhibits the expression of the NLRP3 inflammasome. | ↓: gut-testis axis damage and NLRP3-mediated NLRs signaling. | [78] |
| Dimethyl fumarate | AFB1 | Downregulates NF-κB signaling pathway. | ↓: inflammatory cytokines, pyroptosis, and neurotoxicity. | [80] |
| Melatonin | AFB1 | Inhibits the expression of the NLRP3 inflammasome. | ↓: NLRP3, ASC, caspase-1 p20, IL-1βp17, and myocardial toxicity. | [81] |
| Chlorogenic acid | DON | Inhibits the expression of the NLRP3 inflammasome. | ↓: OS, inflammation, apoptosis, and MAPK/NF-κB/NLRP3 pathway; ↑: Nrf2/HO-1 pathway. | [54] |
| Microalgal astaxanthin | DON | Modulates the mtROS/NF-κB-dependent NLRP3 inflammasome. | ↓: mtROS-NF-κB-dependent NLRP3 inflammasome and pyroptosis. | [53] |
| Taurine | OTA | Inhibits the production of ROS. | ↓: pyroptosis and ROS; ↑: antioxidant enzymes. | [96] |
| Selenomethionine | OTA | Inhibits the expression of the NLRP3 inflammasome. | ↓: NLRP3 inflammasome, NLRP3-caspase-1-dependent pyroptosis, and cytotoxicity. | [130] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.; Xu, F.; Yu, Z.; Ding, K.; Jia, Y. The Novel Role of the NLRP3 Inflammasome in Mycotoxin-Induced Toxicological Mechanisms. Vet. Sci. 2024, 11, 291. https://doi.org/10.3390/vetsci11070291
Liao C, Xu F, Yu Z, Ding K, Jia Y. The Novel Role of the NLRP3 Inflammasome in Mycotoxin-Induced Toxicological Mechanisms. Veterinary Sciences. 2024; 11(7):291. https://doi.org/10.3390/vetsci11070291
Chicago/Turabian StyleLiao, Chengshui, Fengru Xu, Zuhua Yu, Ke Ding, and Yanyan Jia. 2024. "The Novel Role of the NLRP3 Inflammasome in Mycotoxin-Induced Toxicological Mechanisms" Veterinary Sciences 11, no. 7: 291. https://doi.org/10.3390/vetsci11070291
APA StyleLiao, C., Xu, F., Yu, Z., Ding, K., & Jia, Y. (2024). The Novel Role of the NLRP3 Inflammasome in Mycotoxin-Induced Toxicological Mechanisms. Veterinary Sciences, 11(7), 291. https://doi.org/10.3390/vetsci11070291