

Ameliorative Effect of Lycopene on Follicular Reserve Depletion, Oxidative Damage, Apoptosis Rate, and Hormonal Profile during Repeated Superovulations in Mice


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Experimental Design
2.4. Animals’ Euthanasia, Blood, and Tissue Sampling
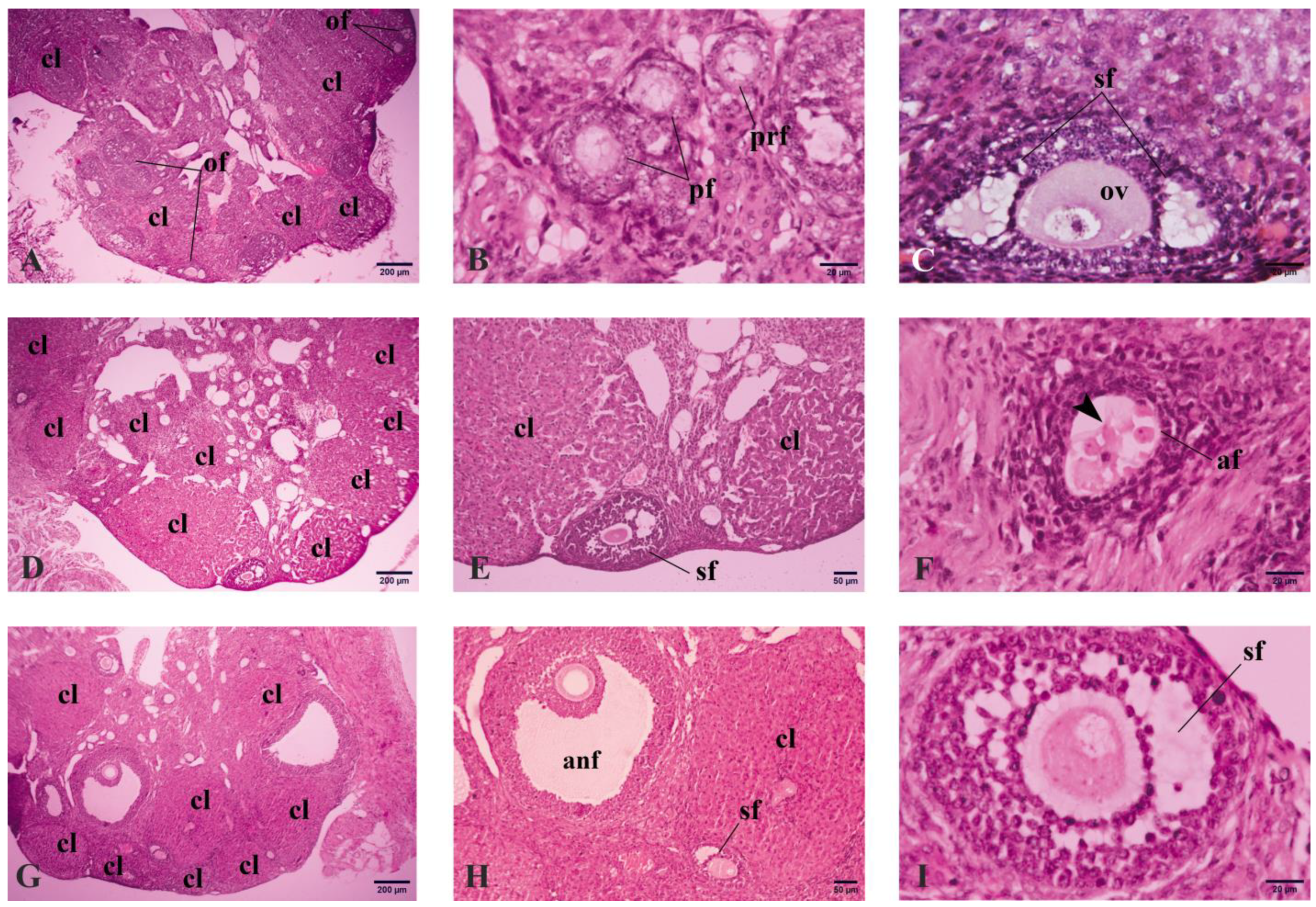
2.5. Histological and Immunohistochemical Analyses
2.6. Biochemical Assessment of the Ovarian Redox Status
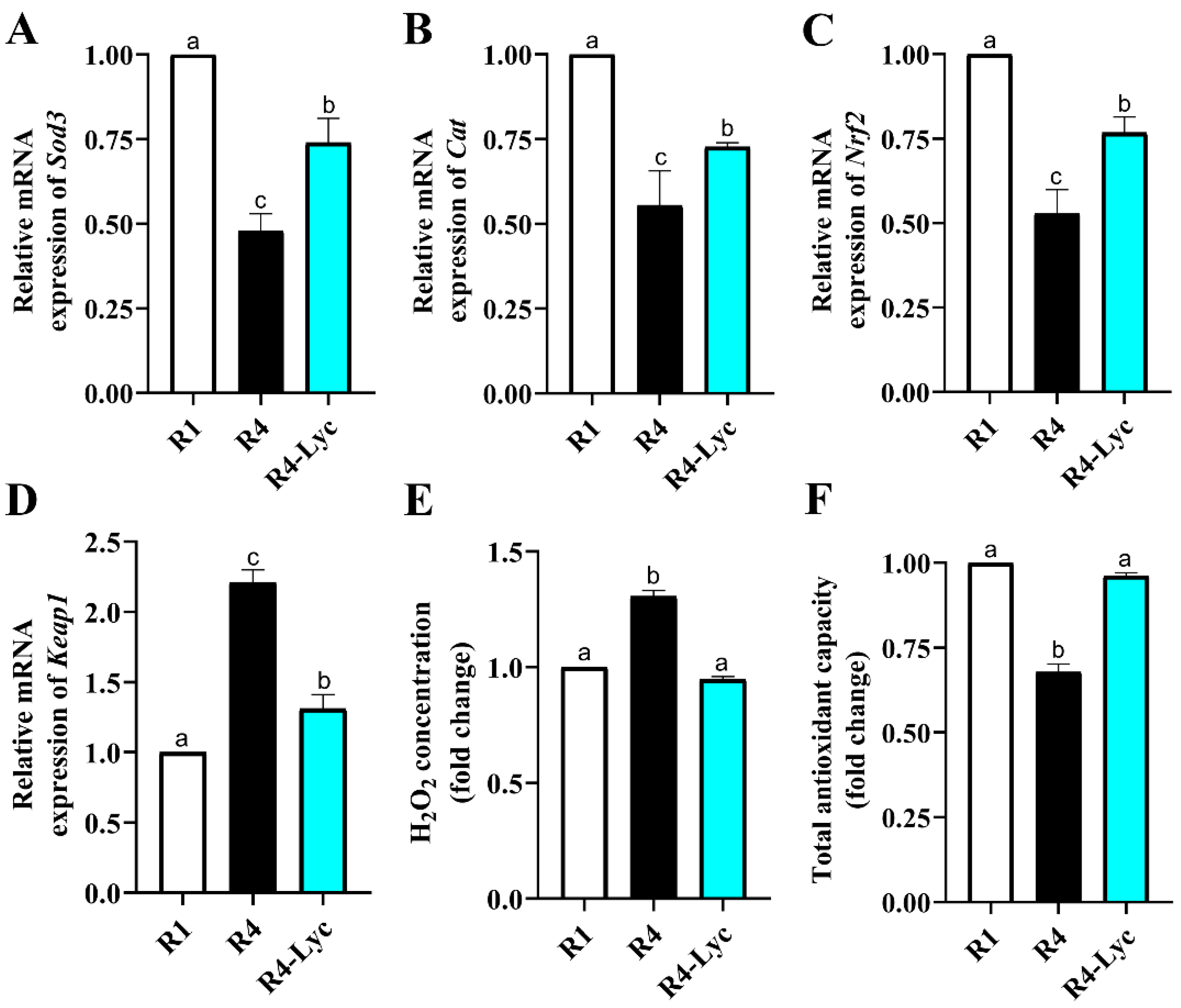
2.7. Gene Expression Analysis
2.8. Serum Hormone Levels Analysis
2.9. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duffy, D.M.; Ko, C.; Jo, M.; Brannstrom, M.; Curry, T.E., Jr. Ovulation: Parallels with inflammatory processes. Endocr. Rev. 2019, 40, 369–416. [Google Scholar] [CrossRef] [PubMed]
- Zaniker, E.J.; Babayev, E.; Duncan, F.E. Common mechanisms of physiological and pathological rupture events in biology: Novel insights into mammalian ovulation and beyond. Biol. Rev. 2023, 98, 1648–1667. [Google Scholar] [CrossRef] [PubMed]
- Lliberos, C.; Liew, S.H.; Mansell, A.; Hutt, K.J. The Inflammasome Contributes to Depletion of the Ovarian Reserve During Aging in Mice. Front. Cell Dev. Biol. 2020, 8, 628473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lai, Z.; Shi, L.; Tian, Y.; Luo, A.; Xu, Z.; Ma, X.; Wang, S. Repeated superovulation increases the risk of osteoporosis and cardiovascular diseases by accelerating ovarian aging in mice. Aging 2018, 10, 1089–1102. [Google Scholar] [CrossRef]
- Miyamoto, K.; Sato, E.F.; Kasahara, E.; Jikumaru, M.; Hiramoto, K.; Tabata, H.; Katsuragi, M.; Odo, S.; Utsumi, K.; Inoue, M. Effect of oxidative stress during repeated ovulation on the structure and functions of the ovary, oocytes, and their mitochondria. Free Radic. Biol. Med. 2010, 49, 674–681. [Google Scholar] [CrossRef]
- Chao, H.T.; Lee, S.Y.; Lee, H.M.; Liao, T.L.; Wei, Y.H.; Kao, S.H. Repeated ovarian stimulations induce oxidative damage and mitochondrial DNA mutations in mouse ovaries. Ann. N. Y. Acad. Sci. 2005, 1042, 148–156. [Google Scholar] [CrossRef]
- Cortell, C.; Salvetti, P.; Joly, T.; Viudes-de-Castro, M.P. Effect of different superovulation stimulation protocols on adenosine triphosphate concentration in rabbit oocytes. Zygote 2015, 23, 507–513. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, T.S.; Kim, J.M.; Chang, K.T.; Lee, H.S.; Lee, D.S. Repeated Superovulation via PMSG/hCG Administration Induces 2-Cys Peroxiredoxins Expression and Overoxidation in the Reproductive Tracts of Female Mice. Mol. Cells 2015, 38, 1071–1078. [Google Scholar] [CrossRef]
- Kalthur, G.; Salian, S.R.; Nair, R.; Mathew, J.; Adiga, S.K.; Kalthur, S.G.; Zeegers, D.; Hande, M.P. Distribution pattern of cytoplasmic organelles, spindle integrity, oxidative stress, octamer-binding transcription factor 4 (Oct4) expression and developmental potential of oocytes following multiple superovulation. Reprod. Fertil. Dev. 2016, 28, 2027–2038. [Google Scholar] [CrossRef]
- Xie, J.K.; Wang, Q.; Zhang, T.T.; Yin, S.; Zhang, C.L.; Ge, Z.J. Repeated superovulation may affect mitochondrial functions of cumulus cells in mice. Sci. Rep. 2016, 6, 31368. [Google Scholar] [CrossRef]
- Xiao, P.; Nie, J.; Wang, X.; Lu, K.; Lu, S.; Liang, X. Melatonin alleviates the deterioration of oocytes from mice subjected to repeated superovulation. J. Cell. Physiol. 2019, 234, 13413–13422. [Google Scholar] [CrossRef]
- Liu, B.; Wang, J.L.; Wang, X.M.; Zhang, C.; Dai, J.G.; Huang, X.M.; Gao, J.M. Reparative effects of lycium barbarum polysaccharide on mouse ovarian injuries induced by repeated superovulation. Theriogenology 2020, 145, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Lugasi, A.; Bíró, L.; Hóvárie, J.; Sági, K.V.; Brandt, S.; Barna, É. Lycopene content of foods and lycopene intake in two groups of the Hungarian population. Nutr. Res. 2003, 23, 1035–1044. [Google Scholar] [CrossRef]
- Wenli, Y.; Yaping, Z.; Zhen, X.; Hui, J.; Dapu, W. The antioxidant properties of lycopene concentrate extracted from tomato paste. J. Am. Oil Chem. Soc. 2001, 78, 697–701. [Google Scholar] [CrossRef]
- Miller, N.J.; Sampson, J.; Candeias, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef] [PubMed]
- Rakha, S.I.; Elmetwally, M.A.; El-Sheikh Ali, H.; Balboula, A.Z.; Mahmoud, A.M.; Zaabel, S.M. Lycopene Reduces the In Vitro Aging Phenotypes of Mouse Oocytes by Improving Their Oxidative Status. Vet. Sci. 2022, 9, 336. [Google Scholar] [CrossRef]
- Rakha, S.I.; Elmetwally, M.A.; El-Sheikh Ali, H.; Balboula, A.Z.; Mahmoud, A.M.; Zaabel, S.M. Lycopene Improves Maturation Rate and Antioxidant Status of In Vitro Matured Mouse Oocytes. Int. J. Vet. Sci. 2022, 12, 248–254. [Google Scholar]
- Ji, X.; Ye, Y.; Wang, L.; Liu, S.; Dong, X. Association between nutrient intake and female infertility: A study based on NHANES database. J. Obstet. Gynaecol. 2023, 43, 2285025. [Google Scholar] [CrossRef]
- Kulhan, N.G.; Kulhan, M.; Turkler, C.; Ata, N.; Kiremitli, T.; Kiremitli, S.; Keskin Cimen, F.; Suleyman, H.; Toprak, V. Effect of lycopene on oxidative ovary-damage induced by cisplatin in rats. Gen. Physiol. Biophys. 2019, 38, 253–258. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Kumar, A. Neuroprotective Effect of Lycopene Against PTZ-induced Kindling Seizures in Mice: Possible Behavioural, Biochemical and Mitochondrial Dysfunction. Phytother. Res. 2016, 30, 306–313. [Google Scholar] [CrossRef]
- Devaraj, S.; Mathur, S.; Basu, A.; Aung, H.H.; Vasu, V.T.; Meyers, S.; Jialal, I. A dose-response study on the effects of purified lycopene supplementation on biomarkers of oxidative stress. J. Am. Coll. Nutr. 2008, 27, 267–273. [Google Scholar] [CrossRef]
- Stangl, V.; Kuhn, C.; Hentschel, S.; Jochmann, N.; Jacob, C.; Böhm, V.; Fröhlich, K.; Müller, L.; Gericke, C.; Lorenz, M. Lack of effects of tomato products on endothelial function in human subjects: Results of a randomised, placebo-controlled cross-over study. Br. J. Nutr. 2011, 105, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Abdellatif, A.M. Structure of the Eurasian moorhen spleen: A comprehensive study using gross anatomy, light, and transmission electron microscopy. Microsc. Res. Tech. 2021, 84, 1696–1709. [Google Scholar] [CrossRef]
- Abdellatif, A.M.; Farag, A.; Metwally, E. Anatomical, histochemical, and immunohistochemical observations on the gastrointestinal tract of Gallinula chloropus (Aves: Rallidae). BMC Zool. 2022, 7, 61. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, S.X.; He, J.N.; Zhao, H.; Gu, B.X.; Xie, J.K.; Zhao, Y.J.; Zhang, C.L.; Ge, Z.J. Repeated Superovulation Accelerates Primordial Follicle Activation and Atresia. Cells 2022, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Torrejón, R.; Bández, M.J.; López-Cepero, J.M.; Boveris, A. Mitochondrial function and mitochondria-induced apoptosis in an overstimulated rat ovarian cycle. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E1101–E1109. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Guo, Y.; Cao, H.; Zhou, T.; Zhou, Z.; Sha, J.; Guo, X.; Zhu, H. Long-term effects of repeated superovulation on ovarian structure and function in rhesus monkeys. Fertil. Steril. 2014, 102, 1452–1457. [Google Scholar] [CrossRef]
- Nie, X.; Dai, Y.; Zheng, Y.; Bao, D.; Chen, Q.; Yin, Y.; Fu, H.; Hou, D. Establishment of a Mouse Model of Premature Ovarian Failure Using Consecutive Superovulation. Cell. Physiol. Biochem. 2018, 51, 2341–2358. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, Y.; Miao, C.; Wang, R.; Yang, L.; Liu, J.; Chen, Z.; Zhang, Q.; Ma, J. He’s Yangchao Recipe Ameliorates Ovarian Oxidative Stress of Aging Mice under Consecutive Superovulation Involving JNK- and P53-Related Mechanism. Evid. Based Complement. Altern. Med. 2022, 2022, 7705194. [Google Scholar] [CrossRef]
- Feng, Y.; Wu, Z.; Zhao, X.; Chen, M.; Li, S.; Lu, C.; Shi, D.; Lu, F. Epicatechin promotes oocyte quality in mice during repeated superovulation. Theriogenology 2023, 209, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Takahashi, T.; Amita, M.; Matsuo, K.; Igarashi, H.; Kurachi, H. Pioglitazone counteracts the tumor necrosis factor-α inhibition of follicle-stimulating hormone-induced follicular development and estradiol production in an in vitro mouse preantral follicle culture system. J. Ovarian Res. 2013, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Mainardi, G.L.; Bussolati, S.; Tamanini, C. Steroidogenesis, proliferation and apoptosis in bovine granulosa cells: Role of tumour necrosis factor-alpha and its possible signalling mechanisms. Reprod. Fertil. Dev. 2002, 14, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Hu, H. Effects of lycopene on ovarian cancer cell line SKOV3 in vitro: Suppressed proliferation and enhanced apoptosis. Mol. Cell. Probes 2019, 46, 101419. [Google Scholar] [CrossRef]
- Sahin, K.; Yenice, E.; Tuzcu, M.; Orhan, C.; Mizrak, C.; Ozercan, I.H.; Sahin, N.; Yilmaz, B.; Bilir, B.; Ozpolat, B.; et al. Lycopene Protects Against Spontaneous Ovarian Cancer Formation in Laying Hens. J. Cancer Prev. 2018, 23, 25–36. [Google Scholar] [CrossRef]
- Pierine, D.T.; Navarro, M.E.L.; Minatel, I.O.; Luvizotto, R.A.M.; Nascimento, A.F.; Ferreira, A.L.A.; Yeum, K.J.; Corrêa, C.R. Lycopene supplementation reduces TNF-α via RAGE in the kidney of obese rats. Nutr. Diabetes 2014, 4, e142. [Google Scholar] [CrossRef]
- Fenwick, M.A.; Hurst, P.R. Immunohistochemical localization of active caspase-3 in the mouse ovary: Growth and atresia of small follicles. Reproduction 2002, 124, 659–665. [Google Scholar] [CrossRef]
- Harris, B.S.; Jukic, A.M.; Truong, T.; Nagle, C.T.; Erkanli, A.; Steiner, A.Z. Markers of ovarian reserve as predictors of future fertility. Fertil. Steril. 2023, 119, 99–106. [Google Scholar] [CrossRef]
- Zhang, J.; Fang, L.; Shi, L.; Lai, Z.; Lu, Z.; Xiong, J.; Wu, M.; Luo, A.; Wang, S. Protective effects and mechanisms investigation of Kuntai capsule on the ovarian function of a novel model with accelerated aging ovaries. J. Ethnopharmacol. 2017, 195, 173–181. [Google Scholar] [CrossRef]
- Liu, S.; Jia, Y.; Meng, S.; Luo, Y.; Yang, Q.; Pan, Z. Mechanisms of and Potential Medications for Oxidative Stress in Ovarian Granulosa Cells: A Review. Int. J. Mol. Sci. 2023, 24, 9205. [Google Scholar] [CrossRef]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M. Lycopene as a natural antioxidant used to prevent human health disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Week 1 | Week 2 | Week 3 | Week 4 | ||||
|---|---|---|---|---|---|---|---|---|
| R1 | saline | saline | saline | saline | saline | saline | PMSG | hCG |
| R4 | PMSG | hCG | PMSG | hCG | PMSG | hCG | PMSG | hCG |
| R4-Lyc | PMSG; lycopene | hCG; lycopene | PMSG; lycopene | hCG; lycopene | PMSG; lycopene | hCG; lycopene | PMSG; lycopene | hCG; lycopene |
| Researched Marker | Primer | Product Size (bp) | Annealing Temp. (°C) | GenBank Isolate |
|---|---|---|---|---|
| Sod3 | F5′-TTCTACGGCTTGCTACTGGC-3′ R5′-GCTAGGTCGAAGCTGGACTC-3′ | 74 | 60 | NM_011435.3 |
| Cat | F5′-CACTGACGAGATGGCACACT-3′ R5′-TGTGGAGAATCGAACGGCAA-3′ | 175 | 60 | NM_009804.2 |
| Nrf2 | F5′-CCTCACCTCTGCTGCAAGTA-3′ R5′-TCAAATCCATGTCCTGCTGGG-3′ | 120 | 58 | NM_010902.5 |
| Keap1 | F5′-TCGTAGGGTGGTGGCCG-3′ R5′-ATGGGGTTCCGGATGACAAG-3′ | 78 | 60 | AB020063.1 |
| Tnf | F5′-GCCTCTTCTCATTCCTGCTTGT-3′ R5′-CACTTGGTGGTTTGCTACGACG-3′ | 203 | 58 | AY423855.1 |
| Nfkb | F5′-CTCTGGCACAGAAGTTGGGT-3′ R5′-CCCGGAGTTCATCTCATAGTTGT-3′ | 101 | 58 | NM_001410442.1 |
| Casp3 | F5′-GGGGAGCTTGGAACGCTAAG-3′ R5′-CCGTACCAGAGCGAGATGAC-3′ | 232 | 58 | NM_009810.3 |
| β-actin | F5′-AGGGAAATCGTGCGTGACAT-3′ R5′-TCCAGGGAGGAAGAGGATGC-3′ | 59 | 60 | AY618569.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakha, S.I.; Ateya, A.I.; Safhi, F.A.; Abdellatif, A.M. Ameliorative Effect of Lycopene on Follicular Reserve Depletion, Oxidative Damage, Apoptosis Rate, and Hormonal Profile during Repeated Superovulations in Mice. Vet. Sci. 2024, 11, 414. https://doi.org/10.3390/vetsci11090414
Rakha SI, Ateya AI, Safhi FA, Abdellatif AM. Ameliorative Effect of Lycopene on Follicular Reserve Depletion, Oxidative Damage, Apoptosis Rate, and Hormonal Profile during Repeated Superovulations in Mice. Veterinary Sciences. 2024; 11(9):414. https://doi.org/10.3390/vetsci11090414
Chicago/Turabian StyleRakha, Shimaa I., Ahmed I. Ateya, Fatmah A. Safhi, and Ahmed M. Abdellatif. 2024. "Ameliorative Effect of Lycopene on Follicular Reserve Depletion, Oxidative Damage, Apoptosis Rate, and Hormonal Profile during Repeated Superovulations in Mice" Veterinary Sciences 11, no. 9: 414. https://doi.org/10.3390/vetsci11090414