
Pulsatilla Powder Ameliorates Damp-Heat Diarrhea in Piglets Through the Regulation of Intestinal Mucosal Barrier and the Pentose Phosphate Pathway Involving G6PD and NOX


Simple Summary
Abstract

1. Introduction
2. Materials and Method
2.1. Animals
2.2. Preparation of PP
2.3. Primary Instruments and Reagents
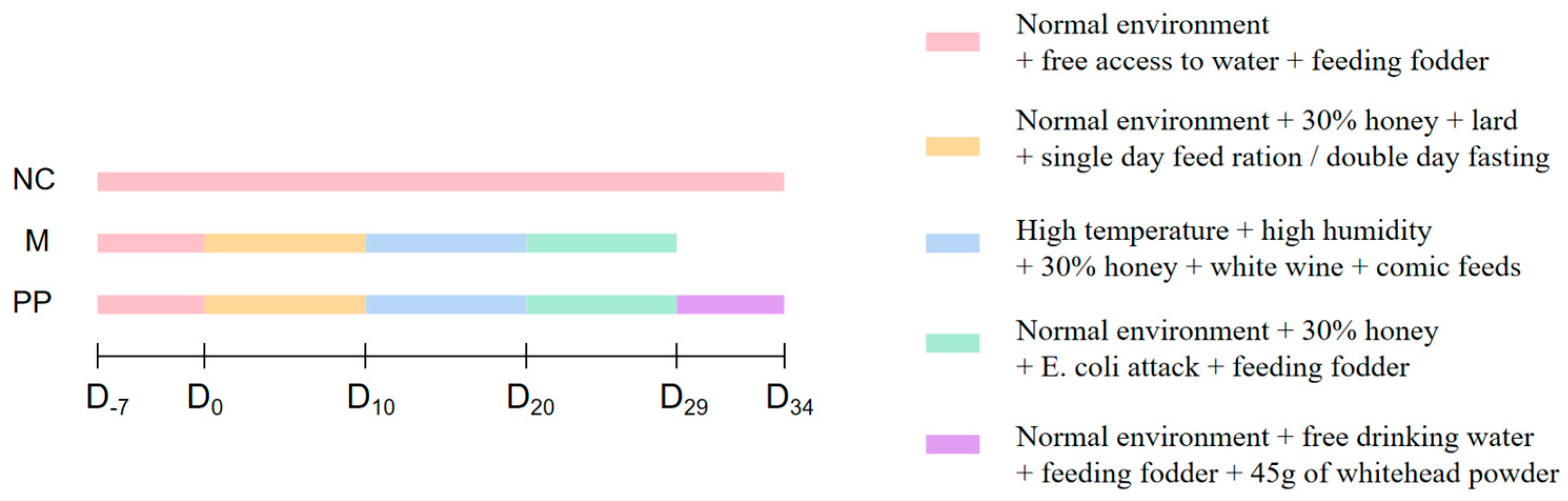
2.4. Animal Model Preparetion and PP Treatment
2.5. Clinical Evaluation of DHD
2.6. Histopathological Examination
2.7. Colon Metabolomics Profiling
2.7.1. Preparation of Colon Tissue Samples
2.7.2. UHPLC-MS/MS Analysis
2.7.3. Data Processing and Analysis in Metabolomics
2.8. Quantitative Real-Time PCR
Determination of Colon G6PP and ROS
2.9. Western Blot Analysis
2.10. Immunohistochemistry
2.11. Statistical Analysis
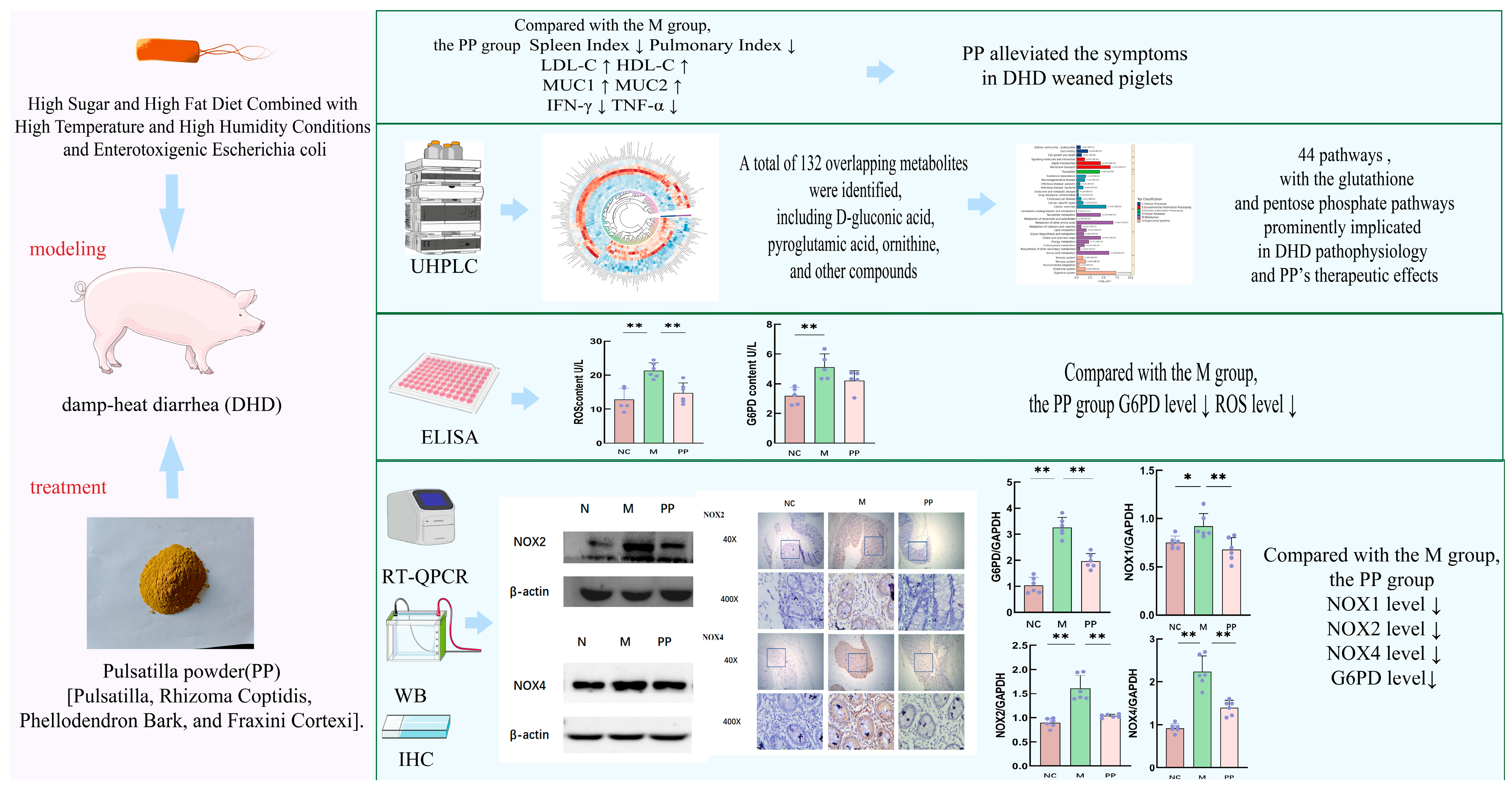
3. Results
3.1. Identification of the Main Components in PP and PP Alleviated the Symptoms in DHD Piglets
3.2. PP Attenuated Colon Pathological Injury in DHD Piglets
3.3. Colon Metabolomics Results
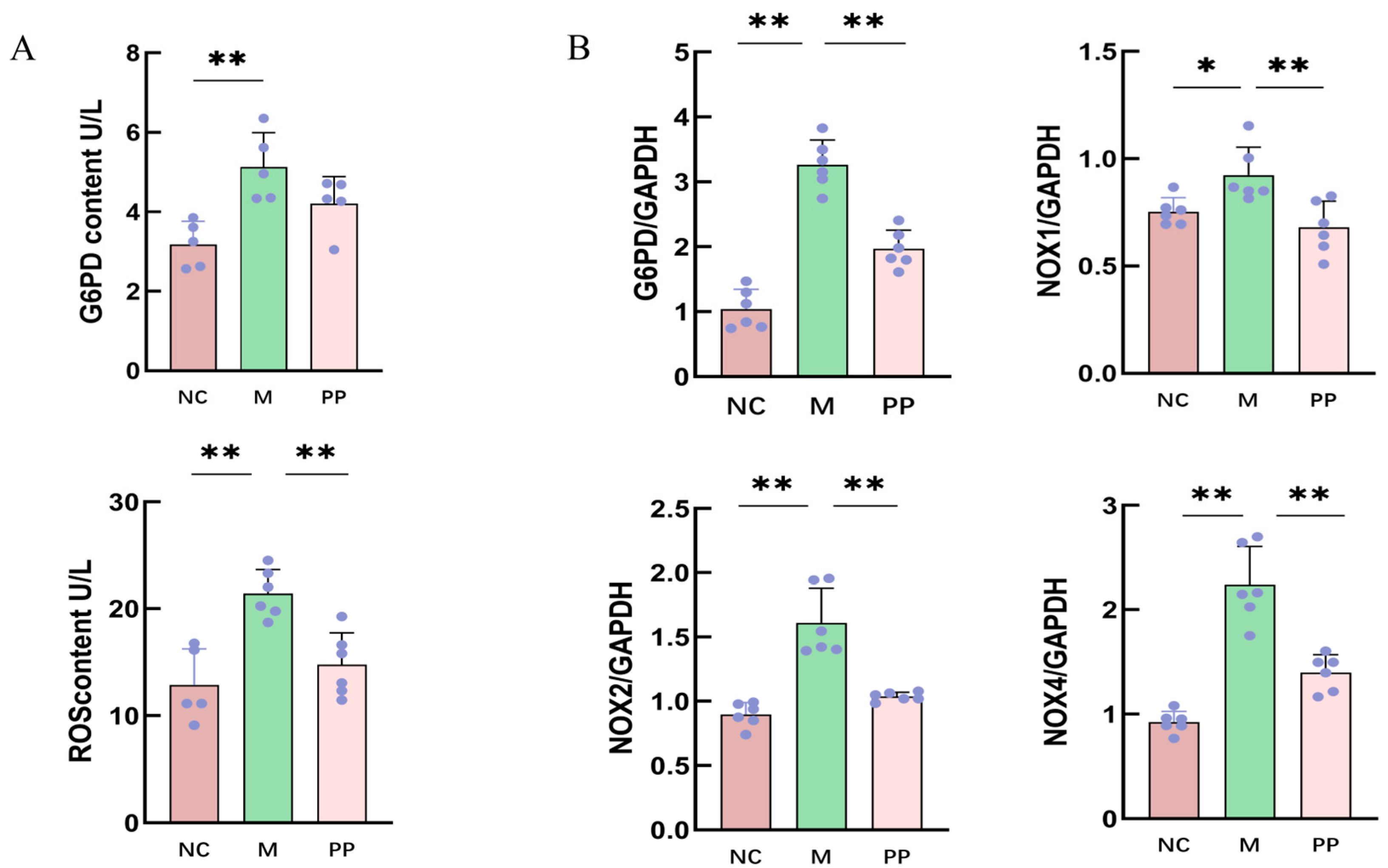
3.4. Inhibitory Effect of PP on G6PD/ROS Signaling Pathway
3.4.1. Gene Expression Analysis of Pentose Phosphate Pathway
3.4.2. Protein Expression of NOX2 and NOX4
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, Q.; Lan, T.; Zhou, C.; Gao, J.; Wu, L.; Wei, H.; Li, W.; Tang, Z.; Tang, W.; Diao, H.; et al. Nutrition strategies to control post-weaning diarrhea of piglets: From the perspective of feeds. Anim. Nutr. 2024, 17, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Hou, L.; Sun, L.; Gao, J.; Gao, K.; Yang, X.; Jiang, Z.; Wang, L. Intestinal morphology and immune profiles are altered in piglets by early-weaning. Int. Immunopharmacol. 2022, 105, 108520. [Google Scholar] [CrossRef]
- Jiang, S.; Ma, Q.; Liao, H.; Cao, L.; Du, H.; Tan, S.; Zhou, J. Establishment and comparison of damp-heat diarrhea related animal models. Chin. J. Vet. Med. 2019, 43, 2597–2605. (In Chinese) [Google Scholar]
- Gao, W.X. Study on Growth Performance, Physiological and Biochemical Indexes of Weaned Piglets and the Effect of Prevention and Treatment of Escherichia coli Diarrhea. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2023. [Google Scholar]
- Qiao, L.; Dou, X.; Song, X.; Chang, J.; Yi, H.; Xu, C. Targeting mitochondria with antioxidant nutrients for the prevention and treatment of postweaning diarrhea in piglets. Anim. Nutr. 2023, 15, 275–287. [Google Scholar] [CrossRef]
- Xu, X.; Wei, Y.; Hua, H.; Jing, X.; Zhu, H.; Xiao, K.; Zhao, J.; Liu, Y. Polyphenols sourced from Ilex latifolia Thunb. relieve intestinal injury via modulating ferroptosis in weanling piglets under oxidative stress. Antioxidants 2022, 11, 966. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Study on the Mechanism of Action of Paektuonung Decoction in the Treatment of Damp-Heat Diarrhea Based on Metabolomics and Network Pharmacology. Ph.D. Thesis, Gansu Agricultural University, Lanzhou, China, 2018. [Google Scholar]
- Otaka, M.; Odashima, M.; Watanabe, S. Role of heat shock proteins (molecular chaperones) in intestinal mucosal protection. Biochem. Biophys. Res. Commun. 2006, 348, 1–5. [Google Scholar] [CrossRef]
- Pizarro, J.G.; Folch, J.; Vazquez De la Torre, A.; Verdaguer, E.; Junyent, F.; Jordán, J.; Pallàs, M.; Camins, A. Oxidative stress-induced DNA damage and cell cycle regulation in B65 dopaminergic cell line. Free Radic. Res. 2009, 43, 985–994. [Google Scholar] [CrossRef]
- Holmes, B.; Page, A.R.; Good, R.A. Studies of the metabolic activity of leukocytes From patients with a genetic abnormality of phagocytic function. J. Clin. Investig. 1967, 46, 1422–1432. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, L.; Long, Y.; Cao, J.; Qu, Q.; Wu, Q.; Liu, L.; Chang, X.; Ai, K.; Qi, F. Effects of moxibustion on CD4+ T-cell proliferation by G6PD-mediated pentose phosphate pathway in adjuvant arthritis rats. J. Acupunct. Tuina Sci. 2024, 22, 273–279. [Google Scholar] [CrossRef]
- Pereira, A.R.; Dias, J.C.E.; de Souza Oliveira, L.; Bitencourt, F.; Mazzola, P.N.; Coelho, J.G.; Mescka, C.P.; Dutra-Filho, C.S. Neonatal hyperglycemia induces oxidative stress in the rat brain: The role of pentose phosphate pathway enzymes and NADPH oxidase. Mol. Cell. Biochem. 2015, 403, 159–167. [Google Scholar]
- Jiao, Y.; Kang, Y.; Wen, M.; Wu, H.; Zhang, X.; Huang, W. Fast Antioxidation Kinetics of Glutathione Intracellularly Monitored by a Dual-Wire Nanosensor. Angew. Chem. 2023, 62, e202313612. [Google Scholar] [CrossRef]
- Rathi, D.; Rossi, C.; Pospíšil, P.; Manoharan, R.R.; Talarico, L.; Magnani, A.; Prasad, A. NOX2 and NOX4 expression in monocytes and macrophages-extracellular vesicles in signalling and therapeutics. Front. Cell Dev. Biol. 2024, 12, 1342227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, S.; Wang, L.; Chen, S.; Xu, X. Inhibitory effect of compound Paetuonung powder on isolates of Salmonella typhimurium derived from geese in vitro. Chin. J. Vet. Med. 2022, 58, 62–67. [Google Scholar]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef]
- Wu, F.; Lin, L.; Zeng, Y.; Wu, X. Clinical application of Baitouweng Decoction in the treatment of anorectal inflammatory diseases. Chin. Mod. Distance Educ. Tradit. Chin. Med. 2019, 21, 149–151. (In Chinese) [Google Scholar]
- Wen, Y. Effects of Yujin Powder on Gastrointestinal Hormone and HPA Axis in Large Intestine Damp-Heat Syndrome Model Rats. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2018. [Google Scholar]
- Wei, J.; Mo, J.; Cao, J. The influence of different weeks weaning to sow distribution effect. J. Livest. Expo. 2007, 12, 14–15. [Google Scholar]
- GB 14963-2011; National Food Safety Standard for Honey. China Standards Press: Beijing, China, 2011.
- GB/T 10781.2-2022; National Technical Committee for Baijiu Standardization (SAC/TC 358) Quality requirements for Baijiu—Part 2: Clear-Aroma Baijiu. China Standards Press: Beijing, China, 2022.
- Da Silva, V.C.; De Araújo, A.A.; Araújo, D.F.D.S.; Lima, M.C.J.S.; Vasconcelos, R.C.; de Araújo Júnior, R.F.; Langasnner, S.M.Z.; Pedrosa, M.D.F.F.; De Medeiros, C.A.C.X.; Guerra, G.C.B. Intestinal Anti-Inflammatory Activity of the Aqueous Extract from Ipomoea asarifolia in DNBS-Induced Colitis in Rats. Int. J. Mol. Sci. 2018, 19, 4016. [Google Scholar] [CrossRef]
- Xue, Y. Viscera Index, Intestinal Morphology and pH Characteristics of Gastrointestinal Contents of Piglets with Diarrhea and Effects of Enterococcus faecalis Administration on Them. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2017. [Google Scholar]
- Liu, X. Zinc Oxide on the Influence and Mechanism of Weaned Piglets Diarrhea Caused by ETEC Research. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2023. [Google Scholar]
- Wu, D.; Wu, K.; Cheng, W.; Li, W.; Wang, T.; Liang, Y.; Wang, Z. Effect of Guchang Zhixie pills on mice with ulcerative colitis. J. Hunan Univ. Chin. Med. 2022, 42, 1626–1631. (In Chinese) [Google Scholar]
- Wang, B.; Jiang, L.; Ji, P.; Yao, W.; Zhang, W.; Zhao, W.; Qin, X.; Wei, Y.; Cai, Y.; Yuan, Z. Exploration of the Potential Mechanism of Yujin Powder Treating Dampness-heat Diarrhea by Integrating UPLC-MS/MS and Network Pharmacology Prediction. Comb. Chem. High Throughput Screen. 2024, 27, 1466–1479. [Google Scholar]
- Vancamelbeke, M.; Vanuytsel, T.; Farré, R.; Verstockt, S.; Ferrante, M.; Van Assche, G.; Rutgeerts, P.; Schuit, F.; Vermeire, S.; Arijs, I.; et al. Genetic and transcriptomicbases of intestinal epithelial barrier dysfunction in inflammatory bowel disease. Inflamm. Bowel Dis. 2017, 23, 1718. [Google Scholar] [CrossRef]
- Zhang, Y. Study on the Mechanism of Mucin MUC1 Inducing Immunosuppression in Colon Cancer. Ph.D. Thesis, Kunming Medical University, Kunming, China, 2021. [Google Scholar]
- Wang, R.; Zhang, T.; Chai, Y. Inflammatory bowel disease disturbance and its related metabolic pathways of drug development. J. Med. 2017, 41, 8. [Google Scholar]
- Leon-Coria, A.; Kumar, M.; Workentine, M.; Moreau, F.; Surette, M.; Chadee, K. Muc2 mucin and nonmucin microbiota confer distinct innate host defense in disease susceptibility and colonic injury. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 77–98. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Zhang, X.; Wang, M.; Wang, J.; Hu, J.; Zhang, L.; Fu, Z.; Shi, L. Effect of Chinese drug-containing serum on the regulation of NLRP6 pathway on MUC2 expression in Caco-2/HT29-MTX cell co-culture model. J. Chin. Med. 2013, 52, 13–18. [Google Scholar]
- Chen, L.; Zhang, Z.; Hoshino, A.; Zheng, H.D.; Morley, M.; Arany, Z.; Rabinowitz, J.D. NADPH production by the oxidative pentose-phosphate pathway supports folate metabolism. Nat. Metab. 2019, 1, 404–415. [Google Scholar] [CrossRef]
- Wang, W.; Sun, Z.; Deng, H. Study on the theoretical basis of Wujia Xiao Zhengqi Powder in the treatment of rotavirus enteritis from the perspective of “Spleen-making”. Shi Zhen Chin. Med. 2024, 35, 2410–2413. [Google Scholar]
- Jia, R. Clinical Study of Qinghuoliang Decoction in the Treatment of Diarrheal Irritable Bowel Syndrome with Spleen-Deficiency Damp-Heat Syndrome. Master’s Thesis, Nanjing University of Chinese Medicine, Nanjing, China, 2024. [Google Scholar]
- Mohammadshahi, J.; Ghobadi, H.; Matinfar, G.; Boskabady, M.H.; Aslani, M.R. Role of Lipid Profile and Its Relative Ratios (Cholesterol/HDL-C, Triglyceride/HDL-C, LDL-C/HDL-C, WBC/HDL-C, and FBG/HDL-C) on Admission Predicts In-Hospital Mortality COVID-19. J. Lipids 2023, 2023, 6329873. [Google Scholar] [CrossRef]
- Li, X.H. Correlation Between HDL, APOA1 and Activity of Inflammatory Bowel Disease and Their Effect on Experimental Mouse Enteritis. Master’s Thesis, Yangzhou University, Yangzhou, China, 2020. [Google Scholar]
- Yan, Z.X. Study on the Mechanism of Huangbai Jianpi Oral Liquid in the Treatment of Damp-Heat Diarrhea of Calves Based on Omics Technology. Ph.D. Thesis, Northwest Agriculture and Forestry University of Science and Technology, Xianyang, China, 2022. [Google Scholar]
- Pan, Z.; Shao, S.; Huang, D.; Zheng, A. Effects of Hudi enteric-soluble capsule combined with oxalazine on intestinal mucosal lesions, serum inflammatory factors, NGAL and MMP-9 expression in patients with chronic ulcerative colitis. Zhejiang Med, J. 2021, 21, 2337–2340. [Google Scholar]
- Yang, M. Expression and Significance of NADPH Oxidase Nox1 and Duox2 in Mice Enteritis. Master’s Thesis, Henan University of Science and Technology, Luoyang, China, 2016. [Google Scholar]
- Han, X.; Gao, L.; Liu, Q. Research progress of NOX family proteins. Life Sci. 2012, 24, 568–577. (In Chinese) [Google Scholar]
- Abid, M.R.; Kachra, Z.; Spokes, K.C.; Aird, W.C. NADPH oxidaseactivity is required for endothelial cell proliferation andmigration. FEBS Lett. 2000, 486, 252–256. [Google Scholar] [CrossRef]
- Hayes, P.; Dhillon, S.; O’Neill, K.; Thoeni, C.; Hui, K.Y.; Elkadri, A.; Guo, C.H.; Kovacic, L.; Aviello, G.; Alvarez, L.A.; et al. Defects in NADPH Oxidase Genes NOX1and DUOX2 in Very Early Onset Inflammatory Bowel Disease. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 489–502. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancercells by ROS-mediated mechanisms:a radical therapeuticapproach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; JeanClaude, M.; AnneLaure, P.; Zhuoyao, S.; Marwa, B.; Kaouthar, B.; Coralie, P.; Thibaut, L.; Samuel, T.; Guillaume, C.; et al. Protein Kinase CK2 acts as a molecular brake to control NADPH oxidase 1 activation and colon inflammation. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1073–1093. [Google Scholar] [CrossRef]
- Cai, P.; Zhang, D.; Yang, J.; Wang, L.; Hu, L. Effect of cisplatin on the expression and activity of glucose 6-phosphate dehydrogenase in HeLa cells. Chin. J. Pathol. Physiol. 2008, 24, 1323–1326. [Google Scholar]
- Chen, J.; Yu, G.; Xie, B. Effects of Qingzao Jiufei Decoction on the activity of G6PD, a key enzyme of pentose phosphate energy metabolism pathway in lung cancer and its regulatory factors. Chin. J. Exp. Formulae 2020, 26, 59–63. [Google Scholar]
- Xu, C.; Ye, Z.; Jiang, W.; Wang, S.; Zhang, H. Cyclosporine A alleviates colitis by inhibiting the formation of neutrophil extracellular traps via the regulating pentose phosphate pathway. Mol. Med. 2023, 29, 169. [Google Scholar] [CrossRef]
- Xuan, Q.; Dai, G.; Chen, P.; Feng, D.; Jin, J.; Gao, Q. Expression of NADPH oxidase NOX1, NOX2 and DUOX2 in ulcerative colitis. J. Cap. Med. Univ. 2017, 38, 220–226. [Google Scholar]
- Brandes, R.P.; Schröder, K. Composition and functions of vascular nicotinamide adenine dinucleotide phosphate oxidases. Trends Cardiovasc. Med. 2008, 18, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Ma, T. The correlation between NOX1 and NOX2 expression levels in ulcerative colitis and intestinal mucosal oxidative stress response and barrier function impairment. J. Hainan Med. Coll. 2017, 23, 2638–2641. [Google Scholar]
- Singel, K.L.; Segal, B.H. NOX2-dependent regulation of inflammation. Clin. Sci. 2016, 130, 479–490. [Google Scholar] [CrossRef]
- Cifuentes-Pagano, E.; Csanyi, G.; Pagano, P.J. NADPH oxidase inhibitors: A decade of discovery from Nox2ds to HTS. Cell Mol. Life Sci. 2012, 69, 2315–2325. [Google Scholar] [CrossRef]
- Allaoui, A.; Botteaux, A.; Dumont, J.E.; Hoste, C.; De Deken, X. Dual oxidases and hydrogen peroxide in a complex dialogue between host mucosae and bacteria. Trends Mol. Med. 2009, 15, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Giang, L.; Vasso, A.; Anthony, Z.; Kulmira, N. NADPH oxidases and inflammatory bowel disease. Curr. Med. Chem. 2015, 22, 2100–2109. [Google Scholar]
- Bauer, K.M.; Watts, T.N.; Buechler, S.; Hummon, A.B. Proteomic and functional investigation of the colon cancer relapse-associated genes NOX4 and ITGA3. J. Proteome Res. 2014, 13, 4910–4918. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, S.; Ma, Q.; Du, H.; Bi, S.; Cao, L. The effect of Baitouweng Decoction on the ileal mucosal barrier function of rats with damp-heat diarrhea. J. Southwest Univ. (Nat. Sci. Ed.) 2024, 46, 85–96. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences (5′→3′) |
|---|---|
| GAPDH-F | TCGGAGTGAACGGATTTGGC |
| GAPDH-R | TGACAAGCTTCCCGTTCTCC |
| MUC1-F | GTGCTTACAGGTGAGGGGC |
| MUC1-R | ACAGATCCTGGCCTGAACTT |
| MUC2-F | GAACGGGGCCATGGTCAG |
| MUC2-R | AGCATGACCGAGTCCTCTCT |
| TNF-α-F | GGCCCAAGGACTCAGATCAT |
| TNF-α-R | CTGTCCCTCGGCTTTGACAT |
| IFN-γ-F | TGAAGAATTGGAAAGAGGAGAGTGA |
| IFN-γ-R | GCTCCTTTGAATGGCCTGGT |
| NOX1-F | AATGGCATCCCTTTACCCTGACCT |
| NOX1-R | CTTGGAACTGGCGAATGCTGTTGT |
| NOX2-F | CCATATCCGCATTGTTGGCG |
| NOX2-R | CCGTCCACAGCGATCTTAGG |
| NOX4-F | GGAACGCACTACCAGGATGT |
| NOX4-R | CAGGTCTGCGGAAAGTTAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, Y.; Ma, Q.; Wang, C.; Zhang, L.; Feng, H.; Lai, S. Pulsatilla Powder Ameliorates Damp-Heat Diarrhea in Piglets Through the Regulation of Intestinal Mucosal Barrier and the Pentose Phosphate Pathway Involving G6PD and NOX. Vet. Sci. 2025, 12, 403. https://doi.org/10.3390/vetsci12050403
Qu Y, Ma Q, Wang C, Zhang L, Feng H, Lai S. Pulsatilla Powder Ameliorates Damp-Heat Diarrhea in Piglets Through the Regulation of Intestinal Mucosal Barrier and the Pentose Phosphate Pathway Involving G6PD and NOX. Veterinary Sciences. 2025; 12(5):403. https://doi.org/10.3390/vetsci12050403
Chicago/Turabian StyleQu, Yunqi, Qi Ma, Chenying Wang, Lifang Zhang, Haolian Feng, and Siyue Lai. 2025. "Pulsatilla Powder Ameliorates Damp-Heat Diarrhea in Piglets Through the Regulation of Intestinal Mucosal Barrier and the Pentose Phosphate Pathway Involving G6PD and NOX" Veterinary Sciences 12, no. 5: 403. https://doi.org/10.3390/vetsci12050403
APA StyleQu, Y., Ma, Q., Wang, C., Zhang, L., Feng, H., & Lai, S. (2025). Pulsatilla Powder Ameliorates Damp-Heat Diarrhea in Piglets Through the Regulation of Intestinal Mucosal Barrier and the Pentose Phosphate Pathway Involving G6PD and NOX. Veterinary Sciences, 12(5), 403. https://doi.org/10.3390/vetsci12050403