
Comparative Study of the Gastric Mucosa of Risso’s Dolphin (Grampus griseus) and Bottlenose Dolphin (Tursiops truncatus): A Key to Manage the Diet in Captive Conditions
 , ,
, ,  , , ,
, , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Morphological and Immunohistochemical Analysis
2.3. Statistical Analysis
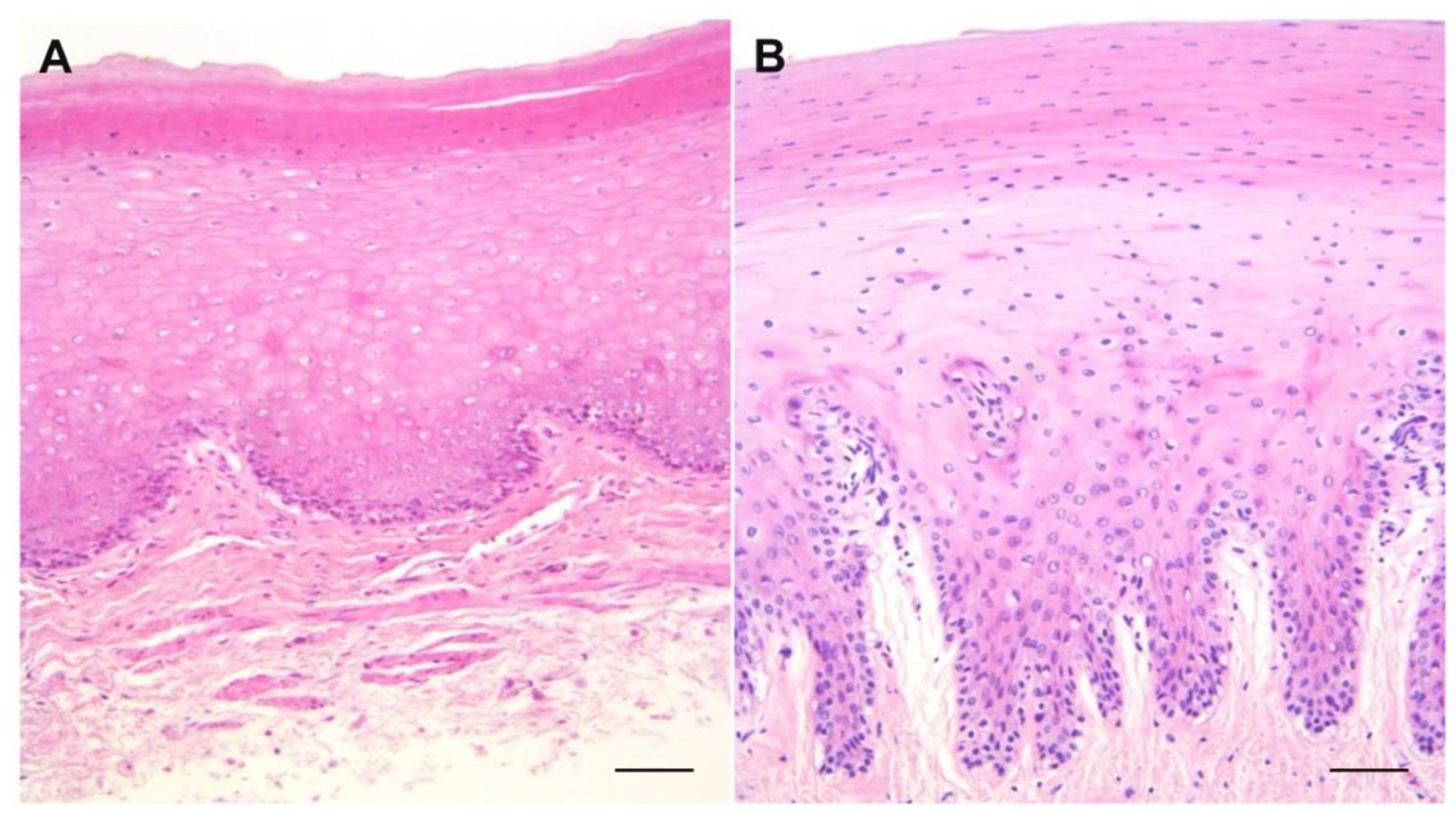
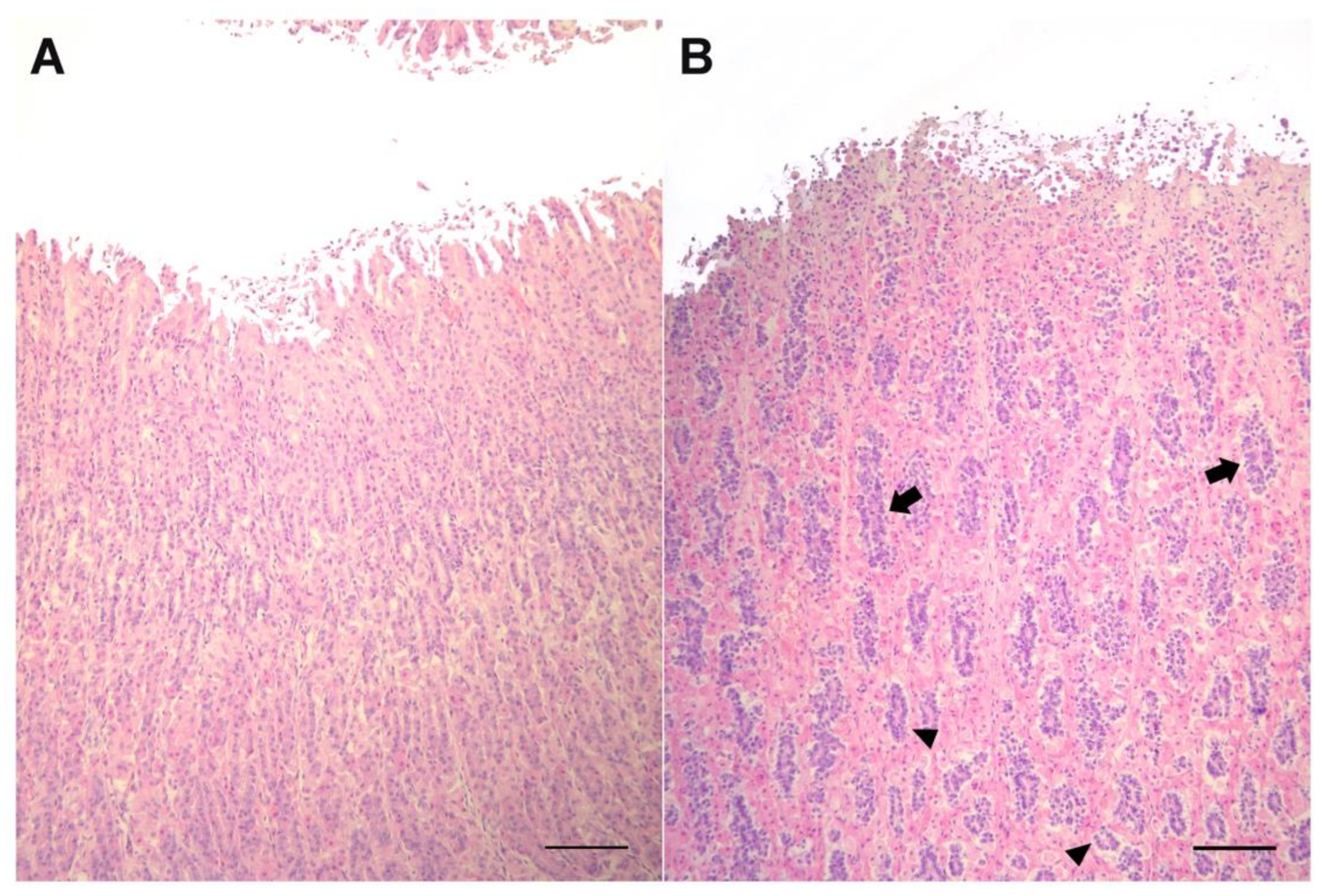
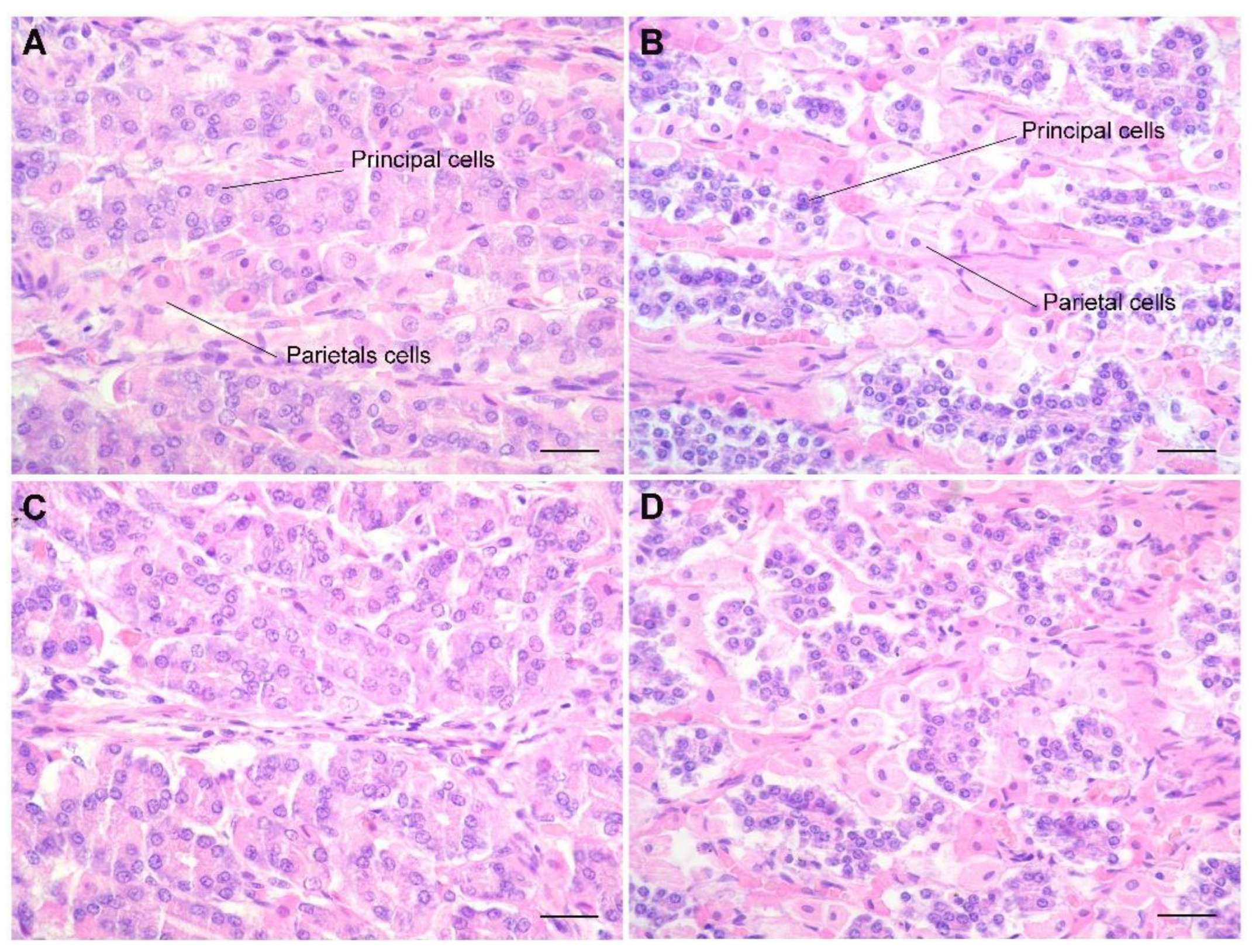
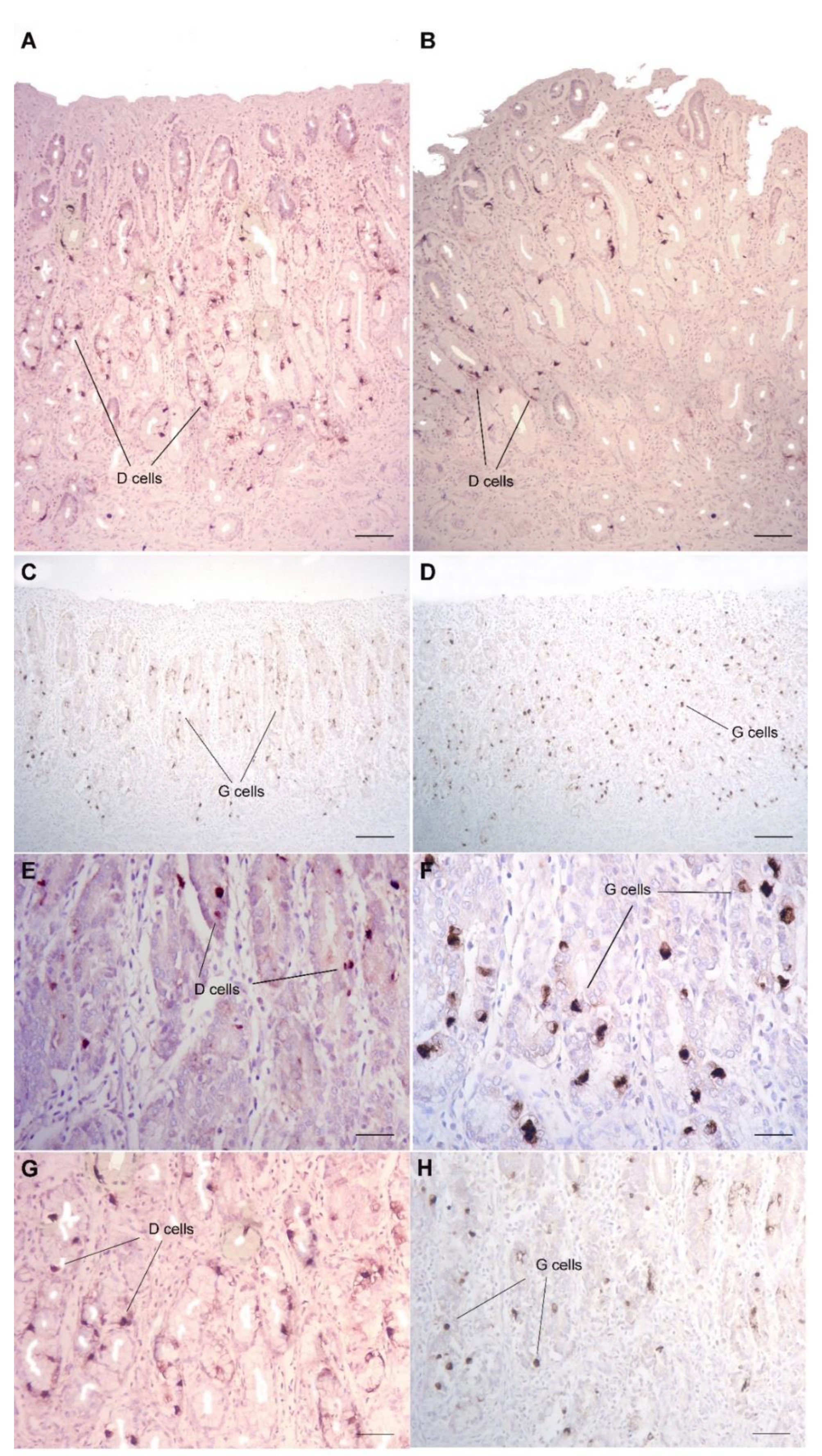
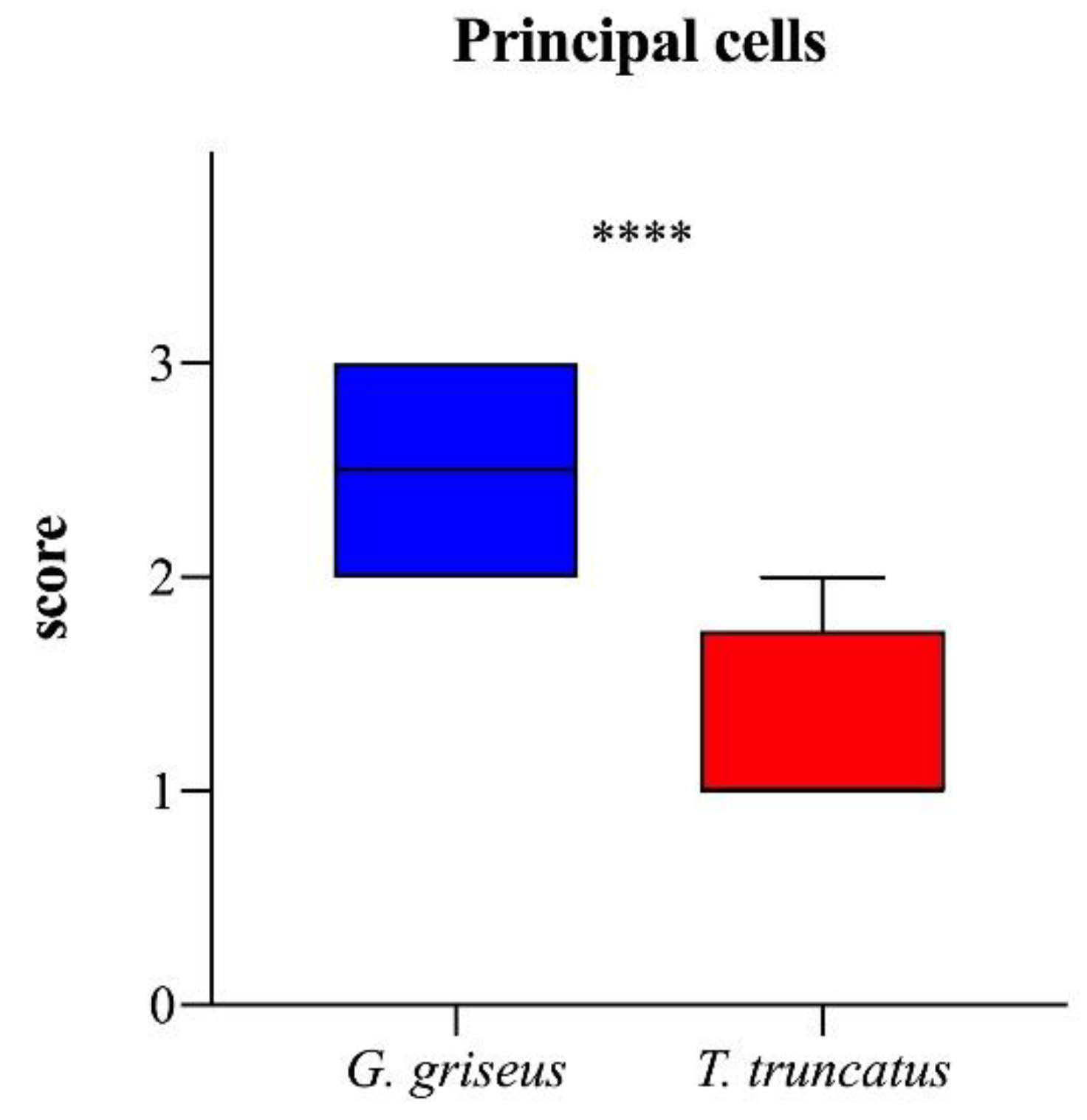
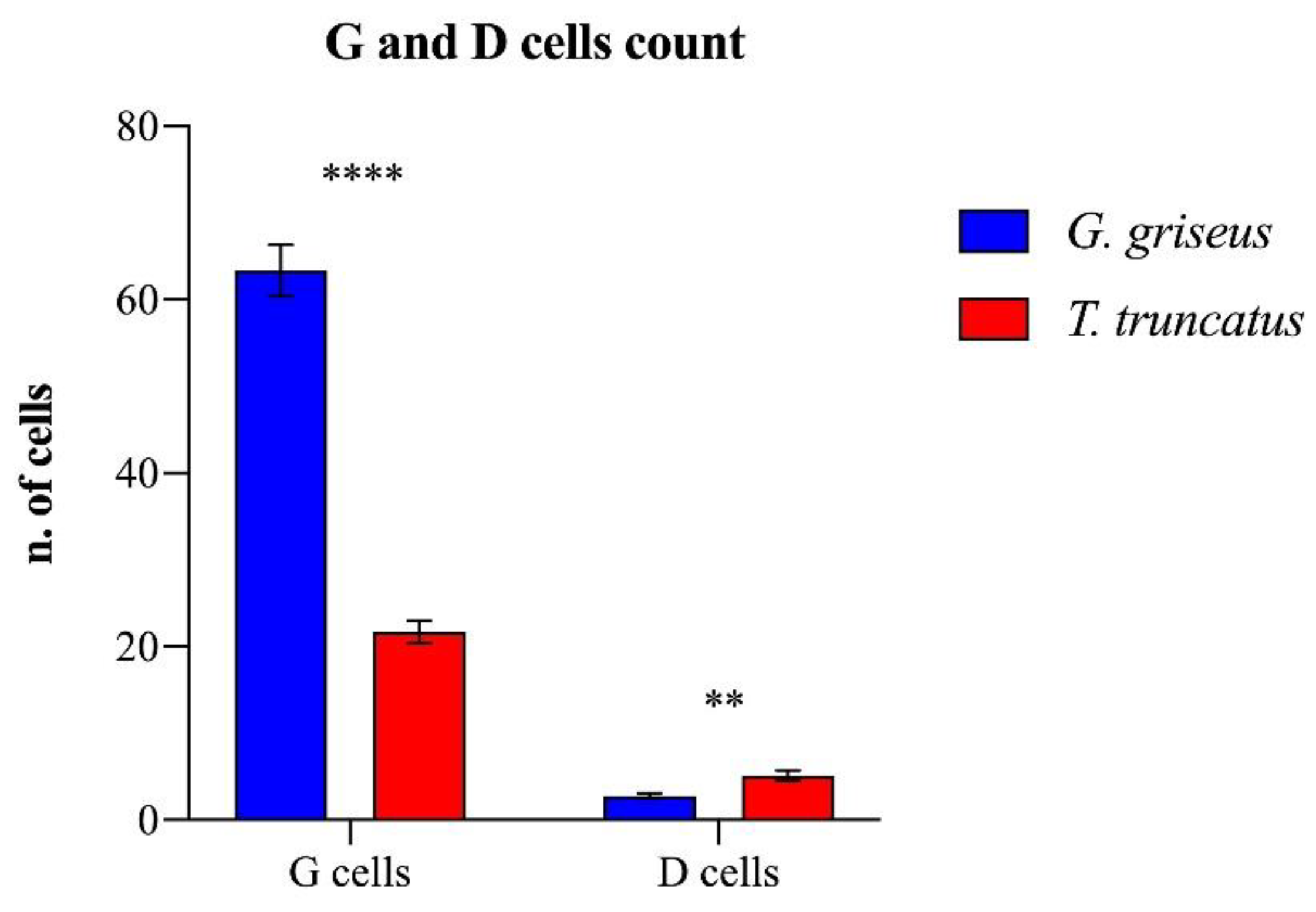
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murie, J. On Risso’s grampus (Grampus rissoanus, Desm.). J. Anat. Physiol. 1870, 5, 118–138. [Google Scholar] [PubMed]
- Turner, W. A contribution to the anatomy of the Pilot whale (Globicephalus svineval, Lacépede). J. Anat. Physiol. 1867, 2, 66–79. [Google Scholar]
- Turner, W. Further observations on the stomach in the Cetacea. J. Anat. Physiol. 1868, 3, 117–119. [Google Scholar]
- Turner, W. Notes on some of the viscera of Risso’s dolphin (Grampus griseus). J. Anat. Physiol. 1889, 23, 466–492. [Google Scholar] [PubMed]
- Turner, W. Additional observations of the stomach in the ziphioid and delphinoid whales. J. Anat. Physiol. 1892, 26, 258–270. [Google Scholar] [PubMed]
- Harrison, R.J.; Johnson, F.R.; Young, B.A. The oesophagus and stomach of dolphins (Tursiops, Delphinus, Stenella). J. Zool. 1970, 160, 377–390. [Google Scholar] [CrossRef]
- Rommel, S.A.; Lowenstein, L.J. Gross and microscopic anatomy. In CRC Handobook of Marine Mammal Medicine, 2nd ed.; Dierauf, L.A., Gulland, F.M.D., Eds.; CRC: Boca Raton, FL, USA, 2001; pp. 12–163. [Google Scholar]
- Dockray, G.J. Gastrin and gastric epithelial physiology. J. Physiol. 1999, 518, 315–324. [Google Scholar] [CrossRef]
- Hunt, R.H.; Camilleri, M.; Crowe, S.E.; El-Omar, E.M.; Fox, J.G.; Kuipers, E.J.; Malfertheiner, P.; McColl, K.E.L.; Pritchard, D.M.; Rugge, M.; et al. The stomach in health and disease. Gut 2015, 64, 1650–1668. [Google Scholar] [CrossRef] [PubMed]
- Soybel, D.I. Anatomy and physiology of the stomach. Surg. Clin. N. Am. 2005, 85, 875–894. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, B.; Huggenberger, S.; Oelschlager, H. Anatomy of Dolphins Insights into Body Structure and Function; Elsevier Publications: San Diego, CA, USA, 2016. [Google Scholar]
- Smith, G.J.D. The stomach of the harbor porpoise Phocoena phocoena (L.). Can. J. Zool. 1972, 50, 1611–1616. [Google Scholar] [CrossRef]
- Gaskin, D.E. Form and function in the digestive tract and associated organs in cetacea, with a consideration of metabolic rates and specific energy budgets. Oceanogr. Mar. Biol. Ann. Rev. 1978, 16, 313–345. [Google Scholar]
- Rommel, S.A.; Costidis, A.M.; Lowenstein, L.J. Gross and microscopic anatomy. In CRC Handobook of Marine Mammal Medicine, 3rd ed.; Dierauf, L.A., Gulland, F.M.D., Whitman, L., Eds.; CRC: Boca Raton, FL, USA, 2018; pp. 89–136. [Google Scholar]
- Geraci, J.R.; Gerstmann, K.E. Relationship of dietary histamine to gastric ulcers in the dolphin. J. Am. Vet. Med. Assoc. 1966, 149, 884–890. [Google Scholar] [PubMed]
- Norday, E.S. Do mink whales (Balenoptera acutorostrata) digest wax esters? Br. J. Nutr. 1995, 74, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.A.; Aagnes, T.H.; Marthiesen, S.D. Digestion of herring by indigenous bacteria in the minke whales forestomach. Appl. Environ. Microbiol. 1994, 60, 4445–4455. [Google Scholar] [CrossRef] [PubMed]
- Biancani, B.; Saviano, P.; Rossi, G. Gastric compartments: Comparative study between bottlenose dolphin (Tursiops truncatus) and Risso’s dolphin (Grampus griseus). In Proceedings of the 41th International Association Aquatic Animal Medicine Annual Meeting, Vancouver, BC, Canada, 8–12 May 2010. [Google Scholar]
- Bearzi, G.; Reeves, R.; Remonato, E.; Pierantonio, N.; Airoldi, S. Risso’s dolphin (Grampus griseus) in the Mediterranean Sea. Mamm. Biol. 2011, 76, 385–400. [Google Scholar] [CrossRef]
- Kruse, S.; Caldwell, D.K.; Caldwell, M.C. Risso’s dolphin Grampus griseus (G. Cuvier, 1812). In Handbook of Marine Mammals, the Second Book of Dolphins and Porpoises; Academic Press: San Diego, CA, USA, 1999; Volume 6, pp. 183–212. [Google Scholar]
- Öztürk, B.; Salman, A.; Öztürk, A.; Tonay, A. Cephalopod remains in the diet of Striped Dolphins (Stenella coeruleoalba) and Risso’s Dolphins (Grampus griseus) in the Eastern Mediterranean Sea. Vie Milieu 2007, 57, 53–59. [Google Scholar]
- Blanco, C.; Salomon, O.; Raga, J.A. Diet of the bottlenose dolphin (Tursiops truncatus) in the western Mediterranean Sea. J. Mar. Biol. Assoc. UK 2001, 81, 1053–1058. [Google Scholar] [CrossRef]
- Gimenez, J.; Marcalo, A.; Ramirez, F.; Verborgh, P.; Gauffier, P.; Esteban, R.; Nicolau, L.; Gonzalez-Ortegon, E.; Balbo, F.; Vilas, C.; et al. Diet of bottlenose dolphins (Tursiops truncates) from the gulf of Cadiz: Insights from stomach content and stable isotope analyses. PLoS ONE 2017, 12, e0184673. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Description | |
|---|---|---|
| Presence of principal cells | 0 | None, 0–15 cells per HPF |
| 1 | Scarce, 16–50 cells per HPF | |
| 2 | Moderate, 51–90 cells per HPF | |
| 3 | Abundant, >90 cells per HPF | |
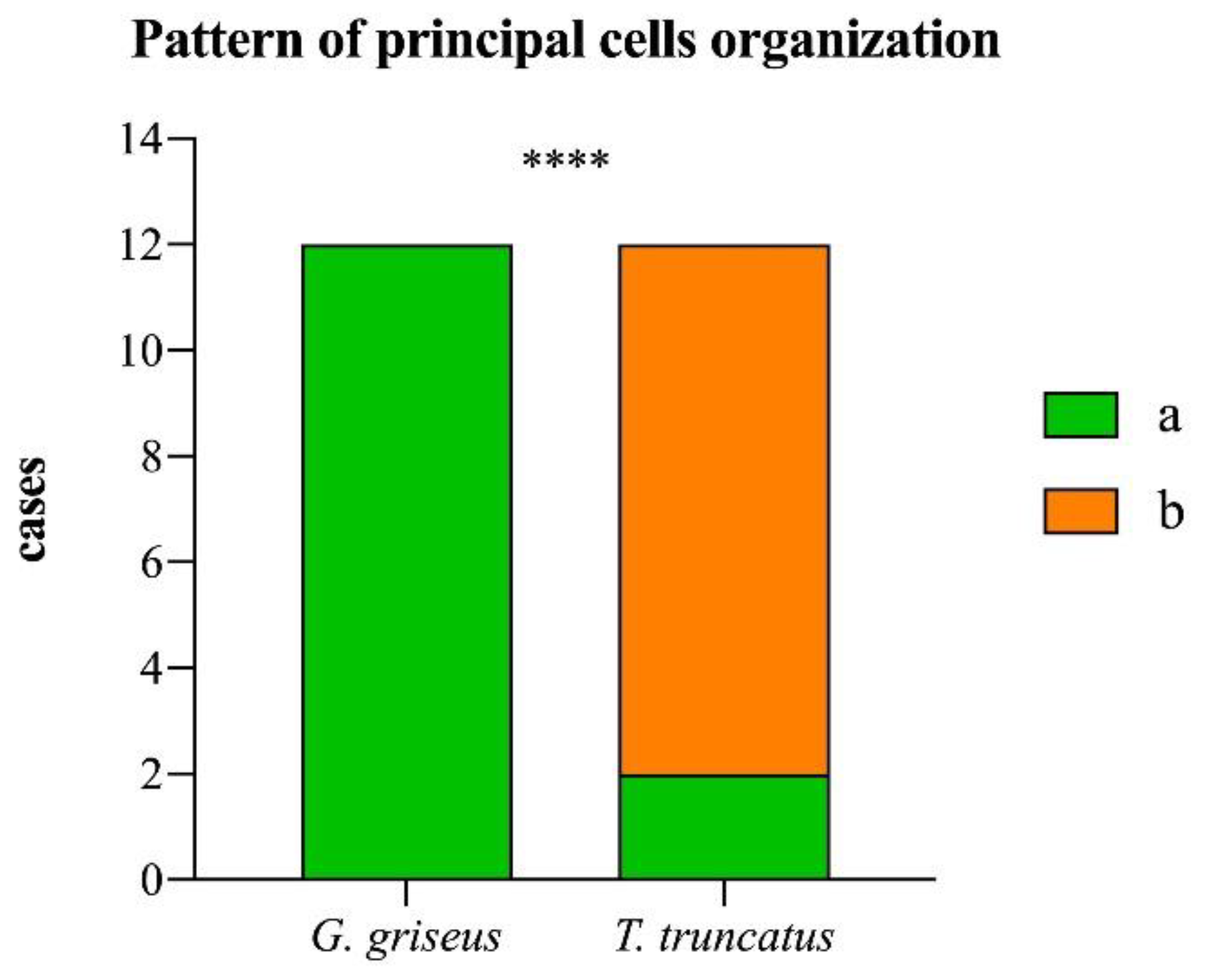
| Pattern of principal cell organization | a | Principal cells well stained with HE, organized in “closed rosettes” |
| b | Principal cells poorly or scantly stained by HE and interspersed between parietal cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biancani, B.; Galosi, L.; Tambella, A.M.; Berardi, S.; Biagini, L.; Mari, S.; Rossi, G. Comparative Study of the Gastric Mucosa of Risso’s Dolphin (Grampus griseus) and Bottlenose Dolphin (Tursiops truncatus): A Key to Manage the Diet in Captive Conditions. Vet. Sci. 2022, 9, 571. https://doi.org/10.3390/vetsci9100571
Biancani B, Galosi L, Tambella AM, Berardi S, Biagini L, Mari S, Rossi G. Comparative Study of the Gastric Mucosa of Risso’s Dolphin (Grampus griseus) and Bottlenose Dolphin (Tursiops truncatus): A Key to Manage the Diet in Captive Conditions. Veterinary Sciences. 2022; 9(10):571. https://doi.org/10.3390/vetsci9100571
Chicago/Turabian StyleBiancani, Barbara, Livio Galosi, Adolfo Maria Tambella, Sara Berardi, Lucia Biagini, Subeide Mari, and Giacomo Rossi. 2022. "Comparative Study of the Gastric Mucosa of Risso’s Dolphin (Grampus griseus) and Bottlenose Dolphin (Tursiops truncatus): A Key to Manage the Diet in Captive Conditions" Veterinary Sciences 9, no. 10: 571. https://doi.org/10.3390/vetsci9100571
APA StyleBiancani, B., Galosi, L., Tambella, A. M., Berardi, S., Biagini, L., Mari, S., & Rossi, G. (2022). Comparative Study of the Gastric Mucosa of Risso’s Dolphin (Grampus griseus) and Bottlenose Dolphin (Tursiops truncatus): A Key to Manage the Diet in Captive Conditions. Veterinary Sciences, 9(10), 571. https://doi.org/10.3390/vetsci9100571