

Reproductive Tract Microbiota of Mares
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
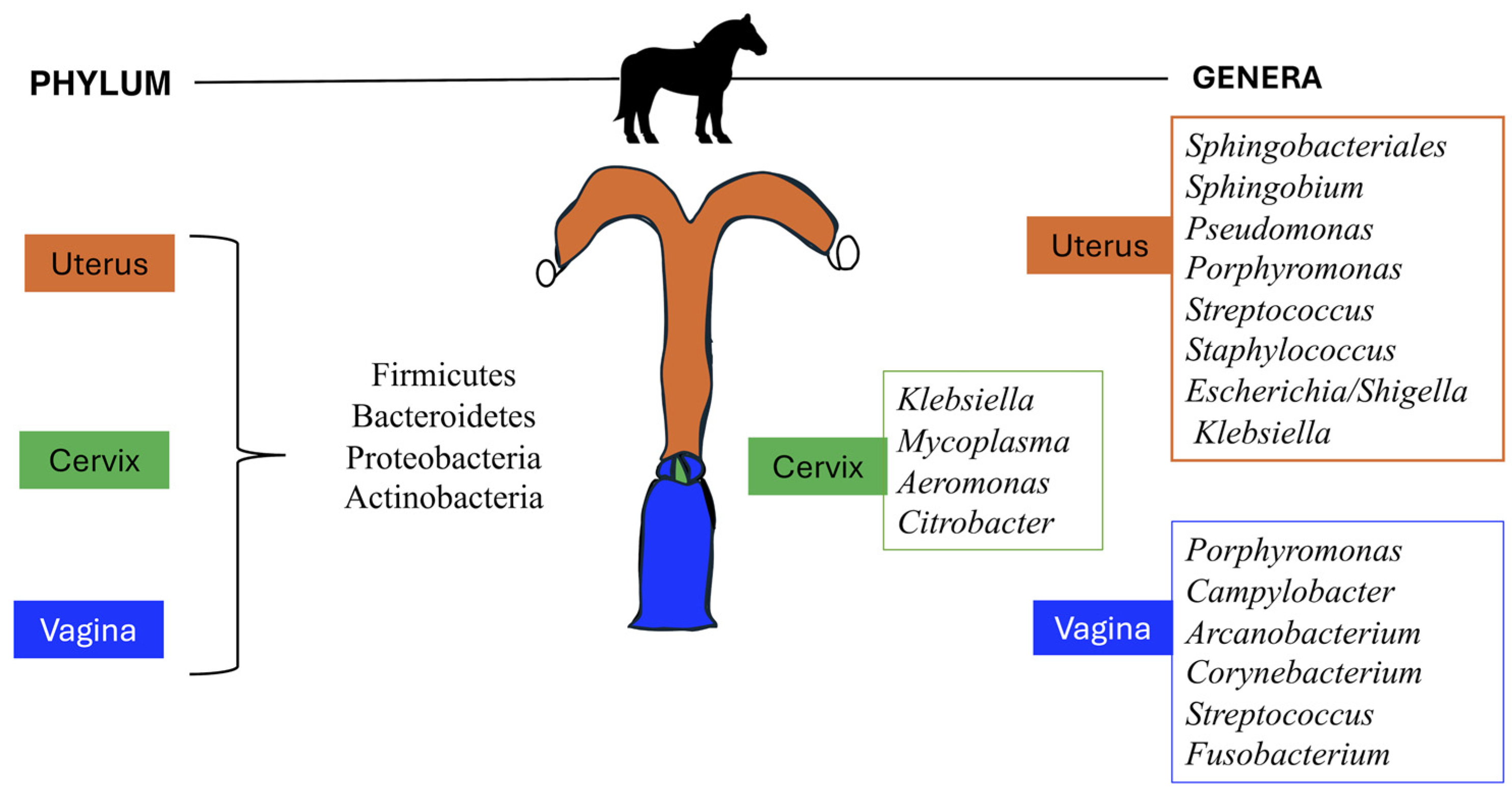
3.1. Methods and Sampling Techniques to Characterize Uterine and Vaginal Microbiota
3.2. Uterine Microbiota
3.3. Vaginal Microbiota
3.4. Factors Associated with Colonization and Establishment of the Reproductive Microbiota: Site, Diet, Parity, Stage of Estrous, and Species
3.5. The Reproductive Microbiota and Disease
3.6. The Reproductive Microbiota and Fertility
3.7. Limitations in Current Equine Reproductive Microbiome Literature
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomez, D.; Toribio, R.; Caddey, B.; Costa, M.; Vijan, S.; Dembek, K. Longitudinal Effects of Oral Administration of Antimicrobial Drugs on Fecal Microbiota of Horses. J. Vet. Intern. Med. 2023, 37, 2562–2572. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.E.; Wong, D.; MacNicol, J.; Dembek, K. The Fecal Bacterial Microbiota of Healthy and Sick Newborn Foals. J. Vet. Intern. Med. 2023, 37, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, C.; Arroyo, L.G.; MacNicol, J.L.; Renaud, D.; Weese, J.S.; Gomez, D.E. Fecal Microbiota of Horses with Colitis and Its Association with Laminitis and Survival during Hospitalization. J. Vet. Intern. Med. 2022, 36, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, C.; Arroyo, L.G.; Renaud, D.; Weese, J.S.; Gomez, D.E. Fecal Microbiota Comparison Between Healthy Teaching Horses and Client-Owned Horses. J. Equine Vet. Sci. 2022, 118, 104105. [Google Scholar] [CrossRef] [PubMed]
- Heil, B.A.; Paccamonti, D.L.; Sones, J.L. Role for the Mammalian Female Reproductive Tract Microbiome in Pregnancy Outcomes. Physiol. Genom. 2019, 51, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Mangogna, A.; Bossi, F.; Ricci, G.; Kishore, U.; Bulla, R. Uterine Immunity and Microbiota: A Shifting Paradigm. Front. Immunol. 2019, 10, 2387. [Google Scholar] [CrossRef] [PubMed]
- Liptáková, A.; Čurová, K.; Záhumenský, J.; Visnyaiová, K.; Varga, I. Microbiota of Female Genital Tract-Functional Overview of Microbial Flora from Vagina to Uterine Tubes and Placenta. Physiol. Res. 2022, 71, S21–S33. [Google Scholar] [CrossRef] [PubMed]
- Garber, A.; Hastie, P.; Murray, J.-A. Factors Influencing Equine Gut Microbiota: Current Knowledge. J. Equine Vet. Sci. 2020, 88, 102943. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.; Daley, P.; Baird, G.G.; Sturgess, S.; Frost, A.J. The Aerobic Bacterial Flora of the Reproductive Tract of the Mare. Vet. Rec. 1971, 88, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, K.; Cummings, M.R.; Sertich, P.L.; Kenney, R.M. Clinical Significance of Aerobic Bacterial Flora of the Uterus, Vagina, Vestibule, and Clitoral Fossa of Clinically Normal Mares. J. Am. Vet. Med. Assoc. 1988, 193, 72–75. [Google Scholar]
- Purswell, B.J.; Ley, W.B.; Sriranganathan, N.; Bowen, J.M. Aerobic and Anaerobic Bacterial Flora in the Postpartum Mare. J. Equine Vet. Sci. 1989, 9, 141–144. [Google Scholar] [CrossRef]
- Ferris, R.A.; McCue, P.M.; Borlee, G.I.; Glapa, K.E.; Martin, K.H.; Mangalea, M.R.; Hennet, M.L.; Wolfe, L.M.; Broeckling, C.D.; Borlee, B.R. Model of Chronic Equine Endometritis Involving a Pseudomonas Aeruginosa Biofilm. Infect. Immun. 2017, 85, e00332-17. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.R.; Skive, B.; Christoffersen, M.; Lu, K.; Nielsen, J.M.; Troedsson, M.H.T.; Bojesen, A.M. Activation of Persistent Streptococcus Equi Subspecies Zooepidemicus in Mares with Subclinical Endometritis. Vet. Microbiol. 2015, 179, 119–125. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, M.M.; Causey, R.C. Clinical and Subclinical Endometritis in the Mare: Both Threats to Fertility. Reprod. Domest. Anim. 2009, 44 (Suppl. S3), 10–22. [Google Scholar] [CrossRef] [PubMed]
- Heil, B.A.; van Heule, M.; Thompson, S.K.; Kearns, T.A.; Oberhaus, E.L.; King, G.; Daels, P.; Dini, P.; Sones, J.L. Effect of Sampling Method on Detection of the Equine Uterine Microbiome during Estrus. Vet. Sci. 2023, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.H.A.; McCue, P.M.; Aurich, C. Equine Endometritis: A Review of Challenges and New Approaches. Reproduction 2020, 160, R95–R110. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting Microbial Biofilms: Current and Prospective Therapeutic Strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Thurlow, L.R.; Hanke, M.L.; Fritz, T.; Angle, A.; Aldrich, A.; Williams, S.H.; Engebretsen, I.L.; Bayles, K.W.; Horswill, A.R.; Kielian, T. Staphylococcus Aureus Biofilms Prevent Macrophage Phagocytosis and Attenuate Inflammation in Vivo. J. Immunol. 2011, 186, 6585–6596. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.F.; O’Toole, G.A. Mechanisms of Biofilm Resistance to Antimicrobial Agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Schnobrich, M.; Atwood, K.; Barr, B.; Bradecamp, E.; Scoggin, C.; Hospital, R. Next Generation DNA Sequencing, Culture and Cytology Results in 10 Clinically Normal Mares. Clin. Theriogenology 2017, 9, 443. [Google Scholar]
- Sathe, S.; Leiken, A.; Plummer, P. Metagenomic Sequencing of the Uterine Microbial Environment during Estrus and Early Pregnancy in Mares. Clin. Theriogenology 2017, 9, 453. [Google Scholar]
- Bharti, R.; Grimm, D.G. Current Challenges and Best-Practice Protocols for Microbiome Analysis. Brief. Bioinform. 2021, 22, 178–193. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. 16S RRNA Gene Sequencing for Bacterial Identification in the Diagnostic Laboratory: Pluses, Perils, and Pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [PubMed]
- Kauter, A.; Epping, L.; Semmler, T.; Antao, E.-M.; Kannapin, D.; Stoeckle, S.D.; Gehlen, H.; Lübke-Becker, A.; Günther, S.; Wieler, L.H.; et al. The Gut Microbiome of Horses: Current Research on Equine Enteral Microbiota and Future Perspectives. Anim. Microbiome 2019, 1, 14. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Holyoak, G.R.; Premathilake, H.U.; Lyman, C.C.; Sones, J.L.; Gunn, A.; Wieneke, X.; DeSilva, U. The Healthy Equine Uterus Harbors a Distinct Core Microbiome plus a Rich and Diverse Microbiome That Varies with Geographical Location. Sci. Rep. 2022, 12, 14790. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, T.L.; Garcia, M.C.; Hurtgen, J.P.; Kenney, R.M. Comparison of Two Techniques for Obtaining Endometrial Bacteriologic Cultures in the Mare. Theriogenology 1981, 16, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Ansbacher, R.; Boyson, W.A.; Morris, J.A. Sterility of the Uterine Cavity. Am. J. Obstet. Gynecol. 1967, 99, 394–396. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed]
- Barba, M.; Martínez-Boví, R.; Quereda, J.J.; Mocé, M.L.; Plaza-Dávila, M.; Jiménez-Trigos, E.; Gómez-Martín, Á.; González-Torres, P.; Carbonetto, B.; García-Roselló, E. Vaginal Microbiota Is Stable throughout the Estrous Cycle in Arabian Maress. Animals 2020, 10, 2020. [Google Scholar] [CrossRef] [PubMed]
- Jones, E. Characterization of the Equine Microbiome during Late Gestation and the Early Postpartum Period, and at Various Times during the Estrous Cycle in Mares Being Bred with Raw or Extended Semen. Master’s Thesis, Kansas City University, Kansas City, MO, USA, 2019. [Google Scholar]
- Beckers, K.F.; Gomes, V.C.L.; Crissman, K.R.; Liu, C.-C.; Schulz, C.J.; Childers, G.W.; Sones, J.L. Metagenetic Analysis of the Pregnant Microbiome in Horses. Animals 2023, 13, 1999. [Google Scholar] [CrossRef] [PubMed]
- Holyoak, R.; Premathilake, H.; Guo, M.; DeSilva, U. The conundrum of the uterine microbiome. Clin. Theriogenology 2022, 14, 247–251. [Google Scholar] [CrossRef]
- Thomson, P.; Pareja, J.; Núñez, A.; Santibáñez, R.; Castro, R. Characterization of Microbial Communities and Predicted Metabolic Pathways in the Uterus of Healthy Mares. Open Vet. J. 2022, 12, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Malaluang, P.; Wilén, E.; Frosth, S.; Lindahl, J.; Hansson, I.; Morrell, J.M. Vaginal Bacteria in Mares and the Occurrence of Antimicrobial Resistance. Microorganisms 2022, 10, 2204. [Google Scholar] [CrossRef] [PubMed]
- Malaluang, P.; Åkerholm, T.; Nyman, G.; Lindahl, J.; Hansson, I.; Morrell, J.M. Bacteria in the Healthy Equine Vagina during the Estrous Cycle. Theriogenology 2024, 213, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Virendra, A.; Gulavane, S.U.; Ahmed, Z.A.; Reddy, R.; Chaudhari, R.J.; Gaikwad, S.M.; Shelar, R.R.; Ingole, S.D.; Thorat, V.D.; Khanam, A.; et al. Metagenomic analysis unravels novel taxonomic differences in the uterine microbiome between healthy mares and mares with endometritis. Vet. Med. Sci. 2024, 10, e1369. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.G.; Ericsson, A.C.; Poock, S.E.; Melendez, P.; Lucy, M.C. Hot Topic: 16S RRNA Gene Sequencing Reveals the Microbiome of the Virgin and Pregnant Bovine Uterus. J. Dairy. Sci. 2017, 100, 4953–4960. [Google Scholar] [CrossRef]
- Moreno, I.; Simon, C. Deciphering the Effect of Reproductive Tract Microbiota on Human Reproduction. Reprod. Med. Biol. 2019, 18, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Lyman, C.C.; Holyoak, G.R.; Meinkoth, K.; Wieneke, X.; Chillemi, K.A.; DeSilva, U. Canine Endometrial and Vaginal Microbiomes Reveal Distinct and Complex Ecosystems. PLoS ONE 2019, 14, e0210157. [Google Scholar] [CrossRef]
- Lorenzen, E.; Kudirkiene, E.; Gutman, N.; Grossi, A.B.; Agerholm, J.S.; Erneholm, K.; Skytte, C.; Dalgaard, M.D.; Bojesen, A.M. The Vaginal Microbiome Is Stable in Prepubertal and Sexually Mature Ellegaard Göttingen Minipigs throughout an Estrous Cycle. Vet. Res. 2015, 46, 125. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cheng, G.; Li, C.; Yang, J.; Li, J.; Chen, D.; Zou, W.; Jin, S.; Zhang, H.; Li, D.; et al. The Normal Vaginal and Uterine Bacterial Microbiome in Giant Pandas (Ailuropoda melanoleuca). Microbiol. Res. 2017, 199, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Miranda-CasoLuengo, R.; Lu, J.; Williams, E.J.; Miranda-CasoLuengo, A.A.; Carrington, S.D.; Evans, A.C.O.; Meijer, W.G. Delayed Differentiation of Vaginal and Uterine Microbiomes in Dairy Cows Developing Postpartum Endometritis. PLoS ONE 2019, 14, e0200974. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Chase, D.M.; Herbst-Kralovetz, M.M. Uterine Microbiota: Residents, Tourists, or Invaders? Front. Immunol. 2018, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.K.; Becher, N.; Bastholm, S.; Glavind, J.; Ramsing, M.; Kim, C.J.; Romero, R.; Jensen, J.S.; Uldbjerg, N. The Cervical Mucus Plug Inhibits, but Does Not Block, the Passage of Ascending Bacteria from the Vagina during Pregnancy. Acta Obstet. Gynecol. Scand. 2014, 93, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Kunz, G.; Leyendecker, G. Uterine Peristaltic Activity during the Menstrual Cycle: Characterization, Regulation, Function and Dysfunction. Reprod. Biomed. Online 2002, 4 (Suppl. S3), 5–9. [Google Scholar] [CrossRef] [PubMed]
- Kunz, G.; Beil, D.; Deiniger, H.; Einspanier, A.; Mall, G.; Leyendecker, G. The Uterine Peristaltic Pump. Normal and Impeded Sperm Transport within the Female Genital Tract. Adv. Exp. Med. Biol. 1997, 424, 267–277. [Google Scholar] [PubMed]
- Lehtoranta, L.; Ala-Jaakkola, R.; Laitila, A.; Maukonen, J. Healthy Vaginal Microbiota and Influence of Probiotics Across the Female Life Span. Front. Microbiol. 2022, 13, 819958. [Google Scholar] [CrossRef] [PubMed]
- Ault, T.B.; Clemmons, B.A.; Reese, S.T.; Dantas, F.G.; Franco, G.A.; Smith, T.P.L.; Edwards, J.L.; Myer, P.R.; Pohler, K.G. Bacterial Taxonomic Composition of the Postpartum Cow Uterus and Vagina Prior to Artificial Insemination1. J. Anim. Sci. 2019, 97, 4305–4313. [Google Scholar] [CrossRef] [PubMed]
- Quereda, J.J.; Barba, M.; Mocé, M.L.; Gomis, J.; Jiménez-Trigos, E.; García-Muñoz, Á.; Gómez-Martín, Á.; González-Torres, P.; Carbonetto, B.; García-Roselló, E. Vaginal Microbiota Changes During Estrous Cycle in Dairy Heifers. Front. Vet. Sci. 2020, 7, 371. [Google Scholar] [CrossRef] [PubMed]
- Messman, R.D.; Contreras-Correa, Z.E.; Paz, H.A.; Perry, G.; Lemley, C.O. Vaginal Bacterial Community Composition and Concentrations of Estradiol at the Time of Artificial Insemination in Brangus Heifers. J. Anim. Sci. 2020, 98, skaa178. [Google Scholar] [CrossRef] [PubMed]
- Neggers, Y.H.; Nansel, T.R.; Andrews, W.W.; Schwebke, J.R.; Yu, K.; Goldenberg, R.L.; Klebanoff, M.A. Dietary Intake of Selected Nutrients Affects Bacterial Vaginosis in Women. J. Nutr. 2007, 137, 2128–2133. [Google Scholar] [CrossRef] [PubMed]
- Punzón-Jiménez, P.; Labarta, E. The Impact of the Female Genital Tract Microbiome in Women Health and Reproduction: A Review. J. Assist. Reprod. Genet. 2021, 38, 2519–2541. [Google Scholar] [CrossRef] [PubMed]
- Bogado Pascottini, O.; Spricigo, J.F.W.; Van Schyndel, S.J.; Mion, B.; Rousseau, J.; Weese, J.S.; LeBlanc, S.J. Effects of Parity, Blood Progesterone, and Non-Steroidal Anti-Inflammatory Treatment on the Dynamics of the Uterine Microbiota of Healthy Postpartum Dairy Cows. PLoS ONE 2021, 16, e0233943. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal Microbiome of Reproductive-Age Women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, S.; Greenbaum, G.; Moran-Gilad, J.; Weintraub, A.Y. Ecological Dynamics of the Vaginal Microbiome in Relation to Health and Disease. Am. J. Obstet. Gynecol. 2019, 220, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Fraga, M.; Perelmuter, K.; Delucchi, L.; Cidade, E.; Zunino, P. Vaginal Lactic Acid Bacteria in the Mare: Evaluation of the Probiotic Potential of Native Lactobacillus Spp. and Enterococcus Spp. Strains. Antonie Van. Leeuwenhoek 2008, 93, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Swartz, J.D.; Lachman, M.; Westveer, K.; O’Neill, T.; Geary, T.; Kott, R.W.; Berardinelli, J.G.; Hatfield, P.G.; Thomson, J.M.; Roberts, A.; et al. Characterization of the Vaginal Microbiota of Ewes and Cows Reveals a Unique Microbiota with Low Levels of Lactobacilli and Near-Neutral PH. Front. Vet. Sci. 2014, 1, 19. [Google Scholar] [CrossRef] [PubMed]
- Petrova, M.I.; Lievens, E.; Malik, S.; Imholz, N.; Lebeer, S. Lactobacillus Species as Biomarkers and Agents That Can Promote Various Aspects of Vaginal Health. Front. Physiol. 2015, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.E.; Galvão, K.N.; Rodriguez-Lecompte, J.C.; Costa, M.C. The Cattle Microbiota and the Immune System: An Evolving Field. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- Franasiak, J.M.; Scott, R.T.J. Reproductive Tract Microbiome in Assisted Reproductive Technologies. Fertil. Steril. 2015, 104, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S.; Linhares, I.M. Why Do Lactobacilli Dominate the Human Vaginal Microbiota? BJOG Int. J. Obstet. Gynaecol. 2017, 124, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.E.; Schwartz, J.A.; Robinson, C.K.; OʼHanlon, D.E.; Bradford, L.L.; He, X.; Mark, K.S.; Bruno, V.M.; Ravel, J.; Brotman, R.M. The Vaginal Microbiota and Behavioral Factors Associated With Genital Candida Albicans Detection in Reproductive-Age Women. Sex. Transm. Dis. 2019, 46, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, B.A.; Reese, S.T.; Dantas, F.G.; Franco, G.A.; Smith, T.P.L.; Adeyosoye, O.I.; Pohler, K.G.; Myer, P.R. Vaginal and Uterine Bacterial Communities in Postpartum Lactating Cows. Front. Microbiol. 2017, 8, 1047. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.M.; Holman, D.B.; Schmidt, K.N.; Pun, B.; Sedivec, K.K.; Hurlbert, J.L.; Bochantin, K.A.; Ward, A.K.; Dahlen, C.R.; Amat, S. Sequencing and Culture-Based Characterization of the Vaginal and Uterine Microbiota in Beef Cattle That Became Pregnant or Remained Open Following Artificial Insemination. Microbiol. Spectr. 2023, 11, e0273223. [Google Scholar] [CrossRef] [PubMed]
- Bicalho, M.L.S.; Santin, T.; Rodrigues, M.X.; Marques, C.E.; Lima, S.F.; Bicalho, R.C. Dynamics of the Microbiota Found in the Vaginas of Dairy Cows during the Transition Period: Associations with Uterine Diseases and Reproductive Outcome. J. Dairy. Sci. 2017, 100, 3043–3058. [Google Scholar] [CrossRef]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium Review: The Uterine Microbiome Associated with the Development of Uterine Disease in Dairy Cows. J. Dairy. Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef] [PubMed]
- Traub-Dargatz, J.L.; Salman, M.D.; Voss, J.L. Medical Problems of Adult Horses, as Ranked by Equine Practitioners. J. Am. Vet. Med. Assoc. 1991, 198, 1745–1747. [Google Scholar] [CrossRef] [PubMed]
- Canisso, I.F.; Segabinazzi, L.G.T.M.; Fedorka, C.E. Persistent Breeding-Induced Endometritis in Mares—A Multifaceted Challenge: From Clinical Aspects to Immunopathogenesis and Pathobiology. Int. J. Mol. Sci. 2020, 21, 1432. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-L.; Liu, M.-C.; Xu, J.; An, L.-G.; Wang, J.-F.; Zhu, Y.-H. Uterine Microbiota of Dairy Cows With Clinical and Subclinical Endometritis. Front. Microbiol. 2018, 9, 2691. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, Y.; Mi, J.; Zhao, Y.; Holyoak, G.R.; Yi, Z.; Wu, R.; Wang, Z.; Zeng, S. Endometrial and Vaginal Microbiome in Donkeys with and without Clinical Endometritis. Front. Microbiol. 2022, 13, 884574. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Franasiak, J.M. Endometrial Microbiota-New Player in Town. Fertil. Steril. 2017, 108, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.K.; Soffa, D.R.; McAnally, B.E.; Smith, M.S.; Hickman-Brown, K.J.; Stockland, E.L. Reproductive Microbiomes in Domestic Livestock: Insights Utilizing 16S RRNA Gene Amplicon Community Sequencing. Animals 2023, 13, 485. [Google Scholar] [CrossRef] [PubMed]
- Benner, M.; Ferwerda, G.; Joosten, I.; van der Molen, R.G. How Uterine Microbiota Might Be Responsible for a Receptive, Fertile Endometrium. Hum. Reprod. Update 2018, 24, 393–415. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.W.; Ikegami, A.; Bissada, N.F.; Herbst, M.; Redline, R.W.; Ashmead, G.G. Transmission of an Uncultivated Bergeyella Strain from the Oral Cavity to Amniotic Fluid in a Case of Preterm Birth. J. Clin. Microbiol. 2006, 44, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.J.; Cunha, F.; Vieira-Neto, A.; Bicalho, R.C.; Lima, S.; Bicalho, M.L.; Galvão, K.N. Blood as a Route of Transmission of Uterine Pathogens from the Gut to the Uterus in Cows. Microbiome 2017, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, R.M.; Arias-Álvarez, M.; Jordán-Rodríguez, D.; Rebollar, P.G.; Lorenzo, P.L.; Herranz, C.; Rodríguez, J.M. Female Reproduction and the Microbiota in Mammals: Where Are We? Theriogenology 2022, 194, 144–153. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Author(s) Objective/Hypothesis | Inclusion Criteria, (n), and Study’s Location, and Month | Sampling Type, Sequencing Platform, and Variable Region/Culture Medium | Main Taxa Identified | Main Results |
|---|---|---|---|---|
| Sathe et al., [21] * Hypothesize that the uterus of healthy mares is not sterile and is colonized by complex microflora. | Healthy mares in estrus and early pregnancy. n = 20. USA. | Uterine fluid. DNA sequencing of the 16S rRNA gene *** Qiime Software version 2 tm (data analysis) | Phylum: Bacteroidetes Proteobacteria Genera: Mares carrying embryos: Sphingobacteriales Sphingobium Mares not carrying embryos: Rhodocyclaceae and Enterobacteriaceae. | Pilot study illustrating that the uterus of horses is not a sterile environment during and after estrus, yet it can still achieve pregnancy in the presence of certain bacteria. Also, the study demonstrated that conventional culture methods are insufficient to identify bacteria in the uterus, which can be detected more accurately through high-throughput sequencing. |
| Holyoak et al., [33] | Healthy mares n = 29, USA. | Uterine fluid retrieved from small volume lavage (SVL). IlluminaV4 region of the 16S rRNA gene, amplification primers: 530F and 1100R | Phylum: Proteobacteria (100%), Firmicutes (100%), Bacteroidetes (96.2%), and Actinobacteria (100%) Genera: Pseudomonas (100%), Porphyromonas (87.5%), and Streptococcus (61.4%). | The equine uterine microbiota is diverse, although a generalized “core” microbiota was reported in all the mares in the study, there are differences based on the animal origin. |
| Jones, [31] ** A. Describe and compare the vaginal, uterine, and fecal microbiota of the mare and stallion semen. B. Evaluate the impact of raw or extended semen on the uterus and vagina microbiotas following insemination. | A. Healthy mares, n = 16, Healthy stallion n = 1, USA. B. Healthy mares n = 8, PBIE mares (Persistent breeding-induced endometritis). | Uterine fluid from SVL and endometrial swabs collected at estrus, and 48 h post-breeding for two consecutive cycles. Illumina V4 region of the 16S rRNA gene, amplification primers: 515 and 926R | Phylum: Vagina/Uterus Bacteroidetes Firmicutes Actinobacteria Proteobacteria Verrucomicrobia Genera: Uterus Corynebacterium Porphyromonas Enterobacteriaceae Streptococcus Vagina Similar to the uterus, expect no Enterobacteriaceae but RPF12 Feces (Phylum): Bacteriodetes Firmicutes Verrucomicrobia Semen (Phylum): Actinobacteria Bacteroidetes Firmicutes | A. Feces had higher diversity than semen. Uterine and vaginal had similar diversity. All samples had unique and shared microbiotas. Sample contamination could have biased results. B. The vaginal microbiota is more dynamic than the uterine microbiota after breeding, although the dominant phyla were consistent between the two organs. |
| Barba et al., [30] Characterize the vaginal microbiota in Arabian mares using traditional culture-dependent and metagenomics and identify changes in estrous cycle. | Healthy mares in estrus and diestrus. n = 8, Spain (June–July). | Vaginal swabs. Culture-dependent: Columbia blood agar (BA), Man Rogosa Sharpe (MRS) Culture independent: Illumina V3/V4 region of the 16S rRNA gene. | Phylum: Firmicutes (100%), Bacteroidetes (100%), Proteobacteria (100%), and Actinobacteria (87.5) Genera: Porphyromonas (87.5%), Campylobacter (100%), Arcanobacterium (87.5%), Corynebacterium (87.5%), Streptococcus (100%), and Fusobacterium (87.5%). | The composition and diversity of the vaginal microbiota in Arabian mares remain consistent throughout the estrus cycle. Lactobacillus spp. is not dominant in the vaginal microbiota of mares. |
| Thomson et al., [34] Characterize the uterine microbiota in mares and predict its metabolic pathways. | Healthy mares in estrus., n = 21, Chile (October). | Uterine biopsy. IlluminaV3/V4 region of the 16S rRNA gene, amplification primers: 341F and 785 R Positive and negative control | Phylum: Proteobacteria (69.6%), Firmicutes (21.1%), Bacteroidetes (7.8%), Actinobacteria (1.06%) Genera: Staphylococcus (18.88%), Pseudomonas (17.9%), Escherichia/Shigella (10.42%), and Klebsiella (9.92%). | The uterine microbiota in healthy mares is diverse, and the metabolic pathways prediction suggests that the uterus of healthy mares can produce short-chain fatty acids and amino acids. |
| Holyoak et al., [26] Describe the endometrial microbiome of mares in different geographical locations. | Mares with no reproductive history. n = 54 North America (Oklahoma, Louisiana) and Australia. | Uterine fluid retrieved by small volume lavage. IlluminaV4 region of the 16S rRNA gene, amplification primers: 515F and 806R. | Phylum: Proteobacteria (~48%), Firmicutes (30%), Bacteroidetes (12%), Actinobacteria (5%) Genera across all animals: Pseudomonas 27% Lonsdalea 8% Lactobacillus 7.5% Escherichia/Shigella 4.5% Prevotella 3% Oklahoma and Louisiana Dominated by Pseudomanas 75% Australia (only) Lonsdalea 28% Core microbiome of genera present in all samples (min abundance of 0.1%): Lactobacillus, Escherichia/Shigella, Streptococcus, Blautia, Staphylococcus, Klebsiella, Acinetobacter, and Peptoanaerobacter. | Diversity, richness, and evenness of the microbial communities of the mare’s uterus are mainly influenced by geographical location, reporting a distinct core uterine microbiome in all the mares in the study. |
| Heil et al., [15] Explores different sampling techniques to detect uterine microbiome in mares. | Mares in estrus without signs of endometritis on cytology and negative aerobic culture. n = 15, Louisiana State, USA. | Double-guarded swabs (cervix and endometrium), low-volume lavage (LVL), and endometrial biopsy Negative control; sterile unused swab (DNA isolation on same day of sample collection) IlluminaV4-V5 region of the 16S rRNA gene, amplification primers: 515F and 806R. | Phylum: Proteobacteria, Firmicutes, and Bacteroidota Genera: Klebsiella, Mycoplasma, and Aeromonas only. (Cervical swab: Proteobacteria, Firmicutes, Bacteroidota and Acidobacteria) Klebsiella, Mycoplasma, Aeromonas, and Citrobacter. | Alpha and beta diversity did not vary among the three sample techniques, suggesting that any method can be used for metagenomic identification in mares’ uteruses. However, LVL seems to be more efficient in sampling low-abundant or rare taxa compared to endometrial biopsy. Additionally, the cervical microbiota is more abundant than endometrial microbiota, but their compositions are similar. |
| Beckers et al., [32] Identify the microbiome in different sites of pregnant pony mares. | Pregnant mares (96–120 days of gestation length upon necropsy). n = 5, Louisiana State, USA. | Sterile swabs were collected from all sites (Placenta, vagina, anus, and oral cavity, Control-environmental swabs). Illumina V4 region of the 16S rRNA gene, amplification primers: 515F and 806 R | Phylum (in all sites): Firmicutes Bacteroidetes Proteobacteria Actinobacteria Genera Vagina: Rikenellceace_RC9, Porphyromonas, Campylobacter, and Streptococcus. Placenta: Gemella, Rikenellaceae_RC9, Porphyromonas, and Streptococcus. | Different richness and evenness in all samples, meaning that the microbial communities are distinct in all parts of the body tested. The placenta and oral cavity microbiome shared similarities at the genus level (Gemella and Porphyromona). Further research is needed to link the microbiome from different body sites as a biomarker of early equine placentitis. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Miranda, A.; Macnicol, J.; Orellana-Guerrero, D.; Samper, J.C.; Gomez, D.E. Reproductive Tract Microbiota of Mares. Vet. Sci. 2024, 11, 324. https://doi.org/10.3390/vetsci11070324
Gil-Miranda A, Macnicol J, Orellana-Guerrero D, Samper JC, Gomez DE. Reproductive Tract Microbiota of Mares. Veterinary Sciences. 2024; 11(7):324. https://doi.org/10.3390/vetsci11070324
Chicago/Turabian StyleGil-Miranda, Ana, Jennifer Macnicol, Daniela Orellana-Guerrero, Juan C. Samper, and Diego E. Gomez. 2024. "Reproductive Tract Microbiota of Mares" Veterinary Sciences 11, no. 7: 324. https://doi.org/10.3390/vetsci11070324
APA StyleGil-Miranda, A., Macnicol, J., Orellana-Guerrero, D., Samper, J. C., & Gomez, D. E. (2024). Reproductive Tract Microbiota of Mares. Veterinary Sciences, 11(7), 324. https://doi.org/10.3390/vetsci11070324