Recent Advances in Oral Vaccines for Animals


Abstract
:Simple Summary
Abstract
1. Introduction
2. Considerations for Designing Oral Vaccines for Animals
2.1. Challenges of the Harsh Gastrointestinal Environment
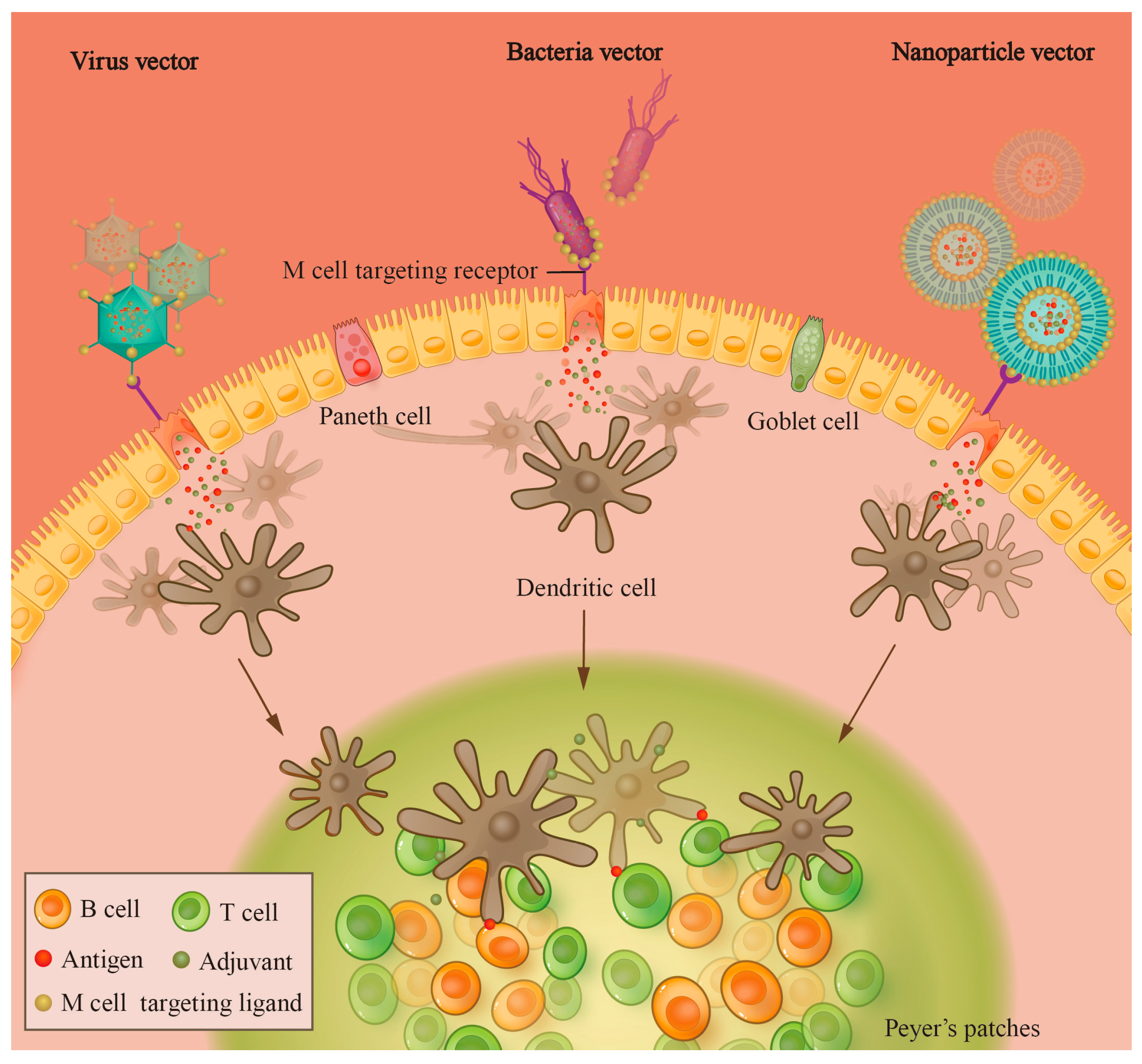
2.2. Microfold Cell
2.3. Gut Microorganisms
2.4. Adjuvants
2.4.1. Microbial Components Adjuvant
2.4.2. Cytokine Adjuvant
2.5. Bait
2.5.1. Conventional Bait
2.5.2. Novel Bait
2.5.3. Biomarkers of Bait
2.5.4. Bait Deployment Strategies
3. Delivery Strategies and Vectors of Oral Vaccines
3.1. Classical Delivery Strategies
3.2. Vectors of Nanoparticles
3.2.1. Poly Lactic Acid (PLA) and Poly Lactide-Co-Glycolide Acid (PLGA)
3.2.2. Chitosan Nanoparticle
3.2.3. Liposome
3.2.4. Bilosome
3.2.5. Virus-like Particles (VLPs)
3.3. Recombinant Bacterial and Yeast Vectors
3.4. Recombinant Virus Vectors
3.4.1. Recombinant Poxvirus Vector
3.4.2. Recombinant Adenovirus Vector
3.5. Transgenic Plant
4. Utilizations of Oral Vaccines in Various Animals
4.1. Oral Vaccines for Urban Stray Animals and Wildlife

{kind=link}
| Pathogen | Trial Animals | Type | Protection Rate | Strain/Vector | Antigen | References |
|---|---|---|---|---|---|---|
| Rabies virus | Mice | Attenuated | 90% | LBNSE-GMCSF | - | [33] |
| Mice | Attenuated | 90% | LBNSE–flagellin | - | [33] | |
| Mice | Attenuated | 100% | ERAG3G | - | [87] | |
| Ferret badgers | Attenuated | - | SRV9 | - | [81] | |
| Mice | Attenuated | 100% | ERA | - | [88] | |
| Canine | Attenuated | 100% | LBNSE-dGM-CSF | - | [50] | |
| Wolves | Attenuated | - | SAG2 | - | [89] | |
| Foxes | Attenuated | 89.6% | SPBN GASGAS | - | [90] | |
| Foxes, raccoon dogs | Attenuated | >90% | SPBN GASGAS | - | [91] | |
| Canine | Attenuated | - | SPBN GASGAS | - | [92] | |
| Foxes, raccoon dogs | Attenuated | - | SPBN GASGAS | - | [93] | |
| Mice | Attenuated | 90% | LBNSE-U-OMP19 | - | [94] | |
| Red foxes, raccoon dogs | Attenuated | - | SAD–Bern | - | [95] | |
| Dogs | Attenuated | - | SPBN GASGAS | - | [96] | |
| Mice | Recombinant vector | 50–60% | Salmonella | Glycoprotein | [97] | |
| Red foxes | Recombinant vector | 33–62% | Adenovirus | ONRAB® | [98] | |
| Skunks | Recombinant vector | 81–100% | Adenovirus | ONRAB® | [80] | |
| Ferret badgers | Recombinant vector | - | Adenovirus | Glycoprotein | [81] | |
| Mongooses | Recombinant vector | - | Adenovirus | ONRAB® | [99] | |
| Mice | Recombinant vector | 60% | Lactobacillus | Glycoprotein | [61] | |
| Goats, foxes | Recombinant vector | - | Newcastle disease virus | Glycoprotein | [100] | |
| Mycobacterium bovis | Wild boar | Inactivated | - | - | - | [101] |
| Red deer | Inactivated | - | - | - | [102] | |
| Wild badger | Attenuated | - | Liporale–BCG | - | [103] | |
| Ebola virus | Chimpanzees | Recombinant vector | - | RABV | Glycoprotein | [104] |
| Bacillus Anthracis | Mice | Attenuated | - | 34F2 spores | - | [105] |
| Toxoplasma gondii | Mice | Plant-based | - | Tobacco | HSP90-SAG1 | [17] |
| Chlamydia muridarum | Mice | Attenuated | - | CMmut/IntrOv | - | [106] |
4.2. Oral Vaccines for Economic Animals
| Pathogen | Trial Animals | Type | Protection Rate | Strain/Vector | Antigen | References |
|---|---|---|---|---|---|---|
| PRV | Domestic pigs | Attenuated | 100% | Bartha | - | [110] |
| CSFV | Pigs | Attenuated | - | C-Strain | - | [111] |
| Wild boars | Attenuated | - | C-Strain | - | [112] | |
| AIV | Mice | Recombinant vector | 100% | Baculovirus | H5N1 HA | [113] |
| Chickens | Nanoparticle | - | Sliver nanoparticles | HA DNA | [114] | |
| Mice | Recombinant vector | 100% | Salmonella | HA, NA | [115] | |
| Mice | Recombinant vector | 100% | Yeast | H7N9 HA | [116] | |
| Chickens | Recombinant vector | 100% | Yeast | H5N1 HA | [117] | |
| Chickens | Recombinant vector | 100% | Lactococcus lactis | HA1 + M2 | [118] | |
| Mice | Nanoparticle | 100% | VLPs | H5N1 HA + VSP | [60] | |
| PEDV | Mice, piglets | Recombinant vector | - | Yeast | S1 | [119] |
| Mice | Recombinant vector | - | Lactobacillus casei | COE | [63] | |
| Piglets | Recombinant vector | - | Bacillus subtilis | COE | [120] | |
| Mice | Recombinant vector | - | Adenovirus | COE + LTB | [121] | |
| Piglets | Plant-based | - | Maize grain | Spike protein | [83] | |
| Mice | Plant-based | - | Nicotiana benthamiana | COE-PIGs | [122] | |
| Mice | Recombinant vector | - | Lactobacillus | S1 | [62] | |
| Clostridium tetani | Mice | Nanoparticle | - | GM-bilosomes | Tetanus toxoid | [58] |
| Campylobacter jejuni | Chickens | Nanoparticle | - | PLGA | OMPs | [36] |
| Newcastle disease virus | Chickens | Attenuated | - | I2 | - | [123] |
| Clostridium perfringens | Chickens | Plant-based | - | Tobacco | NetB, alpha-toxin, metallopeptidase | [124] |
| Broilers | Recombinant vector | - | Lactobacillus casei | α-Toxin | [125] | |
| Duck tembusu virus | Ducks | Recombinant vector | 100% | Salmonella | PrM, E DNA | [126] |
| Ducks | Recombinant vector | 100% | Salmonella | Capsid protein DNA | [127] | |
| Rabbit hemorrhagic disease virus | Rabbits | Recombinant vector | 93.3% | Salmonella | VP60 DNA | [128] |
| Nipah virus | Pigs | Recombinant vector | - | Attenuated RABV | NiVG, NiVF | [129] |
| FMDV | Mice | Recombinant vector | - | Lactococcus lactis | VP1 | [130] |
| Guinea pigs | Nanoparticle | - | PLGA | VP1, VP3 DNA | [38] | |
| Brucella | Mice | Recombinant vector | - | Salmonella | PrVgB | [131] |
| Leptospira spp. | Rats | Recombinant vector | - | Salmonella | LipL32 | [132] |
| Trichinella spiralis | Mice | Recombinant vector | - | Salmonella | Ts43 | [133] |
| Mycoplasma hyopneumoniae | Piglets | Nanoparticle | - | Silica SBA-15 | Extraction proteins | [134] |
| IBDV | Hens | Plant-based | - | Nicotiana benthamiana | VP2 | [135] |
| Mice | Recombinant vector | - | Yeast | VP2 | [136] | |
| E. coli | Mice | Plant-based | - | Canola seeds | STxB, CfaB, LTB, Intimin | [82] |
| Avian | Recombinant vector | 100% | Salmonella | O-antigen | [137] | |
| Cattle | Attenuated | - | E16991,E16992,E16993 | - | [138] | |
| Piglets | Modified subunit | - | αAPN-pIgA | FedF | [139] | |
| Salmonella | Mice | Attenuated | 100% | KST0556 | - | [140] |
| Rabbits | Attenuated | 100% | HB1 | - | [141] | |
| Chickens | Nanoparticle | - | Chitosan | OMPs, Flagellin protein | [142] | |
| Rhesus macaque | Attenuated | 80% | CVD 1926 | - | [143] | |
| Hens | Nanoparticle | - | PVM/MA | Heat extract fraction | [144] | |
| Chickens | Attenuated | 80% | SG01 | - | [145] | |
| Chickens | Attenuated | - | 2S G10 | - | [146] | |
| Broilers | Nanoparticle | - | Chitosan | OMPs, Flagella proteins | [53] | |
| Staphylococcus aureus | Mice | Recombinant vector | - | Salmonella | rEsxAB, rHlam | [147] |
| Mannheimia haemolytica | Mice | Plant-based | - | Nicotiana benthamiana | LktA + PlpE | [148] |
| Eimeria tenella | Chickens | Recombinant vector | - | Yeast | EtAMA1, EtIMP1, EtMIC3 | [149] |
| Chickens | Recombinant vector | - | Lactobacillus plantarum | RON2 | [150] | |
| Goose parvovirus | Ducks | Nanoparticle | - | Cyclic peptide nanotubes | VP2 DNA | [151] |
| Mycobacterium avium subsp. paratuberculosis | Calves | Attenuated | - | BacAΔ | - | [152] |
4.3. Oral Vaccines for Aquaculture
4.4. Approved Oral Vaccine for Animals
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parida, S. Vaccination against Foot-and-Mouth Disease Virus: Strategies and Effectiveness. Expert Rev. Vaccines 2009, 8, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V. Introduction to Classical Swine Fever: Virus, Disease and Control Policy. Vet. Microbiol. 2000, 73, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Alkie, T.N.; Rautenschlein, S. Infectious Bursal Disease Virus in Poultry: Current Status and Future Prospects. Vet. Med. 2016, 7, 9–18. [Google Scholar] [CrossRef]
- Horzinek, M.C. Rinderpest: The Second Viral Disease Eradicated. Vet. Microbiol. 2011, 149, 295–297. [Google Scholar] [CrossRef]
- Dwivedy, A.; Aich, P. Importance of Innate Mucosal Immunity and the Promises It Holds. Int. J. Gen. Med. 2011, 4, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.J.; Eschmann, M.; Mantis, N.J. Inhibition of Salmonella enterica Serovar Typhimurium Motility and Entry into Epithelial Cells by a Protective Antilipopolysaccharide Monoclonal Immunoglobulin A Antibody. Infect. Immun. 2008, 76, 4137–4144. [Google Scholar] [CrossRef] [PubMed]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.C.; Kaetzel, C.S. Secretory IgA Is Concentrated in the Outer Layer of Colonic Mucus along with Gut Bacteria. Pathogens 2014, 3, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s Complex Roles in Immunity and Mucosal Homeostasis in the Gut. Mucosal. Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Iannino, F.; Uriza, P.J.; Duarte, C.M.; Pepe, M.V.; Roset, M.S.; Briones, G. Development of a Salmonella-Based Oral Vaccine to Control Intestinal Colonization of Shiga-Toxin-Producing E. coli (STEC) in Animals. Vaccine 2022, 40, 1065–1073. [Google Scholar] [CrossRef]
- Kwong, K.W.-Y.; Xin, Y.; Lai, N.C.-Y.; Sung, J.C.-C.; Wu, K.-C.; Hamied, Y.K.; Sze, E.T.-P.; Lam, D.M.-K. Oral Vaccines: A Better Future of Immunization. Vaccines 2023, 11, 1232. [Google Scholar] [CrossRef]
- Blume, S.; Geesink, I. A Brief History of Polio Vaccines. Science 2000, 288, 1593–1594. [Google Scholar] [CrossRef]
- Howard, B.D. A Prototype Live Oral Cholera Vaccine. Nature 1971, 230, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liang, B.; Wang, W.; Li, L.; Feng, N.; Zhao, Y.; Wang, T.; Yan, F.; Yang, S.; Xia, X. Viral Vectored Vaccines: Design, Development, Preventive and Therapeutic Applications in Human Diseases. Signal Transduct. Target. Ther. 2023, 8, 149. [Google Scholar] [CrossRef]
- Li, J.-X.; Hou, L.-H.; Meng, F.-Y.; Wu, S.-P.; Hu, Y.-M.; Liang, Q.; Chu, K.; Zhang, Z.; Xu, J.-J.; Tang, R.; et al. Immunity Duration of a Recombinant Adenovirus Type-5 Vector-Based Ebola Vaccine and a Homologous Prime-Boost Immunisation in Healthy Adults in China: Final Report of a Randomised, Double-Blind, Placebo-Controlled, Phase 1 Trial. Lancet Glob. Health 2017, 5, e324–e334. [Google Scholar] [CrossRef]
- Nooraei, S.; Sarkar Lotfabadi, A.; Akbarzadehmoallemkolaei, M.; Rezaei, N. Immunogenicity of Different Types of Adjuvants and Nano-Adjuvants in Veterinary Vaccines: A Comprehensive Review. Vaccines 2023, 11, 453. [Google Scholar] [CrossRef]
- Kwon, K.-C.; Daniell, H. Oral Delivery of Protein Drugs Bioencapsulated in Plant Cells. Mol. Ther. 2016, 24, 1342–1350. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, E.F.; Corigliano, M.G.; Oliferuk, S.; Ramos-Duarte, V.A.; Rivera, M.; Mendoza-Morales, L.F.; Angel, S.O.; Sander, V.A.; Clemente, M. Oral Immunization With a Plant HSP90-SAG1 Fusion Protein Produced in Tobacco Elicits Strong Immune Responses and Reduces Cyst Number and Clinical Signs of Toxoplasmosis in Mice. Front. Plant Sci. 2021, 12, 726910. [Google Scholar] [CrossRef] [PubMed]
- Shahid, N.; Daniell, H. Plant-Based Oral Vaccines against Zoonotic and Non-Zoonotic Diseases. Plant Biotechnol. J. 2016, 14, 2079–2099. [Google Scholar] [CrossRef]
- Hartenstein, V.; Martinez, P. Structure, Development and Evolution of the Digestive System. Cell Tissue Res. 2019, 377, 289–292. [Google Scholar] [CrossRef]
- Pavot, V.; Rochereau, N.; Genin, C.; Verrier, B.; Paul, S. New Insights in Mucosal Vaccine Development. Vaccine 2012, 30, 142–154. [Google Scholar] [CrossRef]
- Vela Ramirez, J.E.; Sharpe, L.A.; Peppas, N.A. Current State and Challenges in Developing Oral Vaccines. Adv. Drug Deliv. Rev. 2017, 114, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.; Du, G.; Chen, X.; Sun, X. Advancedoral Vaccine Delivery Strategies for Improving the Immunity. Adv. Drug Deliv. Rev. 2021, 177, 113928. [Google Scholar] [CrossRef] [PubMed]
- Davitt, C.J.H.; Lavelle, E.C. Delivery Strategies to Enhance Oral Vaccination against Enteric Infections. Adv. Drug Deliv. Rev. 2015, 91, 52–69. [Google Scholar] [CrossRef] [PubMed]
- Misumi, S.; Masuyama, M.; Takamune, N.; Nakayama, D.; Mitsumata, R.; Matsumoto, H.; Urata, N.; Takahashi, Y.; Muneoka, A.; Sukamoto, T.; et al. Targeted Delivery of Immunogen to Primate m Cells with Tetragalloyl Lysine Dendrimer. J. Immunol. 2009, 182, 6061–6070. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-S.; Piao, D.-C.; Jiang, T.; Bok, J.-D.; Cho, C.-S.; Lee, Y.-S.; Kang, S.-K.; Choi, Y.-J. Recombinant Interleukin 6 with M Cell-Targeting Moiety Produced in Lactococcus Lactis IL1403 as a Potent Mucosal Adjuvant for Peroral Immunization. Vaccine 2015, 33, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Yoo, M.-K.; Kang, S.-K.; Choi, J.-H.; Park, I.-K.; Na, H.-S.; Lee, H.-C.; Kim, E.-B.; Lee, N.-K.; Nah, J.-W.; Choi, Y.-J.; et al. Targeted Delivery of Chitosan Nanoparticles to Peyer’s Patch Using M Cell-Homing Peptide Selected by Phage Display Technique. Biomaterials 2010, 31, 7738–7747. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, Z.; Li, X.; Li, J.; Lv, J.; Ma, Z.; Pan, L. Immune Responses to Orally Administered Recombinant Lactococcus Lactis Expressing Multi-Epitope Proteins Targeting M Cells of Foot-and-Mouth Disease Virus. Viruses 2021, 13, 2036. [Google Scholar] [CrossRef] [PubMed]
- Agirman, G.; Yu, K.B.; Hsiao, E.Y. Signaling Inflammation across the Gut-Brain Axis. Science 2021, 374, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. The Influence of the Intestinal Microbiome on Vaccine Responses. Vaccine 2018, 36, 4433–4439. [Google Scholar] [CrossRef]
- Chen, X.; Chen, X.; Qiu, S.; Hu, Y.; Jiang, C.; Wang, D.; Fan, Q.; Zhang, C.; Huang, Y.; Yu, Y.; et al. Effects of Epimedium Polysaccharide-Propolis Flavone Oral Liquid on Mucosal Immunity in Chickens. Int. J. Biol. Macromol. 2014, 64, 6–10. [Google Scholar] [CrossRef]
- Fingerut, E.; Gutter, B.; Goldway, M.; Eliahoo, D.; Pitcovski, J. B Subunit of E. coli Enterotoxin as Adjuvant and Carrier in Oral and Skin Vaccination. Vet. Immunol. Immunopathol. 2006, 112, 253–263. [Google Scholar] [CrossRef]
- Means, T.K.; Hayashi, F.; Smith, K.D.; Aderem, A.; Luster, A.D. The Toll-like Receptor 5 Stimulus Bacterial Flagellin Induces Maturation and Chemokine Production in Human Dendritic Cells. J. Immunol. 2003, 170, 5165–5175. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, G.; Ren, G.; Gnanadurai, C.W.; Li, Z.; Chai, Q.; Yang, Y.; Leyson, C.M.; Wu, W.; Cui, M.; et al. Recombinant Rabies Viruses Expressing GM-CSF or Flagellin Are Effective Vaccines for Both Intramuscular and Oral Immunizations. PLoS ONE 2013, 8, e63384. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yamamoto, S.; Kataoka, T.; Tokunaga, T. Lipofection of Synthetic Oligodeoxyribonucleotide Having a Palindromic Sequence of AACGTT to Murine Splenocytes Enhances Interferon Production and Natural Killer Activity. Microbiol. Immunol. 1994, 38, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Latz, E.; Schoenemeyer, A.; Visintin, A.; Fitzgerald, K.A.; Monks, B.G.; Knetter, C.F.; Lien, E.; Nilsen, N.J.; Espevik, T.; Golenbock, D.T. TLR9 Signals after Translocating from the ER to CpG DNA in the Lysosome. Nat. Immunol. 2004, 5, 190–198. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Hodgins, D.C.; Alkie, T.N.; Quinteiro-Filho, W.; Yitbarek, A.; Astill, J.; Sharif, S. Oral Administration of PLGA-Encapsulated CpG ODN and Campylobacter jejuni Lysate Reduces Cecal Colonization by Campylobacter jejuni in Chickens. Vaccine 2018, 36, 388–394. [Google Scholar] [CrossRef]
- Harrell, J.E.; Kurtz, J.R.; Bauer, D.L.; Prior, J.T.; Gellings, P.S.; Morici, L.A.; McLachlan, J.B. An Outer Membrane Vesicle-Adjuvanted Oral Vaccine Protects Against Lethal, Oral Salmonella Infection. Pathogens 2021, 10, 616. [Google Scholar] [CrossRef]
- Yang, Y.; Teng, Z.; Lu, Y.; Luo, X.; Mu, S.; Ru, J.; Zhao, X.; Guo, H.; Ran, X.; Wen, X.; et al. Enhanced Immunogenicity of Foot and Mouth Disease DNA Vaccine Delivered by PLGA Nanoparticles Combined with Cytokine Adjuvants. Res. Vet. Sci. 2021, 136, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, B.; Wang, M. GM-CSF and IL-2 as Adjuvant Enhance the Immune Effect of Protein Vaccine against Foot-and-Mouth Disease. Virol. J. 2011, 8, 7. [Google Scholar] [CrossRef]
- Caligiuri, M.A.; Murray, C.; Robertson, M.J.; Wang, E.; Cochran, K.; Cameron, C.; Schow, P.; Ross, M.E.; Klumpp, T.R.; Soiffer, R.J. Selective Modulation of Human Natural Killer Cells in Vivo after Prolonged Infusion of Low Dose Recombinant Interleukin 2. J. Clin. Investig. 1993, 91, 123–132. [Google Scholar] [CrossRef]
- Bonwitt, J.; Bonaparte, S.; Blanton, J.; Gibson, A.D.; Hoque, M.; Kennedy, E.; Islam, K.; Siddiqi, U.R.; Wallace, R.M.; Azam, S. Oral Bait Preferences and Feasibility of Oral Rabies Vaccination in Bangladeshi Dogs. Vaccine 2020, 38, 5021–5026. [Google Scholar] [CrossRef] [PubMed]
- Gowtage, S.; Williams, G.A.; Henderson, R.; Aylett, P.; MacMorran, D.; Palmer, S.; Robertson, A.; Lesellier, S.; Carter, S.P.; Chambers, M.A. Testing of a Palatable Bait and Compatible Vaccine Carrier for the Oral Vaccination of European Badgers (Meles meles) against Tuberculosis. Vaccine 2017, 35, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.R.; Crider, N.J.; Weyer, G.A.; Tosh, R.D.; VerCauteren, K.C. Bait Development For Oral Delivery Of Pharmaceuticals To Raccoons (Procyon Lotor) And Striped Skunks (Mephitis Mephitis). J. Wildl. Dis. 2016, 52, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Sage, M.; Fourel, I.; Lahoreau, J.; Siat, V.; Berny, P.; Rossi, S. Iophenoxic Acid Derivatives as Markers of Oral Baits to Wildlife. New Tools for Their Detection in Tissues of a Game Species and Safety Considerations for Human Exposure. Environ. Sci. Pollut. Res. Int. 2013, 20, 2893–2904. [Google Scholar] [CrossRef] [PubMed]
- Robardet, E.; Rieder, J.; Barrat, J.; Cliquet, F. Reconsidering Oral Rabies Vaccine Bait Uptake Evaluation at Population Level: A Simple, Noninvasive, and Ethical Method by Fecal Survey Using a Physical Biomarker. J. Wildl. Dis. 2019, 55, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Langguth, A.; Leelahapongsathon, K.; Wannapong, N.; Kasemsuwan, S.; Ortmann, S.; Vos, A.; Böer, M. Comparative Study of Optical Markers to Assess Bait System Efficiency Concerning Vaccine Release in the Oral Cavity of Dogs. Viruses 2021, 13, 1382. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.P.; Robertson, A.; Palphramand, K.L.; Chambers, M.A.; McDonald, R.A.; Delahay, R.J. Bait Uptake by Wild Badgers and Its Implications for Oral Vaccination against Tuberculosis. PLoS ONE 2018, 13, e0206136. [Google Scholar] [CrossRef] [PubMed]
- Tizard, I.R. Chapter 8—The Administration of Vaccines. In Vaccines for Veterinarians; Tizard, I.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 87–104.e1. ISBN 978-0-323-68299-2. [Google Scholar]
- Vos, A.; Freuling, C.M.; Hundt, B.; Kaiser, C.; Nemitz, S.; Neubert, A.; Nolden, T.; Teifke, J.P.; Te Kamp, V.; Ulrich, R.; et al. Oral Vaccination of Wildlife against Rabies: Differences among Host Species in Vaccine Uptake Efficiency. Vaccine 2017, 35, 3938–3944. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, L.; Zhou, S.; Wang, Z.; Ruan, J.; Tang, L.; Jia, Z.; Cui, M.; Zhao, L.; Fu, Z.F. Recombinant Rabies Virus Expressing Dog GM-CSF Is an Efficacious Oral Rabies Vaccine for Dogs. Oncotarget 2015, 6, 38504–38516. [Google Scholar] [CrossRef]
- Lee, Y.; Kamada, N.; Moon, J.J. Oral Nanomedicine for Modulating Immunity, Intestinal Barrier Functions, and Gut Microbiome. Adv. Drug Deliv. Rev. 2021, 179, 114021. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-Based Nanoparticles: An Overview of Biomedical Applications. J. Control Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Villanueva, K.; Akerele, G.; Al-Hakeem, W.; Adams, D.; Gourapura, R.; Selvaraj, R. Immunization of Broiler Chickens With a Killed Chitosan Nanoparticle Salmonella Vaccine Decreases Salmonella Enterica Serovar Enteritidis Load. Front. Physiol. 2022, 13, 920777. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, D.; Cavaco-Paulo, A.; Nogueira, E. Design of Liposomes as Drug Delivery System for Therapeutic Applications. Int. J. Pharm. 2021, 601, 120571. [Google Scholar] [CrossRef]
- Defrise-Quertain, F.; Chatelain, P.; Ruysschaert, J.M.; Delmelle, M. Spin Label Partitioning in Lipid Vesicles. A Model Study for Drug Encapsulation. Biochim. Biophys. Acta 1980, 628, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Wilkhu, J.S.; McNeil, S.E.; Anderson, D.E.; Perrie, Y. Characterization and Optimization of Bilosomes for Oral Vaccine Delivery. J. Drug Target. 2013, 21, 291–299. [Google Scholar] [CrossRef]
- Conacher, M.; Alexander, J.; Brewer, J.M. Oral Immunisation with Peptide and Protein Antigens by Formulation in Lipid Vesicles Incorporating Bile Salts (Bilosomes). Vaccine 2001, 19, 2965–2974. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Harde, H.; Indulkar, A.; Agrawal, A.K. Improved Stability and Immunological Potential of Tetanus Toxoid Containing Surface Engineered Bilosomes Following Oral Administration. Nanomedicine 2014, 10, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Liu, D.; Booth, G.; Gao, W.; Lu, Y. Virus-Like Particle Engineering: From Rational Design to Versatile Applications. Biotechnol. J. 2018, 13, e1700324. [Google Scholar] [CrossRef]
- Serradell, M.C.; Rupil, L.L.; Martino, R.A.; Prucca, C.G.; Carranza, P.G.; Saura, A.; Fernández, E.A.; Gargantini, P.R.; Tenaglia, A.H.; Petiti, J.P.; et al. Efficient Oral Vaccination by Bioengineering Virus-like Particles with Protozoan Surface Proteins. Nat. Commun. 2019, 10, 361. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, H.; Yang, R.; Zhang, S.; Zhao, W.; Hu, J.; Jiang, Y.; Yang, W.; Huang, H.; Shi, C.; et al. Construction and Evaluation of Recombinant Lactobacillus plantarum NC8 Delivering One Single or Two Copies of G Protein Fused with a DC-Targeting Peptide (DCpep) as Novel Oral Rabies Vaccine. Vet. Microbiol. 2020, 251, 108906. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, X.; Li, Y.; Li, F.; Zhao, H.; Shao, Y.; Zhang, L.; Ding, G.; Li, J.; Jiang, Y.; et al. Evaluation of the Immunogenicity in Mice Orally Immunized with Recombinant Lactobacillus Casei Expressing Porcine Epidemic Diarrhea Virus S1 Protein. Viruses 2022, 14, 890. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.N.; Wang, L.; Zheng, D.Z.; Chen, S.; Shi, W.; Qiao, X.Y.; Jiang, Y.P.; Tang, L.J.; Xu, Y.G.; Li, Y.J. Oral Immunization with a Lactobacillus Casei-Based Anti-Porcine Epidemic Diarrhoea Virus (PEDV) Vaccine Expressing Microfold Cell-Targeting Peptide Co1 Fused with the COE Antigen of PEDV. J. Appl. Microbiol. 2018, 124, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Tam, M.A.; Rydström, A.; Sundquist, M.; Wick, M.J. Early Cellular Responses to Salmonella Infection: Dendritic Cells, Monocytes, and More. Immunol. Rev. 2008, 225, 140–162. [Google Scholar] [CrossRef]
- Curtiss, R.; Nakayama, K.; Kelly, S.M. Recombinant Avirulent Salmonella Vaccine Strains with Stable Maintenance and High Level Expression of Cloned Genes in Vivo. Immunol. Investig. 1989, 18, 583–596. [Google Scholar] [CrossRef]
- Senevirathne, A.; Hewawaduge, C.; Park, S.; Jawalagatti, V.; Kim, C.; Seo, B.-J.; Lee, E.; Lee, J.H. Single Oral Immunization of an Attenuated Salmonella Gallinarium Formulation Consisting of Equal Quantities of Strains Secreting H9N2 Hemagglutinin-HA1, HA2, and M2eCD154 Induces Significant Protection against H9N2 and Partial Protection against Salmonella Gallinarium Challenge in Chickens. Vet. Immunol. Immunopathol. 2021, 240, 110318. [Google Scholar] [CrossRef] [PubMed]
- Grover, A.; McLean, J.L.; Troudt, J.M.; Foster, C.; Izzo, L.; Creissen, E.; MacDonald, E.; Troy, A.; Izzo, A.A. Heat Killed Saccharomyces Cerevisiae as an Adjuvant for the Induction of Vaccine-Mediated Immunity against Infection with Mycobacterium Tuberculosis. Vaccine 2016, 34, 2798–2805. [Google Scholar] [CrossRef]
- Cascio, V.; Gittings, D.; Merloni, K.; Hurton, M.; Laprade, D.; Austriaco, N. S-Adenosyl-L-Methionine Protects the Probiotic Yeast, Saccharomyces Boulardii, from Acid-Induced Cell Death. BMC Microbiol. 2013, 13, 35. [Google Scholar] [CrossRef]
- Lei, H.; Jin, S.; Karlsson, E.; Schultz-Cherry, S.; Ye, K. Yeast Surface-Displayed H5N1 Avian Influenza Vaccines. J. Immunol. Res. 2016, 2016, 4131324. [Google Scholar] [CrossRef]
- Ivanova, E. Yeasts in Nanotechnology-Enabled Oral Vaccine and Gene Delivery. Bioengineered 2021, 12, 8325–8335. [Google Scholar] [CrossRef]
- Maki, J.; Guiot, A.-L.; Aubert, M.; Brochier, B.; Cliquet, F.; Hanlon, C.A.; King, R.; Oertli, E.H.; Rupprecht, C.E.; Schumacher, C.; et al. Oral Vaccination of Wildlife Using a Vaccinia-Rabies-Glycoprotein Recombinant Virus Vaccine (RABORAL V-RG®): A Global Review. Vet. Res. 2017, 48, 57. [Google Scholar] [CrossRef]
- Roess, A.A.; Rea, N.; Lederman, E.; Dato, V.; Chipman, R.; Slate, D.; Reynolds, M.G.; Damon, I.K.; Rupprecht, C.E. National Surveillance for Human and Pet Contact with Oral Rabies Vaccine Baits, 2001-2009. J. Am. Vet. Med. Assoc. 2012, 240, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.; Johnson, S.; Walker, N.; Wickham, C.; Beath, A.; VerCauteren, K. Efficacy of Ontario Rabies Vaccine Baits (ONRAB) against Rabies Infection in Raccoons. Vaccine 2018, 36, 4919–4926. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-C.; Guan, X.-H.; Li, Y.-H.; Huang, J.-Y.; Jiang, T.; Hou, L.-H.; Li, J.-X.; Yang, B.-F.; Wang, L.; Wang, W.-J.; et al. Immunogenicity and Safety of a Recombinant Adenovirus Type-5-Vectored COVID-19 Vaccine in Healthy Adults Aged 18 Years or Older: A Randomised, Double-Blind, Placebo-Controlled, Phase 2 Trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and Efficacy of the ChAdOx1 nCoV-19 Vaccine (AZD1222) against SARS-CoV-2: An Interim Analysis of Four Randomised Controlled Trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Rosatte, R.C.; Donovan, D.; Davies, J.C.; Allan, M.; Bachmann, P.; Stevenson, B.; Sobey, K.; Brown, L.; Silver, A.; Bennett, K.; et al. Aerial Distribution of ONRAB Baits as a Tactic to Control Rabies in Raccoons and Striped Skunks in Ontario, Canada. J. Wildl. Dis. 2009, 45, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Taylor, J.S.; Davies, J.C.; Donovan, D. Immune Response and Protection in Raccoons (Procyon lotor) Following Consumption of Baits Containing ONRAB®, a Human Adenovirus Rabies Glycoprotein Recombinant Vaccine. J. Wildl. Dis. 2012, 48, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Sobey, K.G.; Jamieson, S.E.; Walpole, A.A.; Rosatte, R.C.; Donovan, D.; Fehlner-Gardiner, C.; Nadin-Davis, S.A.; Davies, J.C.; Kyle, C.J. ONRAB® Oral Rabies Vaccine Is Shed from, but Does Not Persist in, Captive Mammals. Vaccine 2019, 37, 4310–4317. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.K.; Nadin-Davis, S.A.; Sheen, M.; Rosatte, R.; Mueller, R.; Beresford, A. Safety Studies on an Adenovirus Recombinant Vaccine for Rabies (AdRG1.3-ONRAB) in Target and Non-Target Species. Vaccine 2009, 27, 6619–6626. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Ellison, J.A.; Jackson, F.R.; Bachmann, P.; Taylor, J.S.; Franka, R.; Donovan, D. Oral Vaccination and Protection of Striped Skunks (Mephitis mephitis) against Rabies Using ONRAB®. Vaccine 2014, 32, 3675–3679. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, Y.; Zhang, S.; Fang, L.; Zhang, F.; Hu, R. Experimental Oral Immunization of Ferret Badgers (Melogale Moschata) with a Recombinant Canine Adenovirus Vaccine CAV-2-E3Δ-RGP and an Attenuated Rabies Virus SRV9. J. Wildl. Dis. 2014, 50, 374–377. [Google Scholar] [CrossRef]
- Shojaei Jeshvaghani, F.; Amani, J.; Kazemi, R.; Karimi Rahjerdi, A.; Jafari, M.; Abbasi, S.; Salmanian, A.H. Oral Immunization with a Plant-Derived Chimeric Protein in Mice: Toward the Development of a Multipotent Edible Vaccine against E. coli O157: H7 and ETEC. Immunobiology 2019, 224, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Maj, M.; Fake, G.M.; Walker, J.H.; Saltzman, R.; Howard, J.A. Oral Administration of Coronavirus Spike Protein Provides Protection to Newborn Pigs When Challenged with PEDV. Vaccines 2021, 9, 1416. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.; Freuling, C.; Ortmann, S.; Kretzschmar, A.; Mayer, D.; Schliephake, A.; Müller, T. An Assessment of Shedding with the Oral Rabies Virus Vaccine Strain SPBN GASGAS in Target and Non-Target Species. Vaccine 2018, 36, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Ortmann, S.; Kretzschmar, A.; Kaiser, C.; Lindner, T.; Freuling, C.; Kaiser, C.; Schuster, P.; Mueller, T.; Vos, A. In Vivo Safety Studies With SPBN GASGAS in the Frame of Oral Vaccination of Foxes and Raccoon Dogs Against Rabies. Front. Vet. Sci. 2018, 5, 91. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Dietzschold, B.; Li, J. Immunogenicity and Safety of Recombinant Rabies Viruses Used for Oral Vaccination of Stray Dogs and Wildlife. Zoonoses Public Health 2009, 56, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-K.; Kim, H.-H.; Choi, S.-S.; Kim, J.-T.; Jeong, W.-H.; Song, J.-Y. Oral Immunization of Mice with Recombinant Rabies Vaccine Strain (ERAG3G) Induces Complete Protection. Clin. Exp. Vaccine Res. 2015, 4, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Shuai, L.; Feng, N.; Wang, X.; Ge, J.; Wen, Z.; Chen, W.; Qin, L.; Xia, X.; Bu, Z. Genetically Modified Rabies Virus ERA Strain Is Safe and Induces Long-Lasting Protective Immune Response in Dogs after Oral Vaccination. Antiviral. Res. 2015, 121, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Sillero-Zubiri, C.; Marino, J.; Gordon, C.H.; Bedin, E.; Hussein, A.; Regassa, F.; Banyard, A.; Fooks, A.R. Feasibility and Efficacy of Oral Rabies Vaccine SAG2 in Endangered Ethiopian Wolves. Vaccine 2016, 34, 4792–4798. [Google Scholar] [CrossRef]
- Freuling, C.M.; Kamp, V.T.; Klein, A.; Günther, M.; Zaeck, L.; Potratz, M.; Eggerbauer, E.; Bobe, K.; Kaiser, C.; Kretzschmar, A.; et al. Long-Term Immunogenicity and Efficacy of the Oral Rabies Virus Vaccine Strain SPBN GASGAS in Foxes. Viruses 2019, 11, 790. [Google Scholar] [CrossRef]
- Freuling, C.M.; Eggerbauer, E.; Finke, S.; Kaiser, C.; Kaiser, C.; Kretzschmar, A.; Nolden, T.; Ortmann, S.; Schröder, C.; Teifke, J.P.; et al. Efficacy of the Oral Rabies Virus Vaccine Strain SPBN GASGAS in Foxes and Raccoon Dogs. Vaccine 2019, 37, 4750–4757. [Google Scholar] [CrossRef]
- Smith, T.G.; Millien, M.; Vos, A.; Fracciterne, F.A.; Crowdis, K.; Chirodea, C.; Medley, A.; Chipman, R.; Qin, Y.; Blanton, J.; et al. Evaluation of Immune Responses in Dogs to Oral Rabies Vaccine under Field Conditions. Vaccine 2019, 37, 4743–4749. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.; Nokireki, T.; Isomursu, M.; Gadd, T.; Kovacs, F. Oral Vaccination of Foxes and Raccoon Dogs against Rabies with the 3rd Generation Oral Rabies Virus Vaccine, SPBN GASGAS, in Finland. Acta Vet. Scand. 2021, 63, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Y.; Chen, Y.; Zhang, J.; Pei, J.; Cui, M.; Fu, Z.F.; Zhao, L.; Zhou, M. A Novel Oral Rabies Vaccine Enhances the Immunogenicity through Increasing Dendritic Cells Activation and Germinal Center Formation by Expressing U-OMP19 in a Mouse Model. Emerg. Microbes. Infect. 2021, 10, 913–928. [Google Scholar] [CrossRef] [PubMed]
- Zienius, D.; Mickutė, J.; Pautienius, A.; Grigas, J.; Stankevičius, A.; Pridotkas, G.; Jacevičius, E.; Kemeraitė, J.; Jacevičienė, I. Analysis of Seroprevalence in Target Wildlife during the Oral Rabies Vaccination Programme in Lithuania. Acta Vet. Scand. 2021, 63, 12. [Google Scholar] [CrossRef]
- Megawati Saputra, I.L.; Suwarno, S.; Husein, W.F.; Suseno, P.P.; Prayoga, I.M.A.; Vos, A.; Arthawan, I.M.; Schoonman, L.; Weaver, J.; Zainuddin, N. Immunogenicity of Oral Rabies Vaccine Strain SPBN GASGAS in Local Dogs in Bali, Indonesia. Viruses 2023, 15, 1405. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, J.; Wu, X.; Yu, L.; Chen, H.; Guo, H.; Zhang, M.; Li, H.; Liu, X.; Sun, S.; et al. Oral Immunisation of Mice with a Recombinant Rabies Virus Vaccine Incorporating the Heat-Labile Enterotoxin B Subunit of E. coli in an Attenuated Salmonella Strain. Res. Vet. Sci. 2012, 93, 675–681. [Google Scholar] [CrossRef]
- Brown, L.J.; Rosatte, R.C.; Fehlner-Gardiner, C.; Bachmann, P.; Ellison, J.A.; Jackson, F.R.; Taylor, J.S.; Davies, C.; Donovan, D. Oral Vaccination and Protection of Red Foxes (Vulpes vulpes) against Rabies Using ONRAB, an Adenovirus-Rabies Recombinant Vaccine. Vaccine 2014, 32, 984–989. [Google Scholar] [CrossRef]
- Berentsen, A.R.; Leinbach, I.L.; Rivera-Rodriguez, M.J.; Gilbert, A.T. Oral Rabies Vaccination of Small Indian Mongooses (Urva Auropunctata) with ONRAB via Ultralite Baits. Viruses 2021, 13, 734. [Google Scholar] [CrossRef]
- Murr, M.; Freuling, C.; Pérez-Bravo, D.; Grund, C.; Mettenleiter, T.C.; Römer-Oberdörfer, A.; Müller, T.; Finke, S. Immune Response after Oral Immunization of Goats and Foxes with an NDV Vectored Rabies Vaccine Candidate. PLoS Negl. Trop Dis. 2024, 18, e0011639. [Google Scholar] [CrossRef]
- Garrido, J.M.; Sevilla, I.A.; Beltrán-Beck, B.; Minguijón, E.; Ballesteros, C.; Galindo, R.C.; Boadella, M.; Lyashchenko, K.P.; Romero, B.; Geijo, M.V.; et al. Protection against Tuberculosis in Eurasian Wild Boar Vaccinated with Heat-Inactivated Mycobacterium Bovis. PLoS ONE 2011, 6, e24905. [Google Scholar] [CrossRef]
- Thomas, J.; Risalde, M.Á.; Serrano, M.; Sevilla, I.; Geijo, M.; Ortíz, J.A.; Fuertes, M.; Ruíz-Fons, J.F.; de la Fuente, J.; Domínguez, L.; et al. The Response of Red Deer to Oral Administration of Heat-Inactivated Mycobacterium Bovis and Challenge with a Field Strain. Vet. Microbiol. 2017, 208, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Gormley, E.; Ní Bhuachalla, D.; O’Keeffe, J.; Murphy, D.; Aldwell, F.E.; Fitzsimons, T.; Stanley, P.; Tratalos, J.A.; McGrath, G.; Fogarty, N.; et al. Oral Vaccination of Free-Living Badgers (Meles meles) with Bacille Calmette Guérin (BCG) Vaccine Confers Protection against Tuberculosis. PLoS ONE 2017, 12, e0168851. [Google Scholar] [CrossRef] [PubMed]
- Walsh, P.D.; Kurup, D.; Hasselschwert, D.L.; Wirblich, C.; Goetzmann, J.E.; Schnell, M.J. The Final (Oral Ebola) Vaccine Trial on Captive Chimpanzees? Sci. Rep. 2017, 7, 43339. [Google Scholar] [CrossRef]
- Benn Felix, J.; Chaki, S.P.; Xu, Y.; Ficht, T.A.; Rice-Ficht, A.C.; Cook, W.E. Protective Antibody Response Following Oral Vaccination with Microencapsulated Bacillus Anthracis Sterne Strain 34F2 Spores. NPJ Vaccines 2020, 5, 59. [Google Scholar] [CrossRef]
- Wang, Y.; He, R.; Winner, H.; Gauduin, M.-C.; Zhang, N.; He, C.; Zhong, G. Induction of Transmucosal Protection by Oral Vaccination with an Attenuated Chlamydia. Infect. Immun. 2023, 91, e0004323. [Google Scholar] [CrossRef] [PubMed]
- Tizard, I.R. Chapter 18—Porcine Vaccines. In Vaccines for Veterinarians; Tizard, I.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 225–242.e1. ISBN 978-0-323-68299-2. [Google Scholar]
- Opriessnig, T.; Mattei, A.A.; Karuppannan, A.K.; Halbur, P.G. Future Perspectives on Swine Viral Vaccines: Where Are We Headed? Porc. Health Manag. 2021, 7, 1. [Google Scholar] [CrossRef]
- Abdelaziz, K.; Helmy, Y.A.; Yitbarek, A.; Hodgins, D.C.; Sharafeldin, T.A.; Selim, M.S.H. Advances in Poultry Vaccines: Leveraging Biotechnology for Improving Vaccine Development, Stability, and Delivery. Vaccines 2024, 12, 134. [Google Scholar] [CrossRef]
- Maresch, C.; Lange, E.; Teifke, J.P.; Fuchs, W.; Klupp, B.; Müller, T.; Mettenleiter, T.C.; Vahlenkamp, T.W. Oral Immunization of Wild Boar and Domestic Pigs with Attenuated Live Vaccine Protects against Pseudorabies Virus Infection. Vet. Microbiol. 2012, 161, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Kunu, W.; Jiwakanon, J.; Porntrakulpipat, S. A Bread-Based Lyophilized C-Strain CSF Virus Vaccine as an Oral Vaccine in Pigs. Transbound. Emerg. Dis. 2019, 66, 1597–1601. [Google Scholar] [CrossRef]
- Hayama, Y.; Sawai, K.; Murato, Y.; Yamaguchi, E.; Kondo, S.; Yamamoto, T. Analysis of Effective Spatial Range of Oral Vaccination against Classical Swine Fever for Wild Boar. Prev. Vet. Med. 2023, 221, 106080. [Google Scholar] [CrossRef]
- Prabakaran, M.; Madhan, S.; Prabhu, N.; Geng, G.Y.; New, R.; Kwang, J. Reverse Micelle-Encapsulated Recombinant Baculovirus as an Oral Vaccine against H5N1 Infection in Mice. Antiviral. Res. 2010, 86, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Jazayeri, S.D.; Ideris, A.; Zakaria, Z.; Shameli, K.; Moeini, H.; Omar, A.R. Cytotoxicity and Immunological Responses Following Oral Vaccination of Nanoencapsulated Avian Influenza Virus H5 DNA Vaccine with Green Synthesis Silver Nanoparticles. J. Control Release 2012, 161, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.; Jiang, X.; Yang, Z.; Ren, X.; Gong, H.; Reeves, M.; Sheng, J.; Wang, Y.; Pan, Z.; Liu, F.; et al. Oral Delivery of a Novel Attenuated Salmonella Vaccine Expressing Influenza A Virus Proteins Protects Mice against H5N1 and H1N1 Viral Infection. PLoS ONE 2015, 10, e0129276. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Xie, B.; Gao, T.; Cen, Q.; Ren, Y. Yeast Display Platform Technology to Prepare Oral Vaccine against Lethal H7N9 Virus Challenge in Mice. Microb. Cell Fact. 2020, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Lu, X.; Li, S.; Ren, Y. High Immune Efficacy against Different Avian Influenza H5N1 Viruses Due to Oral Administration of a Saccharomyces Cerevisiae-Based Vaccine in Chickens. Sci. Rep. 2021, 11, 8977. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Lu, X.; Yang, Z.; Lei, H. Protective Immunity Induced by Oral Vaccination with a Recombinant Lactococcus Lactis Vaccine against H5Nx in Chickens. BMC Vet. Res. 2022, 18, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Z.; Xu, H.; Xiang, B.; Dang, R.; Yang, Z. Orally Administrated Whole Yeast Vaccine Against Porcine Epidemic Diarrhea Virus Induced High Levels of IgA Response in Mice and Piglets. Viral. Immunol. 2016, 29, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, L.; Mou, C.; Zhang, E.; Wang, Y.; Cao, Y.; Yang, Q. Mucosal Immune Responses Induced by Oral Administration Recombinant Bacillus Subtilis Expressing the COE Antigen of PEDV in Newborn Piglets. Biosci. Rep. 2019, 39, BSR20182028. [Google Scholar] [CrossRef] [PubMed]
- Do, V.T.; Jang, J.; Park, J.; Dao, H.T.; Kim, K.; Hahn, T.-W. Recombinant Adenovirus Carrying a Core Neutralizing Epitope of Porcine Epidemic Diarrhea Virus and Heat-Labile Enterotoxin B of E. coli as a Mucosal Vaccine. Arch. Virol. 2020, 165, 609–618. [Google Scholar] [CrossRef]
- Tien, N.-Q.-D.; Yang, M.-S.; Jang, Y.-S.; Kwon, T.-H.; Reljic, R.; Kim, M.-Y. Systemic and Oral Immunogenicity of Porcine Epidemic Diarrhea Virus Antigen Fused to Poly-Fc of Immunoglobulin G and Expressed in ΔXT/FT Nicotiana Benthamiana Plants. Front. Pharmacol. 2021, 12, 653064. [Google Scholar] [CrossRef]
- Mebrahtu, K.; Teshale, S.; Esatu, W.; Habte, T.; Gelaye, E. Evaluation of Spray and Oral Delivery of Newcastle Disease I2 Vaccine in Chicken Reared by Smallholder Farmers in Central Ethiopia. BMC Vet. Res. 2018, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Katalani, C.; Ahmadian, G.; Nematzadeh, G.; Amani, J.; Ehsani, P.; Razmyar, J.; Kiani, G. Immunization with Oral and Parenteral Subunit Chimeric Vaccine Candidate Confers Protection against Necrotic Enteritis in Chickens. Vaccine 2020, 38, 7284–7291. [Google Scholar] [CrossRef] [PubMed]
- Shamshirgaran, M.A.; Golchin, M.; Salehi, M.; Kheirandish, R. Evaluation the Efficacy of Oral Immunization of Broiler Chickens with a Recombinant Lactobacillus Casei Vaccine Vector Expressing the Carboxy-Terminal Fragment of α-Toxin from Clostridium Perfringens. BMC Vet. Res. 2023, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jia, R.; Shen, H.; Wang, M.; Zhu, D.; Chen, S.; Liu, M.; Zhao, X.; Wu, Y.; Yang, Q.; et al. Oral Delivery of a DNA Vaccine Expressing the PrM and E Genes: A Promising Vaccine Strategy against Flavivirus in Ducks. Sci. Rep. 2018, 8, 12360. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Shen, H.; Jia, R.; Wang, M.; Chen, S.; Zhu, D.; Liu, M.; Zhao, X.; Yang, Q.; Wu, Y.; et al. Oral Vaccination with a DNA Vaccine Encoding Capsid Protein of Duck Tembusu Virus Induces Protection Immunity. Viruses 2018, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Geng, Y.; Wang, K.; Yu, Z.; Yang, P.O.; Yang, Z.; He, C.; Huang, C.; Yin, L.; He, M.; et al. Adjuvant Effects of Interleukin-2 Co-Expression with VP60 in an Oral Vaccine Delivered by Attenuated Salmonella Typhimurium against Rabbit Hemorrhagic Disease. Vet. Microbiol. 2019, 230, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Shuai, L.; Ge, J.; Wen, Z.; Wang, J.; Wang, X.; Bu, Z. Immune Responses in Mice and Pigs after Oral Vaccination with Rabies Virus Vectored Nipah Disease Vaccines. Vet. Microbiol. 2020, 241, 108549. [Google Scholar] [CrossRef]
- Liu, X.; Qi, L.; Lv, J.; Zhang, Z.; Zhou, P.; Ma, Z.; Wang, Y.; Zhang, Y.; Pan, L. The Immune Response to a Recombinant Lactococcus Lactis Oral Vaccine against Foot-and-Mouth Disease Virus in Mice. Biotechnol. Lett. 2020, 42, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Hewawaduge, C.; Senevirathne, A.; Lee, J.H. Enhancement of Host Infectivity, Immunity, and Protective Efficacy by Addition of Sodium Bicarbonate Antacid to Oral Vaccine Formulation of Live Attenuated Salmonella Secreting Brucella Antigens. Microb. Pathog. 2020, 138, 103857. [Google Scholar] [CrossRef]
- Samakchan, N.; Thinwang, P.; Boonyom, R. Oral Immunization of Rat with Chromosomal Expression LipL32 in Attenuated Salmonella Vaccine Induces Immune Respond against Pathogenic Leptospira. Clin. Exp. Vaccine Res. 2021, 10, 217–228. [Google Scholar] [CrossRef]
- Wang, N.; Wang, J.-Y.; Pan, T.-X.; Jiang, Y.-L.; Huang, H.-B.; Yang, W.-T.; Shi, C.-W.; Wang, J.-Z.; Wang, D.; Zhao, D.-D.; et al. Oral Vaccination with Attenuated Salmonella Encoding the Trichinella Spiralis 43-kDa Protein Elicits Protective Immunity in BALB/c Mice. Acta Trop. 2021, 222, 106071. [Google Scholar] [CrossRef] [PubMed]
- Mechler-Dreibi, M.L.; Almeida, H.M.S.; Sonalio, K.; Martines, M.A.C.; Petri, F.A.M.; Zambotti, B.B.; Ferreira, M.M.; Storino, G.Y.; Martins, T.S.; Montassier, H.J.; et al. Oral Vaccination of Piglets against Mycoplasma Hyopneumoniae Using Silica SBA-15 as an Adjuvant Effectively Reduced Consolidation Lung Lesions at Slaughter. Sci. Rep. 2021, 11, 22377. [Google Scholar] [CrossRef] [PubMed]
- Lucero, M.S.; Chimeno Zoth, S.; Jaton, J.; Gravisaco, M.J.; Pinto, S.; Richetta, M.; Berinstein, A.; Gómez, E. Oral Immunization With Plant-Based Vaccine Induces a Protective Response Against Infectious Bursal Disease. Front. Plant Sci. 2021, 12, 741469. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hua, D.; Qu, Q.; Cao, H.; Feng, Z.; Liu, N.; Huang, J.; Zhang, L. Oral Immunization with Recombinant Saccharomyces Cerevisiae Expressing Viral Capsid Protein 2 of Infectious Bursal Disease Virus Induces Unique Specific Antibodies and Protective Immunity. Vaccines 2023, 11, 1849. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Luo, P.; Chen, Y.; Xu, J.; Sun, J.; Guan, C.; Wang, P.; Chen, M.; Zhang, X.; Zhu, Y.; et al. Regulated Delayed Attenuation Improves Vaccine Efficacy in Preventing Infection from Avian Pathogenic E. coli O78 and Salmonella Typhimurium. Vet. Microbiol. 2021, 254, 109012. [Google Scholar] [CrossRef] [PubMed]
- Shringi, S.; Sheng, H.; Potter, A.A.; Minnich, S.A.; Hovde, C.J.; Besser, T.E. Repeated Oral Vaccination of Cattle with Shiga Toxin-Negative E. coli O157:H7 Reduces Carriage of Wild-Type E. coli O157:H7 after Challenge. Appl. Environ. Microbiol. 2021, 87, e02183-20. [Google Scholar] [CrossRef]
- Van der Weken, H.; Jahantigh, H.R.; Cox, E.; Devriendt, B. Targeted Delivery of Oral Vaccine Antigens to Aminopeptidase N Protects Pigs against Pathogenic E. coli Challenge Infection. Front. Immunol. 2023, 14, 1192715. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Y.; Lin, S.M.; Jang, A.-Y.; Ahn, K.B.; Ji, H.J.; Guo, H.-C.; Lim, S.; Seo, H.S. Effective Mucosal Live Attenuated Salmonella Vaccine by Deleting Phosphotransferase System Component Genes ptsI and Crr. J. Microbiol. 2019, 57, 64–73. [Google Scholar] [CrossRef]
- Zafar, H.; Rahman, S.U.; Ali, S.; Javed, M.T. Evaluation of a Salmonella Strain Isolated from Honeybee Gut as a Potential Live Oral Vaccine Against Lethal Infection of Salmonella Typhimurium. Pol. J. Microbiol. 2019, 68, 173–183. [Google Scholar] [CrossRef]
- Renu, S.; Han, Y.; Dhakal, S.; Lakshmanappa, Y.S.; Ghimire, S.; Feliciano-Ruiz, N.; Senapati, S.; Narasimhan, B.; Selvaraj, R.; Renukaradhya, G.J. Chitosan-Adjuvanted Salmonella Subunit Nanoparticle Vaccine for Poultry Delivered through Drinking Water and Feed. Carbohydr. Polym. 2020, 243, 116434. [Google Scholar] [CrossRef]
- Higginson, E.E.; Panda, A.; Toapanta, F.R.; Terzi, M.C.; Jones, J.A.; Sen, S.; Permala-Booth, J.; Pasetti, M.F.; Sztein, M.B.; DeTolla, L.; et al. Immunogenicity and Efficacy of Live-Attenuated Salmonella Typhimurium Vaccine Candidate CVD 1926 in a Rhesus Macaque Model of Gastroenteritis. Infect. Immun. 2021, 89, e0008721. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Repáraz, J.; Sebastià, E.; Sitjà, M.; Tamayo, I.; Irache, J.M.; Gamazo, C. Protection Conferred by Drinking Water Administration of a Nanoparticle-Based Vaccine against Salmonella Enteritidis in Hens. Vaccines 2021, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Wu, H.-C.; Ding, H.-C.; Li, S.-J.; Bao, E.-D.; Yang, B.-S.; Li, Y.-J.; Gao, X.-L.; Duan, Q.; Zhu, G.-Q. Safety and Protective Effects of an Avirulent Salmonella Gallinarum Isolate as a Vaccine Candidate against Salmonella Gallinarum Infections in Young Chickens. Vet. Immunol. Immunopathol. 2022, 253, 110501. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; La, T.-M.; Lee, H.-J.; Kim, T.; Song, S.-U.; Park, E.; Park, G.-H.; Choi, I.-S.; Park, S.-Y.; Lee, J.-B.; et al. Evaluation of Immune Responses and Protective Efficacy of a Novel Live Attenuated Salmonella Enteritidis Vaccine Candidate in Chickens. Vaccines 2022, 10, 1405. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, H.; Pan, N.; Liu, Y.; Sheng, S.; Li, H.; Bao, X.; Wang, X. Oral Administration with Recombinant Attenuated Regulated Delayed Lysis Salmonella Vaccines Protecting against Staphylococcus Aureus Kidney Abscess Formation. Vaccines 2022, 10, 1073. [Google Scholar] [CrossRef]
- Kaldis, A.; Uddin, M.S.; Guluarte, J.O.; Martin, C.; Alexander, T.W.; Menassa, R. Development of a Plant-Based Oral Vaccine Candidate against the Bovine Respiratory Pathogen Mannheimia Haemolytica. Front. Plant Sci. 2023, 14, 1251046. [Google Scholar] [CrossRef] [PubMed]
- Soutter, F.; Werling, D.; Nolan, M.; Küster, T.; Attree, E.; Marugán-Hernández, V.; Kim, S.; Tomley, F.M.; Blake, D.P. A Novel Whole Yeast-Based Subunit Oral Vaccine Against Eimeria Tenella in Chickens. Front. Immunol. 2022, 13, 809711. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Qu, H.; Zheng, W.; Zhang, Y.; Li, Y.; Pan, T.; Li, J.; Yang, W.; Cao, X.; Jiang, Y.; et al. Oral Vaccination with a Recombinant Lactobacillus plantarum Expressing the Eimeria Tenella Rhoptry Neck 2 Protein Elicits Protective Immunity in Broiler Chickens Infected with Eimeria Tenella. Parasit. Vectors. 2024, 17, 277. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Chang, W.-C.; Wu, M.-C.; Liaw, J.; Shiau, A.-L.; Chu, C.-Y. Oral DNA Vaccine Adjuvanted with Cyclic Peptide Nanotubes Induced a Virus-Specific Antibody Response in Ducklings against Goose Parvovirus. Vet. Q. 2023, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Eshraghisamani, R.; Facciuolo, A.; Harman-McKenna, V.; Illanes, O.; De Buck, J. Immunogenicity and Efficacy of an Oral Live-Attenuated Vaccine for Bovine Johne’s Disease. Front. Immunol. 2023, 14, 1307621. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.-E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and Future Perspectives of Vaccines for Industrialised Fin-Fish Farming. Fish Shellfish Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Embregts, C.W.E.; Forlenza, M. Oral Vaccination of Fish: Lessons from Humans and Veterinary Species. Dev. Comp. Immunol. 2016, 64, 118–137. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.K.; Manna, S.K.; Thomas Allnutt, F.C. Viral Vaccines for Farmed Finfish. Virusdisease 2014, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sommerset, I.; Krossøy, B.; Biering, E.; Frost, P. Vaccines for Fish in Aquaculture. Expert Rev. Vaccines 2005, 4, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Hu, X.; Miao, L.; Chen, J. Current Status and Development Prospects of Aquatic Vaccines. Front. Immunol. 2022, 13, 1040336. [Google Scholar] [CrossRef] [PubMed]
- Mutoloki, S.; Munang’andu, H.M.; Evensen, Ø. Oral Vaccination of Fish—Antigen Preparations, Uptake, and Immune Induction. Front. Immunol. 2015, 6, 519. [Google Scholar] [CrossRef]
- Anuradha, K.; Foo, H.L.; Mariana, N.S.; Loh, T.C.; Yusoff, K.; Hassan, M.D.; Sasan, H.; Raha, A.R. Live Recombinant Lactococcus Lactis Vaccine Expressing Aerolysin Genes D1 and D4 for Protection against Aeromonas Hydrophila in Tilapia (Oreochromis niloticus). J. Appl. Microbiol. 2010, 109, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Li, L.P.; Wang, R.; Liang, W.W.; Huang, T.; Huang, Y.; Luo, F.G.; Lei, A.Y.; Chen, M.; Gan, X. Development of Live Attenuated Streptococcus Agalactiae Vaccine for Tilapia via Continuous Passage in Vitro. Fish Shellfish Immunol. 2015, 45, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Megat Mazhar Khair, M.H.; Song, A.A.L.; Masarudin, M.J.; Loh, J.Y.; Chong, C.M.; Beardall, J.; Teo, M.Y.M.; In, L.L.A. Recombinant Lactococcal-Based Oral Vaccine for Protection against Streptococcus Agalactiae Infections in Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2024, 149, 109572. [Google Scholar] [CrossRef]
- Wise, D.J.; Greenway, T.E.; Byars, T.S.; Griffin, M.J.; Khoo, L.H. Oral Vaccination of Channel Catfish against Enteric Septicemia of Catfish Using a Live Attenuated Edwardsiella Ictaluri Isolate. J. Aquat. Anim. Health 2015, 27, 135–143. [Google Scholar] [CrossRef]
- Nho, S.W.; Abdelhamed, H.; Karsi, A.; Lawrence, M.L. Improving Safety of a Live Attenuated Edwardsiella Ictaluri Vaccine against Enteric Septicemia of Catfish and Evaluation of Efficacy. Vet. Microbiol. 2017, 210, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Hua, X.; Wang, Y.; Wang, Y.; Chen, Y.; Shi, W.; Tang, L.; Li, Y.; Liu, M. Oral Immunization with a Recombinant Lactobacillus Expressing CK6 Fused with VP2 Protein against IPNV in Rainbow Trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2018, 83, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hua, X.; Ren, X.; Duan, K.; Gao, S.; Sun, J.; Feng, Y.; Zhou, Y.; Guan, X.; Li, D.; et al. Oral Immunization with Recombinant Lactobacillus Casei Displayed AHA1-CK6 and VP2 Induces Protection against Infectious Pancreatic Necrosis in Rainbow Trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2020, 100, 18–26. [Google Scholar] [CrossRef]
- Seo, J.Y.; Chung, H.J.; Kim, T.J. Codon-Optimized Expression of Fish Iridovirus Capsid Protein in Yeast and Its Application as an Oral Vaccine Candidate. J. Fish Dis. 2013, 36, 763–768. [Google Scholar] [CrossRef]
- Shin, Y.J.; Kwon, T.H.; Seo, J.Y.; Kim, T.J. Oral Immunization of Fish against Iridovirus Infection Using Recombinant Antigen Produced from Rice Callus. Vaccine 2013, 31, 5210–5215. [Google Scholar] [CrossRef]
- Wi, G.R.; Hwang, J.Y.; Kwon, M.-G.; Kim, H.J.; Kang, H.A.; Kim, H.-J. Protective Immunity against Nervous Necrosis Virus in Convict Grouper Epinephelus Septemfasciatus Following Vaccination with Virus-like Particles Produced in Yeast Saccharomyces Cerevisiae. Vet. Microbiol. 2015, 177, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, N.A.; Rodriguez Saint-Jean, S.; Perez-Prieto, S.I. Immune Responses to Oral pcDNA-VP2 Vaccine in Relation to Infectious Pancreatic Necrosis Virus Carrier State in Rainbow Trout Oncorhynchus mykiss. Vet. Immunol. Immunopathol. 2015, 165, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Adomako, M.; St-Hilaire, S.; Zheng, Y.; Eley, J.; Marcum, R.D.; Sealey, W.; Donahower, B.C.; Lapatra, S.; Sheridan, P.P. Oral DNA Vaccination of Rainbow Trout, Oncorhynchus mykiss (Walbaum), against Infectious Haematopoietic Necrosis Virus Using PLGA [Poly(D,L-Lactic-Co-Glycolic Acid)] Nanoparticles. J. Fish Dis. 2012, 35, 203–214. [Google Scholar] [CrossRef]
- Ballesteros, N.A.; Alonso, M.; Saint-Jean, S.R.; Perez-Prieto, S.I. An Oral DNA Vaccine against Infectious Haematopoietic Necrosis Virus (IHNV) Encapsulated in Alginate Microspheres Induces Dose-Dependent Immune Responses and Significant Protection in Rainbow Trout (Oncorrhynchus Mykiss). Fish Shellfish Immunol. 2015, 45, 877–888. [Google Scholar] [CrossRef]
- Smreczak, M.; Orłowska, A.; Müller, T.; Freuling, C.M.; Kawiak-Sadurska, M.; Trębas, P. Vaccine-Induced Rabies in a Red Fox in Poland. J. Vet. Res. 2022, 66, 473–477. [Google Scholar] [CrossRef]
- Tsai, C.J.Y.; Loh, J.M.S.; Fujihashi, K.; Kiyono, H. Mucosal Vaccination: Onward and Upward. Expert Rev. Vaccines 2023, 22, 885–899. [Google Scholar] [CrossRef] [PubMed]
| Types of Oral Vaccines | Advantage | Disadvantage |
|---|---|---|
| Oral attenuated vaccine | Affordable and accessible, high immunogenicity | Virulence reversion risk, occasional side effects, low stability |
| Recombinant vectored oral vaccine | Good scalability, Multivalency [13] | Potential safety risks, pre-existing immunity (e.g., adenovirus vector) [13,14] |
| Nanoparticle oral vaccine | Control release, effect of adjuvants, biodegradable, can be designed rationally [15] | Complex production process, some potential risks of side effects and may transferred from biological barriers in animals [15] |
| Transgenic plant oral vaccine | Cost-effective and can be mass-produced in a green way [16], prolong the residence time on the mucosa [17], stable at room temperature [18] | Low expression level, risk of transgene contamination via pollen or seeds, need for oral priming with adjuvants [18] |
| Pathogen | Trial Animals | Type | Protection Rate | Strain/Vector | Antigen | References |
|---|---|---|---|---|---|---|
| Aeromonas hydrophila | Tilapia | Recombinant vector | 70–100% | Lactococcus lactis | D1, D4 | [159] |
| Streptococcus agalactiae | Nile tilapia | Attenuated | 50–85% | YM001 | - | [160] |
| Tilapia | Recombinant vector | 50–89% | Lactococcus lactis | SIP | [161] | |
| Edwardsiella ictaluri | Channel Catfish | Attenuated | 54–100% | S97-773 | - | [162] |
| Catfish | Attenuated | 81–94% | NDKL1 | - | [163] | |
| Infectious pancreatic necrosis virus | Rainbow trout | Recombinant vector | - | Lactobacillus casei | CK6, VP2 | [164] |
| Rainbow trout | Recombinant vector | - | Lactobacillus casei | AHA1, CK6, VP2 | [165] | |
| Rock bream iridovirus | Rock bream | Recombinant vector | - | Yeast | Major capsid protein | [166] |
| Rock bream | Plant-based | - | Rice callus | Major capsid protein | [167] | |
| Nervous necrosis virus | Convict grouper | Nanoparticle | - | VLPs | - | [168] |
| Infectious pancreatic virus | Rainbow trout | Nanoparticle | 80% | Alginates | VP2 DNA | [169] |
| Infectious hematopoietic necrosis virus | Rainbow trout | Nanoparticle | 17–33% | PLGA | Glycoprotein DNA | [170] |
| Rainbow trout | Nanoparticle | 21–56% | Alginates | Glycoprotein DNA | [171] |
| Pathogen | Name of Medicine | Type | Company | Species |
|---|---|---|---|---|
| Bordetella bronchiseptica, Canine parainfluenza virus | Nobivac® Intra-Trac® Oral BbPi | Attenuated | Merck animal health | Canine |
| Bordetella bronchiseptica | TruCan B (Oral) | Attenuated | Elanco Animal Health | Canine |
| Rabies virus | Rabitec | Attenuated | CEVA Santé Animale | Foxes, raccoon dogs |
| Rabigen SAG2 | Attenuated | Virbac S.A. | Foxes, raccoon dogs | |
| Rabidog | Attenuated | Virbac S.A. | Canine | |
| RABORAL V-RG® | Recombinant vector | Boehringer Ingelheim | Raccoons, coyotes | |
| LYSVULPEN | Attenuated | Bioveta A.S | Foxes, raccoon dogs | |
| Salmonella typhimurium | POULVAC® ST | Attenuated | Zoetis | Poultry |
| Rotavirus | PROSYSTEM® ROTA | Attenuated | Merck animal health | Swine |
| Erysipelothrix rhusiopathiae | ERY VAC 100 | Attenuated | Arko Laboratories | Swine |
| Lawsonia intracellularis | Enterisol Ileitis | Attenuated | Boehringer Ingelheim | Swine |
| Rotavirus, coronavirus | Calf-Guard | Attenuated | Zoetis Animal Health | Calves |
| Yersinia ruckeri | AQUAVAC® ERM Oral | Inactivated | Merck animal health | Rainbow trout |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, K.; Chen, X.; Zhang, J.; Jiang, X.; Zhang, J.; Huang, M.; Bi, S.; Ju, C.; Luo, Y. Recent Advances in Oral Vaccines for Animals. Vet. Sci. 2024, 11, 353. https://doi.org/10.3390/vetsci11080353
Zhong K, Chen X, Zhang J, Jiang X, Zhang J, Huang M, Bi S, Ju C, Luo Y. Recent Advances in Oral Vaccines for Animals. Veterinary Sciences. 2024; 11(8):353. https://doi.org/10.3390/vetsci11080353
Chicago/Turabian StyleZhong, Kaining, Xinting Chen, Junhao Zhang, Xiaoyu Jiang, Junhui Zhang, Minyi Huang, Shuilian Bi, Chunmei Ju, and Yongwen Luo. 2024. "Recent Advances in Oral Vaccines for Animals" Veterinary Sciences 11, no. 8: 353. https://doi.org/10.3390/vetsci11080353