
Putative mRNA Biomarkers for the Eradication of Infection in an Equine Experimental Model of Septic Arthritis

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. RNA Extraction and Library Preparation
2.2. Sequencing
2.3. Data Processing
2.4. Differential Gene Expression Analysis
3. Results
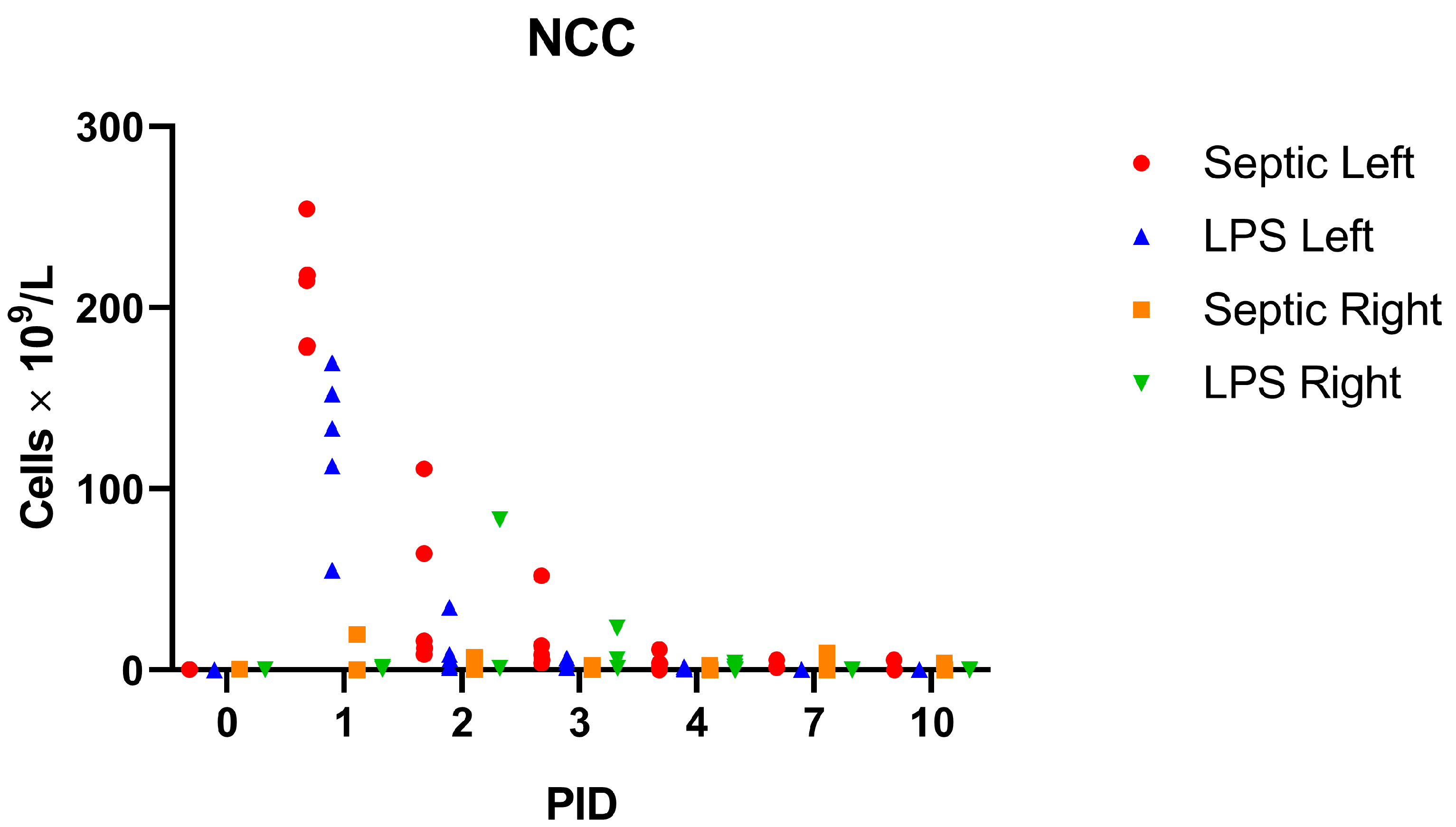
3.1. Experimental Model
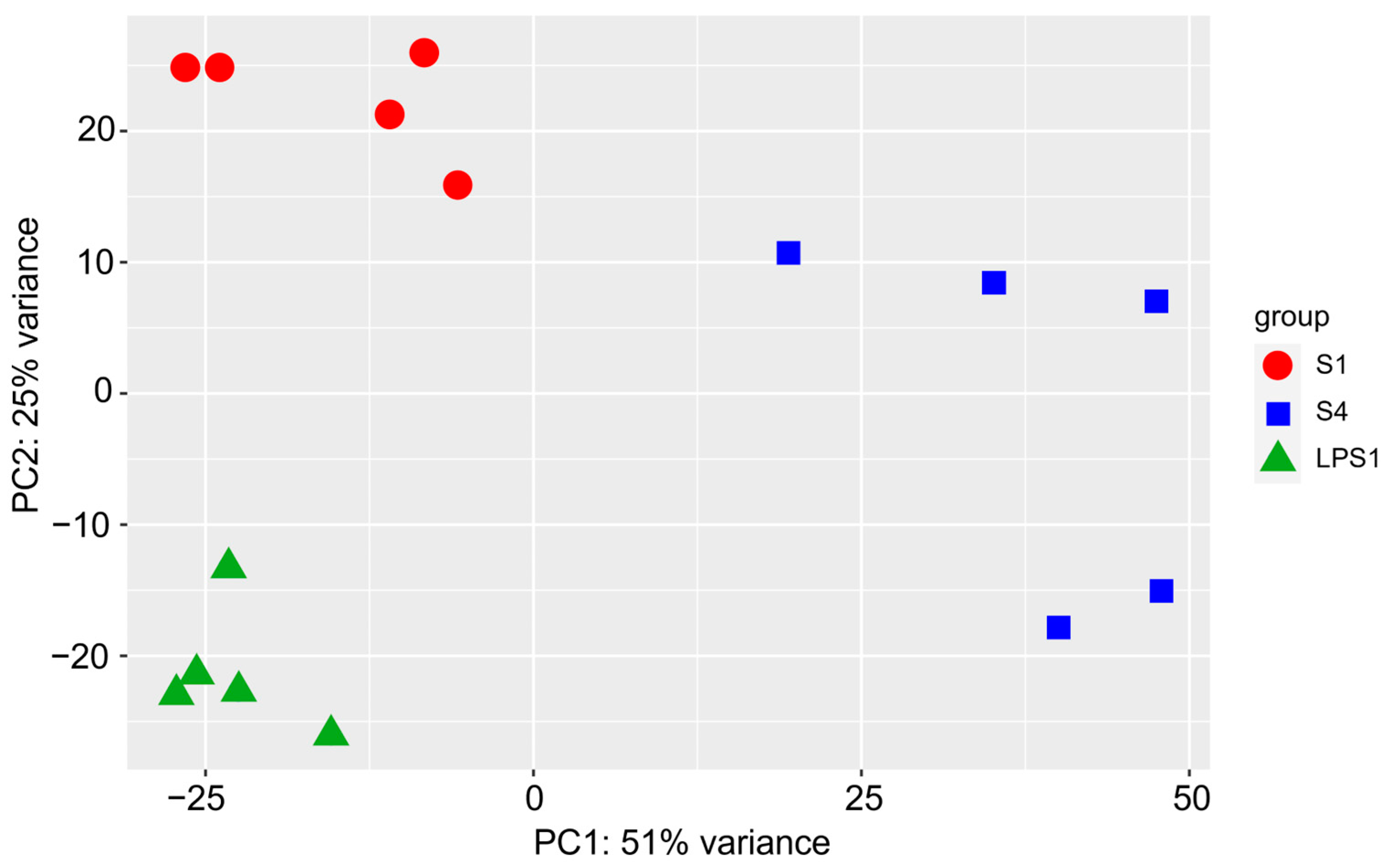
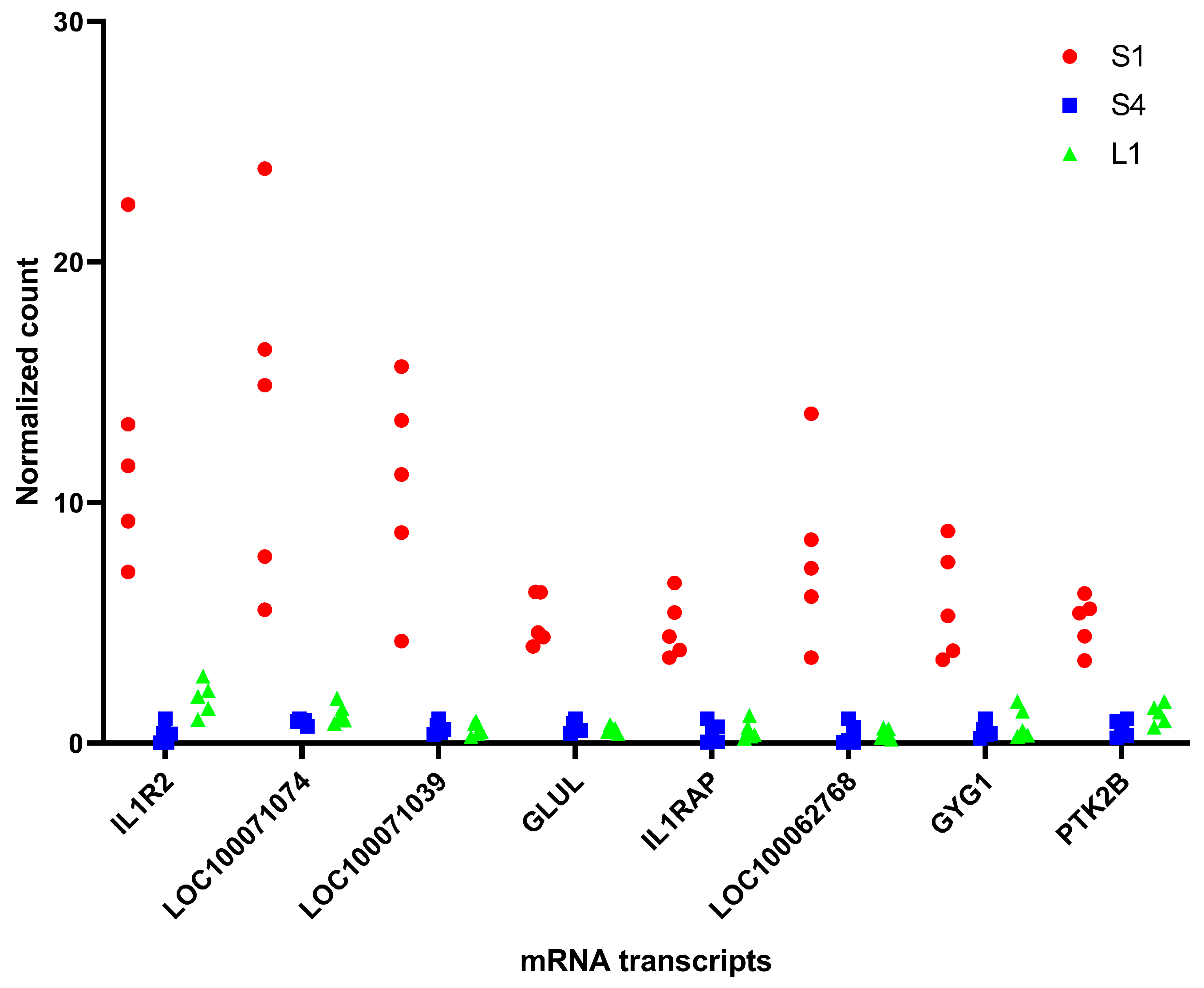
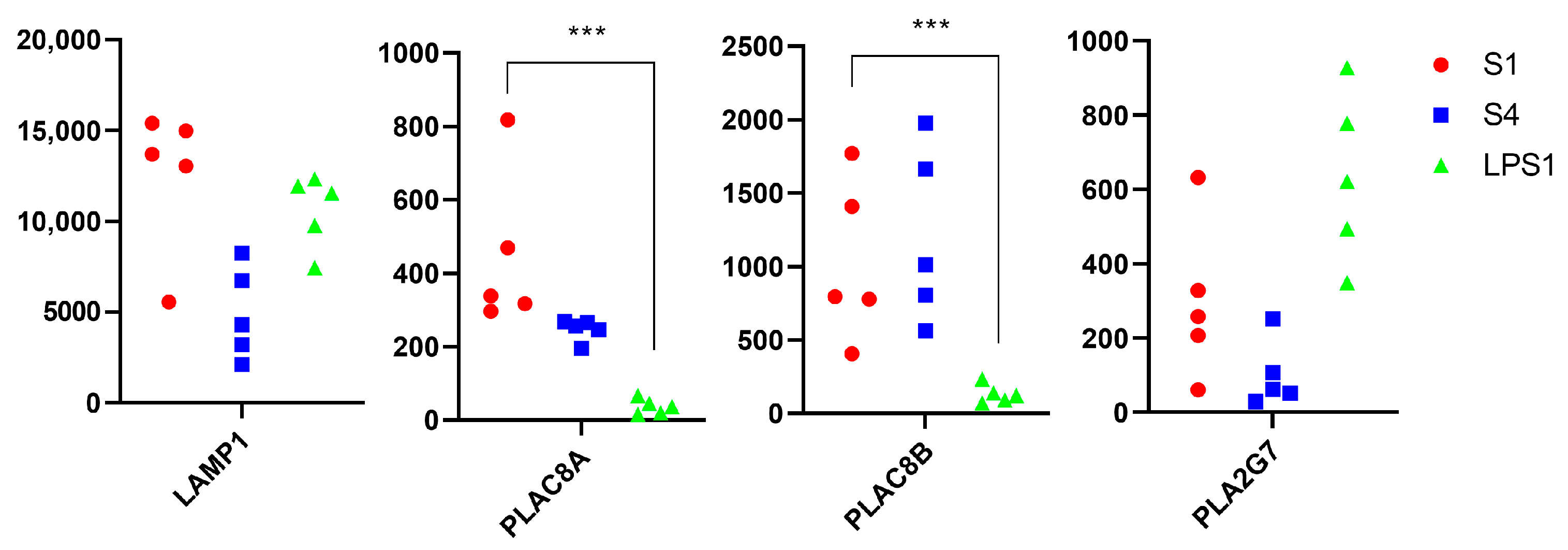
3.2. Transcriptomics Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koziy, R.V.; Yoshimura, S.; Dickinson, R.; Rybicka, J.M.; Moshynskyy, I.; Ngeleka, M.; Bracamonte, J.L.; Simko, E. Use of Standard Diagnostic Techniques to Determine Eradication of Infection in Experimental Equine Septic Arthritis. Can. J. Vet. Res. 2019, 83, 24–33. [Google Scholar] [PubMed]
- Schneider, R.K.; Bramlage, L.R.; Moore, R.M.; Mecklenburg, L.M.; Kohn, C.W.; Gabel, A.A. A Retrospective Study of 192 Horses Affected with Septic Arthritis/Tenosynovitis. Equine Vet. J. 1992, 24, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.H.; Mair, T.S.; Smith, L.J.; Perkins, J.D. Bacterial Culture of Septic Synovial Structures of Horses: Does a Positive Bacterial Culture Influence Prognosis? Equine Vet. J. 2010, 42, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Motta, R.G.; Martins, L.S.A.; Motta, I.G.; Guerra, S.T.; de Paula, C.L.; Bolanos, C.A.D.; da Silva, R.C.; Ribeiro, M.G. Multidrug Resistant Bacteria Isolated from Septic Arthritis in Horses. Pesqui. Veterinária Bras. Braz. J. Vet. Res. 2017, 37, 325–330. [Google Scholar] [CrossRef]
- Califf, R.M. Biomarker Definitions and Their Applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Fietz, S.A. Biomarkers in Equine Synovial Fluid and Serum for the Diagnosis of Joint Diseases. Ph.D. Dissertation, Faculty of Veterinary Medicine, Freie Universität Berlin, Berlin, Germany, 2009. [Google Scholar] [CrossRef]
- Rifai, N.; Gillette, M.A.; Carr, S.A. Protein Biomarker Discovery and Validation: The Long and Uncertain Path to Clinical Utility. Nat. Biotechnol. 2006, 24, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.R.; Phelan, M.M.; Foddy, L.; Clegg, P.D.; Peffers, M.J. Ex Vivo Equine Cartilage Explant Osteoarthritis Model: A Metabolomics and Proteomics Study. J. Proteome Res. 2020, 19, 3652–3667. [Google Scholar] [CrossRef]
- Yoshimura, S.; Koziy, R.V.; Dickinson, R.; Moshynskyy, I.; McKenzie, J.A.; Simko, E.; Bracamonte, J.L. Use of Serum Amyloid A in Serum and Synovial Fluid to Detect Eradication of Infection in Experimental Septic Arthritis in Horses. Can. J. Vet. Res. 2020, 84, 198–204. [Google Scholar]
- Koziy, R.V.; Bracamonte, J.L.; Yoshimura, S.; Chumala, P.; Simko, E.; Katselis, G.S. Discovery Proteomics for the Detection of Putative Markers for Eradication of Infection in an Experimental Model of Equine Septic Arthritis Using LC-MS/MS. J. Proteomics 2022, 261, 104571. [Google Scholar] [CrossRef]
- McHugh, L.; Seldon, T.A.; Brandon, R.A.; Kirk, J.T.; Rapisarda, A.; Sutherland, A.J.; Presneill, J.J.; Venter, D.J.; Lipman, J.; Thomas, M.R.; et al. A Molecular Host Response Assay to Discriminate between Sepsis and Infection-Negative Systemic Inflammation in Critically Ill Patients: Discovery andvValidation in Independent Cohorts. PLoS Med. 2015, 12, e1001916. [Google Scholar] [CrossRef]
- Miller, R.R.; Lopansri, B.K.; Burke, J.P.; Levy, M.; Opal, S.; Rothman, R.E.; D’Alessio, F.R.; Sidhaye, V.K.; Aggarwal, N.R.; Balk, R.; et al. Validation of a Host Response Assay, SeptiCyte LAB, for Discriminating Sepsis from Systemic Inflammatory Response Syndrome in the ICU. Am. J. Respir. Crit. Care Med. N. Y. 2018, 198, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.N.; Therneau, T.M.; Zhang, Y.; Poland, G.A.; Kocher, J.-P. Calculating Sample Size Estimates for RNA Sequencing Data. J. Comput. Biol. 2013, 20, 970–978. [Google Scholar] [CrossRef]
- Karunarathna, R.; Popowich, S.; Wawryk, M.; Chow-Lockerbie, B.; Ahmed, K.A.; Yu, C.; Liu, M.; Goonewardene, K.; Gunawardana, T.; Kurukulasuriya, S.; et al. Increased Incidence of Enterococcal Infection in Nonviable Broiler Chicken Embryos in Western Canadian Hatcheries as Detected by Matrix-Assisted Laser Desorption/Ionization-Time-of-Flight Mass Spectrometry. Avian Dis. 2017, 61, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The R Package Rsubread Is Easier, Faster, Cheaper and Better for Alignment and Quantification of RNA Sequencing Reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Steel, C.M. Equine Synovial Fluid Analysis. Vet. Clin. N. Am. Equine Pract. 2008, 24, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Mahaffey, E.A. Chapter 10—Synovial Fluid. In Diagnostic Cytology and Hematology of the Horse, 2nd ed.; Cowell, R.L., Tyler, R.D., Eds.; Mosby: St. Louis, MO, USA, 2002; pp. 163–170. ISBN 978-0-323-01317-8. [Google Scholar]
- Yu, J.; Zhang, J.; Shi, M.; Ding, H.; Ma, L.; Zhang, H.; Liu, J. Maintenance of Glutamine Synthetase Expression Alleviates Endotoxin-Induced Sepsis via Alpha-Ketoglutarate-Mediated Demethylation. FASEB J. 2022, 36, e22281. [Google Scholar] [CrossRef]
- Bianchi, M.G.; Chiu, M.; Taurino, G.; Bergamaschi, E.; Cubadda, F.; Macaluso, G.M.; Bussolati, O. The TLR4/NFκB-Dependent Inflammatory Response Activated by LPS Is Inhibited in Human Macrophages Pre-Exposed to Amorphous Silica Nanoparticles. Nanomaterials 2022, 12, 2307. [Google Scholar] [CrossRef]
- Colotta, F.; Dower, S.K.; Sims, J.E.; Mantovani, A. The Type II ‘Decoy’ Receptor: A Novel Regulatory Pathway for Interleukin 1. Immunol. Today 1994, 15, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Jiang, Y.; Gao, M.; Wang, W.; Wang, N.; Wang, K.; Zhang, H.; Chen, G.; Liu, K.; Liu, M.; et al. Interleukin-1 Receptor 2: A New Biomarker for Sepsis Diagnosis and Gram-Negative/Gram-Positive Bacterial Differentiation. Shock 2017, 47, 119. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.L.; Dickinson, P.; Forster, T.; Craigon, M.; Ross, A.; Khondoker, M.R.; France, R.; Ivens, A.; Lynn, D.J.; Orme, J.; et al. Identification of a Human Neonatal Immune-Metabolic Network Associated with Bacterial Infection. Nat. Commun. 2014, 5, 4649. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Strunk, T.; Lee, A.H.; Gill, E.E.; Falsafi, R.; Woodman, T.; Hibbert, J.; Hancock, R.E.W.; Currie, A. Whole Blood Transcriptional Responses of Very Preterm Infants during Late-Onset Sepsis. PLoS ONE 2020, 15, e0233841. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gao, X.; Li, S.; Cao, Z. Recruitment of IRAK to the Interleukin 1 Receptor Complex Requires Interleukin 1 Receptor Accessory Protein. Proc. Natl. Acad. Sci. USA 1997, 94, 12829–12832. [Google Scholar] [CrossRef] [PubMed]
- Kulohoma, B.W.; Marriage, F.; Vasieva, O.; Mankhambo, L.; Nguyen, K.; Molyneux, M.E.; Molyneux, E.M.; Day, P.J.R.; Carrol, E.D. Peripheral Blood RNA Gene Expression in Children with Pneumococcal Meningitis: A Prospective Case–Control Study. BMJ Paediatr. Open 2017, 1, e000092. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | UniProt Accession Number | Protein Name |
|---|---|---|---|
| IL1R2 | ENSECAG00000000288 | F7DK71 | Interleukin-1 receptor type 2 |
| LOC100071074 | ENSECAG00000039739 | F6QB61 | Peptidoglycan-recognition protein |
| LOC100071039 | ENSECAG00000032321 | F7DQG1 | Peptidoglycan-recognition protein |
| GLUL | ENSECAG00000015865 | F6TAZ0 | Glutamine synthetase |
| IL1RAP | ENSECAG00000005083 | F7AWQ9 | Interleukin 1 receptor accessory protein |
| LOC100062768 | ENSECAG00000006255 | F7ADT4 | G-protein coupled receptors family 1 profile domain-containing protein |
| GYG1 | ENSECAG00000023192 | F6U6H7 | Glycogenin 1 |
| PTK2B | ENSECAG00000018924 | F7CGD4 | Non-specific protein-tyrosine kinase |
| Gene Name | Septic Arthritis PID1 | Eradicated Septic Arthritis PID4 | Non-Septic Synovitis PID1 | |
|---|---|---|---|---|
| IL1RAP | Mean | 614,082 | 623,448 | 1,358,139 |
| SD | 408,114 | 241,337 | 1,041,416 | |
| GYG1 | Mean | 0 | 0 | 30,143 |
| SD | 0 | 0 | 45,269 |
| Protein Name | Gene ID | Gene Name | p-Value Septic Arthritis PID1 vs. Eradicated Septic Arthritis PID4 | Fold Change | p-Value Septic Arthritis PID1 vs. Non-Septic Synovitis PID1 | Fold Change |
|---|---|---|---|---|---|---|
| E3 ubiquitin-protein ligase TRIM9 | ENSECAG00000003278 | TRIM9 | 0.082 | −4.1 | 0.859 | 1.3 |
| DCC-interacting protein 13-alpha | ENSECAG00000017473 | APPL1 | 0.130 | −1.3 | 0.031 | −1.5 |
| Ubiquitin-like-conjugating enzyme ATG3 | ENSECAG00000011954 | ATG3 | 0.586 | −1.1 | 0.849 | 1.1 |
| Aldehyde dehydrogenase family 16 member A1 | ENSECAG00000023437 | ALDH16A1 | 0.145 | 1.3 | 0.638 | −1.1 |
| Serine/threonine-protein kinase 24 | ENSECAG00000007682 | STK24 | 0.007 | 1.5 | 0.604 | 1.1 |
| Protein ABHD14B | ENSECAG00000022421 | ABHD14B | 0.700 | −1.1 | 0.359 | −1.2 |
| Transforming protein RhoA | ENSECAG00000033791 | RHOA | 0.009 | 1.6 | 0.032 | 1.5 |
| Guanine nucleotide-binding protein subunit beta−2 | ENSECAG00000017221 | GNB2 | 0.000 | 2.4 | 0.000 | 1.9 |
| Ubiquitin-conjugating enzyme E2 L3 | ENSECAG00000032131 | UBE2L6 | 0.170 | 1.6 | 0.000 | 7.1 |
| N-acetyltransferase ESCO1 | ENSECAG00000016120 | ESCO1 | 0.000 | 2.8 | 0.077 | 1.6 |
| Twinfilin-2 | ENSECAG00000014047 | TWF2 | 0.068 | 1.3 | 0.003 | 1.5 |
| Nuclear transport factor 2 | ENSECAG00000021530 | NUTF2 | 0.000 | −2.7 | 0.048 | −1.8 |
| Elongation factor 1-gamma | ENSECAG00000014334 | EEF1G | 0.000 | −2.3 | 0.841 | 1.1 |
| Septin-7 (Fragment) | ENSECAG00000022693 | SEPTIN7 | 0.216 | −1.2 | 0.473 | 1.2 |
| GTP-binding nuclear protein Ran | ENSECAG00000020532 | RAN | 0.071 | −1.5 | 0.283 | 1.3 |
| Synaptic vesicle membrane protein VAT-1 homolog | ENSECAG00000016700 | VAT1 | 0.790 | 1.1 | 0.001 | −2.4 |
| Alpha-1-acid glycoprotein 2 | ENSECAG00000036760 | NA | 0.656 | −1.3 | 0.126 | −2.2 |
| BTB/POZ domain-containing protein KCTD12 | ENSECAG00000002083 | KCTD12 | 0.000 | 3.6 | 0.002 | 2.8 |
| Interleukin-1 receptor antagonist protein | ENSECAG00000027864 | IL1RN | 0.014 | 4.2 | 0.000 | 9.3 |
| Ubiquitin-like modifier-activating enzyme 1 | ENSECAG00000014002 | UBA1 | 0.000 | 2.6 | 0.000 | 2.8 |
| Heterogeneous nuclear ribonucleoprotein D0 | ENSECAG00000022692 | HNRNPDL | 0.051 | −1.3 | 0.678 | 1.1 |
| Neutrophil collagenase | ENSECAG00000023733 | MMP1/8 | 0.000 | 22.9 | 0.000 | 4.2 |
| Fermitin family homolog 3 | ENSECAG00000008107 | FERMT3 | 0.815 | 1.1 | 0.332 | 1.3 |
| Coronin-7 | ENSECAG00000024788 | CORO1A | 0.769 | 1.1 | 0.026 | 1.8 |
| Poly(rC)-binding protein 1 | ENSECAG00000024218 | PCBP4 | 0.020 | −7.0 | 0.547 | −2.0 |
| Myotrophin | ENSECAG00000024284 | MTPN | 0.927 | −1.0 | 0.229 | 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koziy, R.V.; Bracamonte, J.L.; Katselis, G.S.; Udenze, D.; Hayat, S.; Hammond, S.A.; Simko, E. Putative mRNA Biomarkers for the Eradication of Infection in an Equine Experimental Model of Septic Arthritis. Vet. Sci. 2024, 11, 299. https://doi.org/10.3390/vetsci11070299
Koziy RV, Bracamonte JL, Katselis GS, Udenze D, Hayat S, Hammond SA, Simko E. Putative mRNA Biomarkers for the Eradication of Infection in an Equine Experimental Model of Septic Arthritis. Veterinary Sciences. 2024; 11(7):299. https://doi.org/10.3390/vetsci11070299
Chicago/Turabian StyleKoziy, Roman V., José L. Bracamonte, George S. Katselis, Daniel Udenze, Shahina Hayat, S. Austin Hammond, and Elemir Simko. 2024. "Putative mRNA Biomarkers for the Eradication of Infection in an Equine Experimental Model of Septic Arthritis" Veterinary Sciences 11, no. 7: 299. https://doi.org/10.3390/vetsci11070299
APA StyleKoziy, R. V., Bracamonte, J. L., Katselis, G. S., Udenze, D., Hayat, S., Hammond, S. A., & Simko, E. (2024). Putative mRNA Biomarkers for the Eradication of Infection in an Equine Experimental Model of Septic Arthritis. Veterinary Sciences, 11(7), 299. https://doi.org/10.3390/vetsci11070299