

A Reciprocal Transplant Experiment Confirmed Mite-Resistance in a Honey Bee Population from Uruguay

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Overview
2.2. Estimation of the Bee Population and Brood Area
2.3. Evaluation of Hygienic and Grooming Behaviors
2.4. Estimation of Mites on Bees and in Brood Cells
2.5. Statistical Analysis
3. Results
3.1. Evolution of Colonies Strength
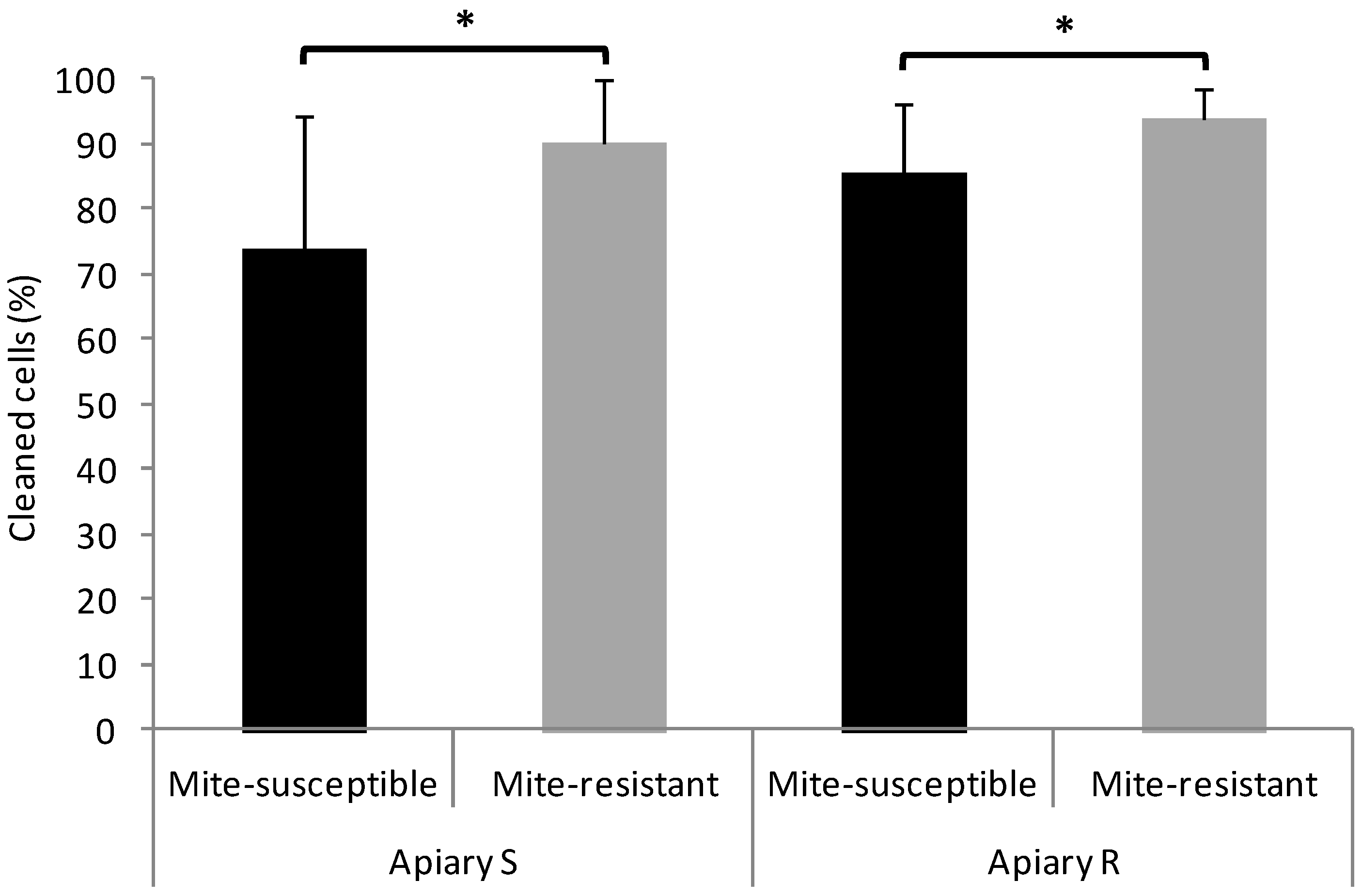
3.2. Hygienic and Grooming Behavior
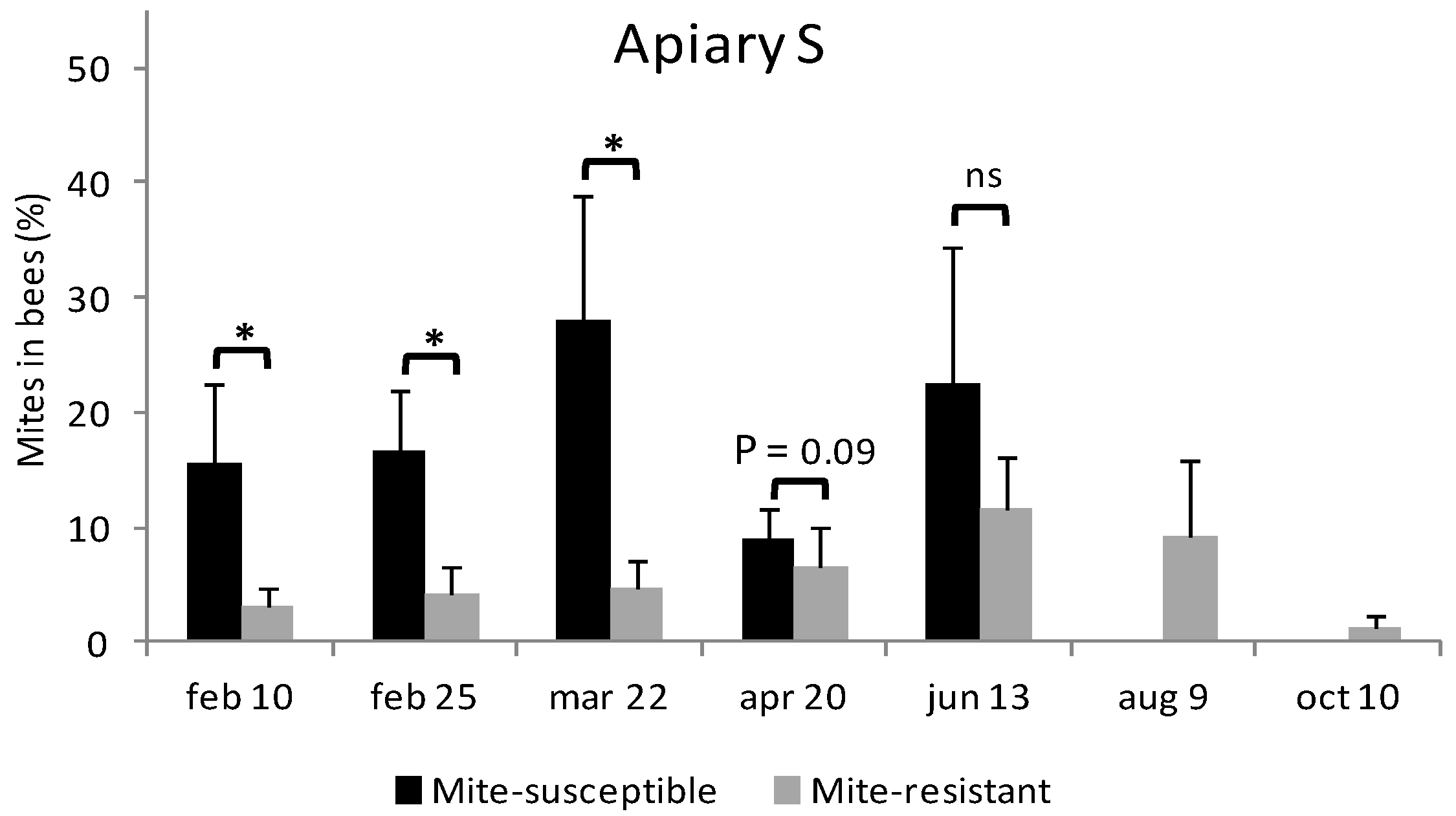
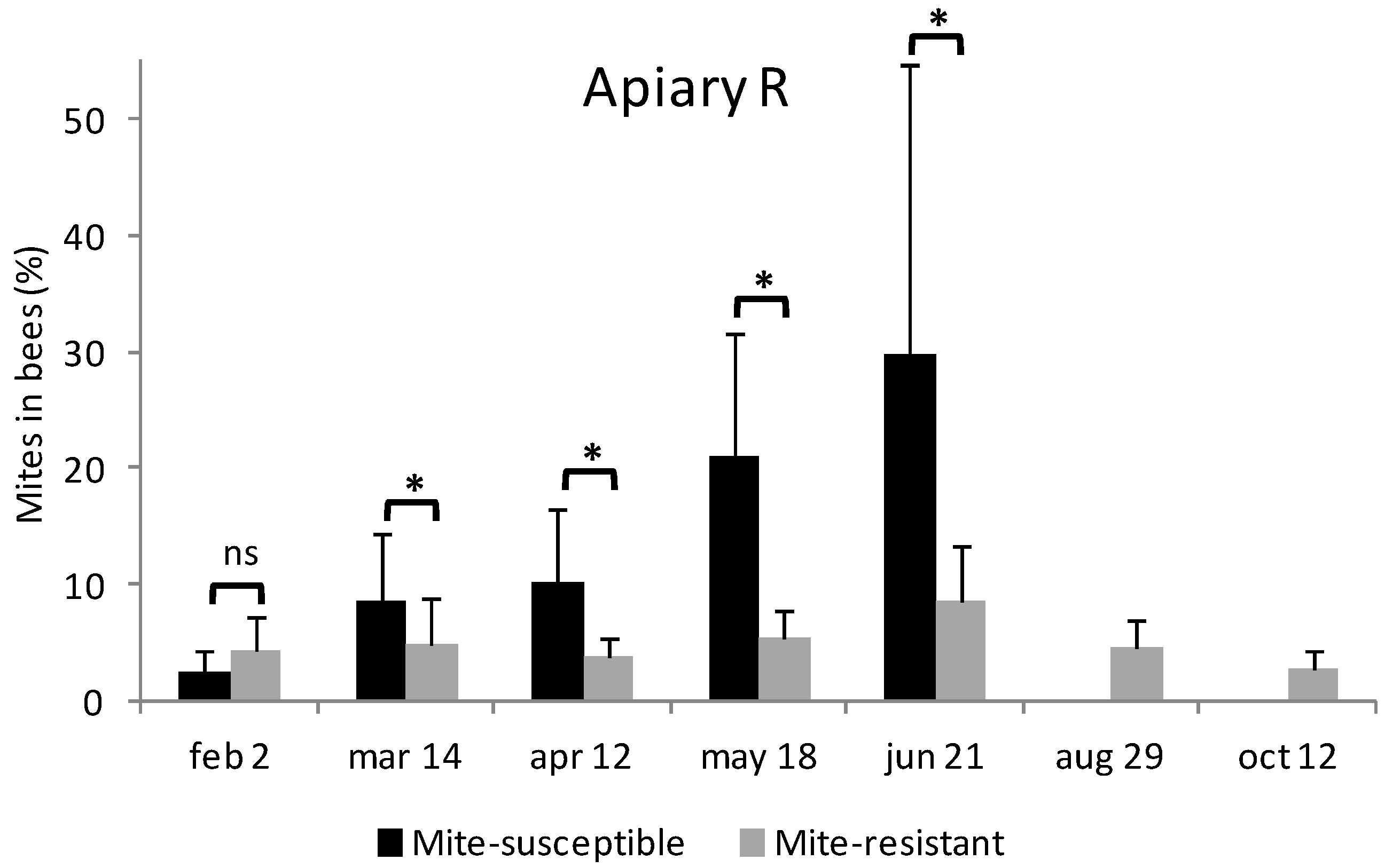
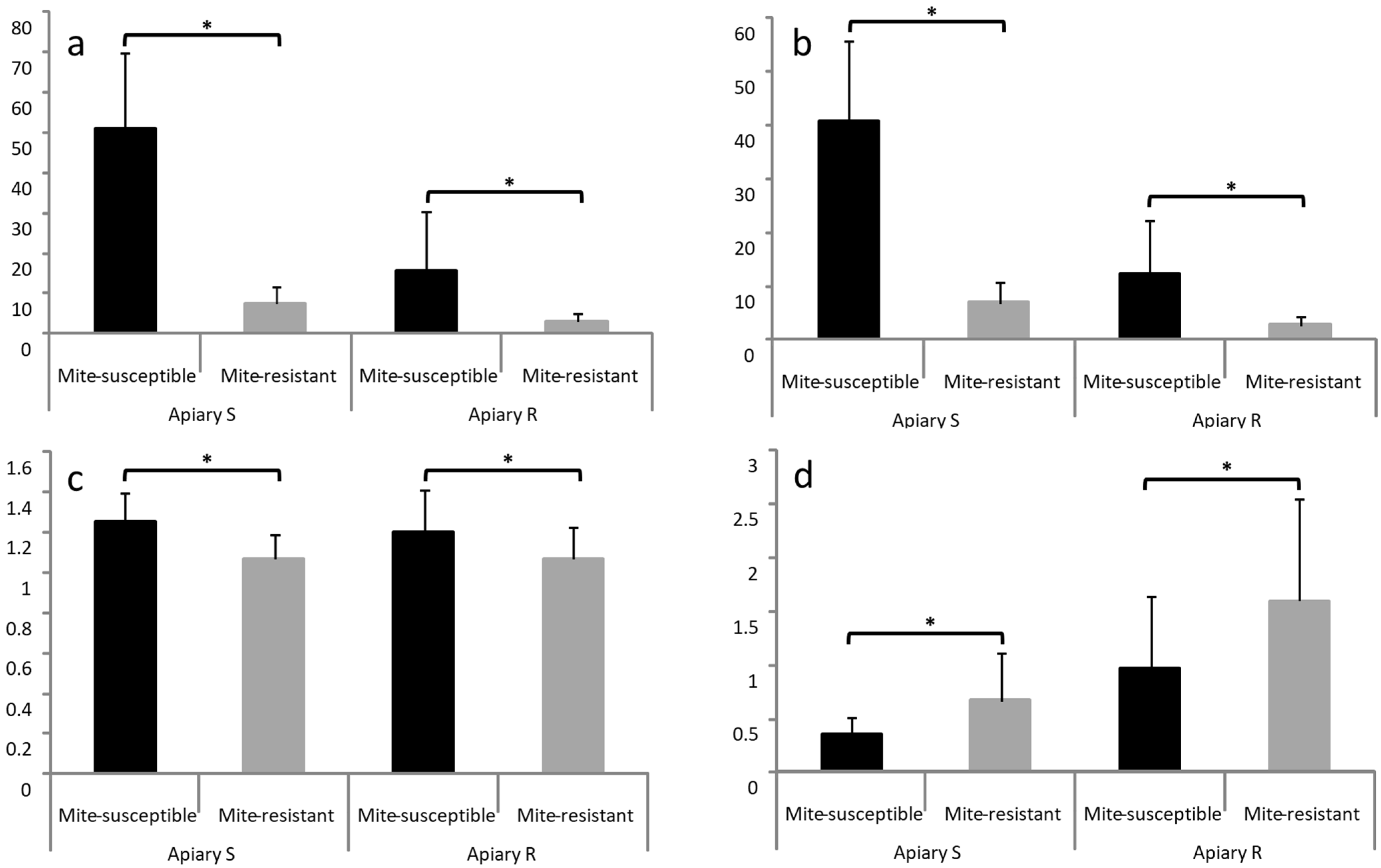
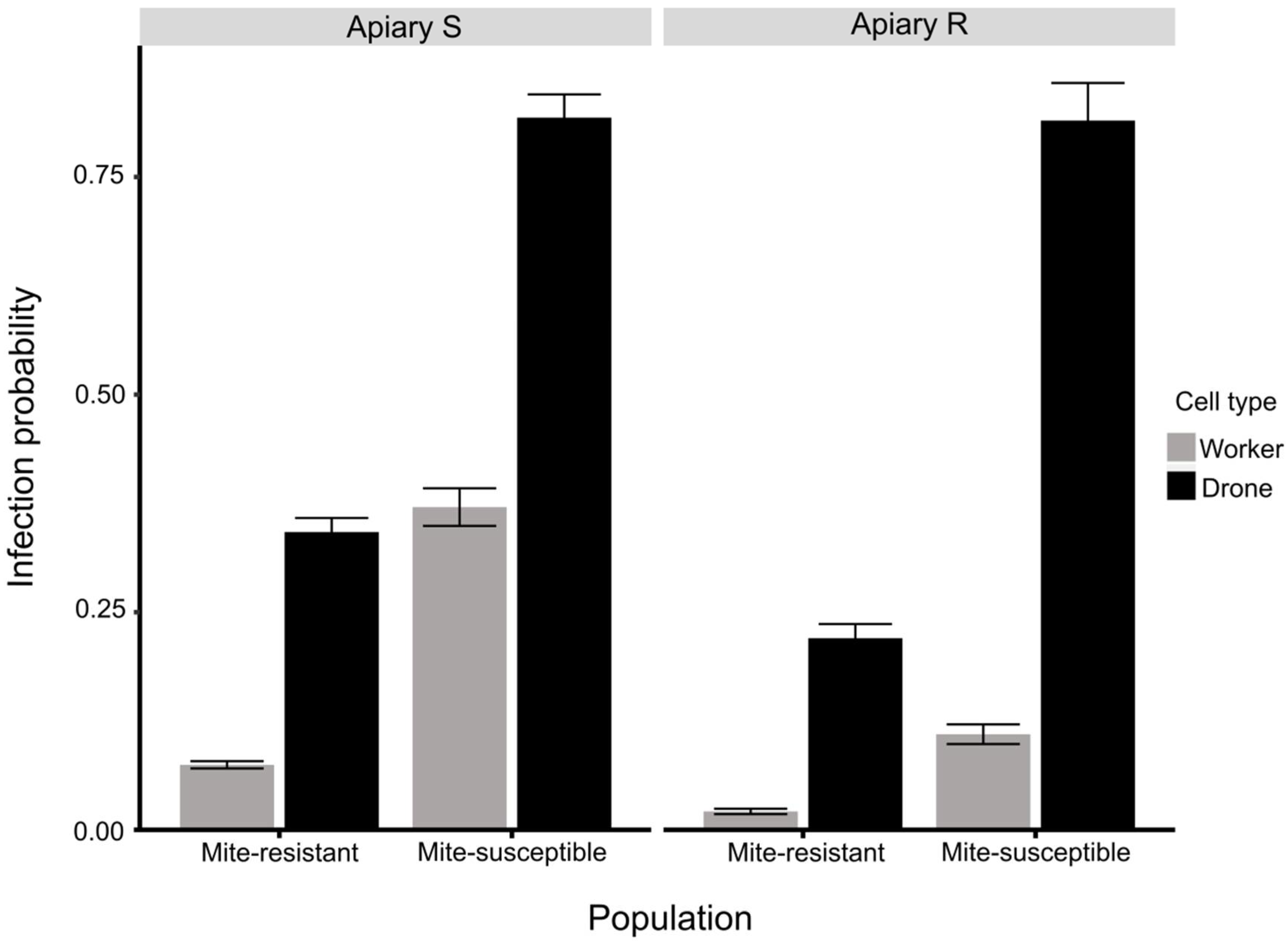
3.3. Estimation of Mites on Bees and in Brood Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa mites and honey bee health: Can Varroa explain part of the colony losses? Apidologie 2010, 41, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaurepaire, A.; Piot, N.; Doublet, V.; Antunez, K.; Campbell, E.; Chantawannakul, P.; Chejanovsky, N.; Gajda, A.; Heerman, M.; Panziera, D.; et al. Diversity and global distribution of viruses of the western honey bee Apis mellifera. Insects 2020, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- de Miranda, J.R.; Genersch, E. Deformed wing virus. J. Invertebr. Pathol. 2010, 103, S48–S61. [Google Scholar] [CrossRef]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.M.; Boots, M. Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Brettell, L.E. Deformed wing virus in honeybees and other insects. Ann. Rev. Virol. 2019, 6, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Maggi, M.; Antúnez, K.; Invernizzi, C.; Aldea, P.; Vargas, M.; Negri, P.; Brasesco, C.; De Jong, D.; Message, D.; Teixeira, E.W.; et al. Honeybee health in South America. Apidologie 2016, 47, 835–854. [Google Scholar] [CrossRef] [Green Version]
- Locke, B. Natural Varroa mite-surviving Apis mellifera honeybee populations. Apidologie 2016, 47, 467–482. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Blacquière, T. The Darwin cure for apiculture? Natural selection and managed honeybee health. Evol. Appl. 2017, 10, 226–230. [Google Scholar] [CrossRef]
- Dynes, T.L.; Berry, J.A.; Delaplane, K.S.; De Roode, J.C.; Brosi, B.J. Assessing virulence of Varroa destructor mites from different honey bee management regimes. Apidologie 2020, 51, 276–289. [Google Scholar] [CrossRef] [Green Version]
- Mondet, F.; Beaurepaire, A.; McAfee, A.; Locke, B.; Alaux, C.; Blanchard, S.; Danka, B.; Conte Le, Y. Honey bee survival mechanisms against the parasite Varroa destructor: A systematic review of phenotypic and genomic research eforts. Int. J. Parasitol. 2020, 50, 433–447. [Google Scholar] [CrossRef]
- Moro, A.; Blacquière, T.; Dahle, B.; Dietemann, V.; Le Conte, Y.; Locke, B.; Neumann, P.; Beaurepaire, A. Adaptive population structure shifts in invasive parasitic mites, Varroa destructor. Ecol. Evol. 2021, 11, 5937–5949. [Google Scholar] [CrossRef] [PubMed]
- Grindrod, I.; Martin, S.J. Parallel evolution of Varroa resistance in honey bees: A common mechanism across continents? Proc. R. Soc. B 2021, 288, 20211375. [Google Scholar] [CrossRef]
- Spivak, M.; Danka, R.G. Perspectives on hygienic behavior in Apis mellifera and other social insects. Apidologie 2021, 52, 1–16. [Google Scholar] [CrossRef]
- Spivak, M. Honey bee hygienic behavior and defense against Varroa jacobsoni. Apidologie 1996, 27, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Spivak, M.; Reuter, G. Performance of hygienic honey bee colonies in a commercial apiary. Apidologie 1998, 29, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Spivak, M.; Reuter, G.S. Varroa jacobsoni infestation in untreated honey bee (Hymenoptera: Apidae) colonies selected for hygienic behavior. J. Econ. Entomol. 2001, 94, 326–331. [Google Scholar] [CrossRef]
- Marcangeli, J.A. Relación entre el comportamiento higiénico de la abeja Apis mellifera (Hymenoptera: Apidae) y el tamaño poblacional del acaro Varroa jacobsoni (Mesostigmata: Varroidae). Nat. Neotrop. 2005, 2, 125–129. [Google Scholar] [CrossRef]
- Ibrahim, A.; Spivak, M. The relationship between hygienic behavior and suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of resistance to Varroa destructor. Apidologie 2006, 37, 31–40. [Google Scholar] [CrossRef]
- Ibrahim, A.; Reuter, G.S.; Spivak, M. Field trial of honey bee colonies bred for mechanisms of resistance against Varroa destructor. Apidologie 2007, 38, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Toufailia, H.M.A.; Amiri, E.; Scandian, L.; Kryger, P.; Ratnieks, F.L. Towards integrated control of varroa: Effect of variation in hygienic behaviour among honey bee colonies on mite population increase and deformed wing virus incidence. J. Apicul. Res. 2014, 53, 555–562. [Google Scholar] [CrossRef]
- Muli, E.; Patch, H.; Frazier, M.; Frazier, J.; Torto, B.; Baumgarten, T.; Kilonzo, J.; Ng’ang’a Kimani, J.; Mumoki, F.; Masiga, D.; et al. Evaluation of the distribution and impacts of parasites, pathogens, and pesticides on honey bee (Apis mellifera) populations in East Africa. PLoS ONE 2014, 9, e94459. [Google Scholar] [CrossRef]
- Mondragón, L.; Spivak, M.; Vandame, R. A multifactorial study of the resistance of honeybees Apis mellifera to the mite Varroa destructor over one year in Mexico. Apidologie 2005, 36, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Leclercq, G.; Blacquière, T.; Gengler, N.; Francis, F. Hygienic removal of freeze-killed brood does not predict Varroa-resistance traits in unselected stocks. J. Apicul. Res. 2018, 57, 292–299. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Suppressed mite reproduction explained by the behaviour of adult bees. J. Apic. Res. 2005, 44, 21–23. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Responses to Varroa by honey bees with different levels of Varroa Sensitive Hygiene. J. Apicul. Res. 2009, 48, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Boecking, O.; Spivak, M. Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidologie 1999, 30, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Arechavaleta-Velasco, M.E.; Guzman-Novoa, E. Relative effect of four characteristics that restrain the population growth of the mite Varroa destructor in honey bee (Apis mellifera) colonies. Apidologie 2001, 32, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Novoa, E.; Emsen, B.; Unger, P.; Espinosa Montaño, L.G.; Petukhova, T. Genotypic variability and relationships between mite infestation levels, mite damage, grooming intensity, and removal of Varroa destructor mites in selected strains of worker honey bees (Apis mellifera L.). J. Invertebr. Pathol. 2012, 110, 314–320. [Google Scholar] [CrossRef]
- Morfin, N.; Given, K.; Evans, M.; Guzmán-Novoa, E.; Hunt, G.J. Grooming behavior and gene expression of the Indiana “mite-biter” honey bee stock. Apidologie 2020, 51, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Harbo, J.R.; Harris, J.W. Selecting honey bees for resistance to Varroa jacobsoni. Apidologie 1999, 30, 183–196. [Google Scholar] [CrossRef]
- Lodesani, M.; Crailsheim, K.; Moritz, R.F.A. Effect of some characters on the population growth of mite Varroa jacobsoni in Apis mellifera L colonies and results of a bi-directional selection. J. Appl. Entomol. 2002, 126, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Locke, B.; Fries, I. Characteristics of honey bee colonies (Apis mellifera) in Sweden surviving Varroa destructor infestation. Apidologie 2011, 42, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.S.; Fang, Y.; Xu, S.; Ge, L. The resistance mechanism of the Asian honey bee, Apis cerana Fabr., to an ectoparasitic mite Varroa jacobsoni Oudemanns. J. Invertebr. Pathol. 1987, 49, 54–60. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Tewarson, N.C.; Singh, A.; Engels, W. Differential hygienic behaviour towards Varroa jacobsoni in capped worker brood of Apis cerana depends on alien scent adhering to the mites. J. Apicult. Res. 1993, 32, 89–93. [Google Scholar] [CrossRef]
- Rath, W. Co-adaptation of Apis cerana Fabr. and Varroa jacobsoni Oud. Apidologie 1999, 30, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, Y.; Tomasco, I.H.; Antúnez, K.; Castelli, L.; Branchiccela, B.; Santos, E.; Invernizzi, C. Unraveling honey bee–Varroa destructor interaction: Multiple factors involved in differential resistance between two Uruguayan populations. Vet. Sci. 2020, 7, 116. [Google Scholar] [CrossRef]
- Delaplane, K.S.; van der Steen, J.; Guzmán-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apicult. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Hatjina, F.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J. Standard methods for rearing and selection of Apis mellifera queens. J. Apicult. Res. 2013, 52, 1–30. [Google Scholar] [CrossRef]
- Invernizzi, C.; Zefferino, I.; Santos, E.; Sánchez, L.; Mendoza, Y. Multilevel assessment of grooming behaviour against Varroa destructor in Italian and Africanized honey bees. J. Apicult. Res. 2016, 54, 1–7. [Google Scholar]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard methods for varroa research. J. Apicult. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. Autoregressive model fitting for control. Ann. Inst. Statist. Math. 1971, 23, 163–180. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Harris, J.W.; Harbo, J.R.; Villa, J.D.; Danka, R.G. Variable population growth of Varroa destructor (Mesostigmata: Varroidae) in colonies of honey bees (Hymenoptera: Apidae) during a 10-year period. Environ. Entomol. 2003, 32, 1305–1312. [Google Scholar] [CrossRef]
- De la Mora, A.; Emsen, B.; Morfin, N.; Borges, D.; Eccles, L.; Kelly, P.G.; Goodwin, P.H.; Guzman-Novoa, E. Selective breeding for low and high Varroa destructor growth in honey bee (Apis mellifera) colonies: Initial results of two generations. Insects 2020, 11, 864. [Google Scholar] [CrossRef]
- Martin, S.; Holland, K.; Murray, M. Non-reproduction in the honeybee mite Varroa jacobsoni. Exp. Appl. Acarol. 1997, 21, 539–549. [Google Scholar] [CrossRef]
- Martin, S.J.; Medina, L.M. Africanized honey bees have unique tolerance to Varroa mites. Trends Parasitol. 2004, 20, 112–114. [Google Scholar] [CrossRef]
- Nganso, B.; Fombong, A.; Yusuf, A.; Pirk, C.; Stuhl, C.; Torto, B. Low fertility, fecundity and numbers of mated female offspring explain the lower reproductive success of the parasitic mite Varroa destructor in African honeybees. Parasitology 2018, 145, 1633–1639. [Google Scholar] [CrossRef]
- Fries, I.; Rosenkranz, P. Number of reproductive cycles of Varroa jacobsoni in honey-bee (Apis mellifera) colonies. Exp. Appl. Acarol. 1996, 20, 103–112. [Google Scholar] [CrossRef]
- Kirrane, M.J.; De Guzman, L.I.; Rinderer, T.E.; Frake, A.M.; Wagnitz, J.; Whelan, P.M. Asynchronous development of honey bee host and Varroa destructor (Mesostigmata: Varroidae) influences reproductive potential of mites. J. Econ. Entomol. 2011, 104, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Oddie, M.A.Y.; Büchler, R.; Dahle, B.; Kovacic, M.; Le Conte, Y.; Locke, B.; de Miranda, J.; Mondet, F.; Neumann, P. Rapid parallel evolution overcomes global honey bee parasite. Sci. Rep. 2018, 8, 7704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Alphen, J.J.M.; Fernhout, B. A small shift in VSH-gene frequency instead of rapid parallel evolution in bees. A comment on Oddie et al. 2018. PeerJ Prepr. 2019, 7, e27938v2. [Google Scholar]
- Martin, S.J.; Hawkins, G.P.; Brettell, L.E.; Reece1, N.; Correia-Oliveira, M.E.; Allsopp, M.H. Varroa destructor reproduction and cell re-capping in mite-resistant Apis mellifera populations. Apidologie 2020, 51, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Amdam, G.V.; Hartfelder, K.; Norberg, K.; Hagen, A.; Omholt, S.W. Altered physiology in worker honey bees (Hymenoptera: Apidae) infested with the mite Varroa destructor (Acari: Varroidae): A factor in colony loss during overwintering? J. Econ. Entomol. 2004, 97, 741–747. [Google Scholar] [CrossRef]
- Aldea, P.; Bozinovic, F. The energetic and survival costs of Varroa parasitism in honeybees. Apidologie 2020, 51, 997–1005. [Google Scholar] [CrossRef]
- Emsen, B.; Hamiduzzaman, M.M.; Goodwin, P.H.; Guzmán-Novoa, E. Lower virus infections in Varroa destructor-infested and uninfested brood and adult honey bees (Apis mellifera) of a low mite population growth colony compared to a high mite population growth colony. PLoS ONE 2015, 10, e0118885. [Google Scholar] [CrossRef] [Green Version]
- Giménez Martínez, P.; Mendoza, Y.; Invernizzi, C.; Fuselli, S.; Alonso Salces, R.; Fernández Iriarte, P.; Maggi, M. Morphometric correlation between Apis mellifera morphotypes (Hymenoptera) and Varroa destructor (Acari) from Uruguay. J. Apicul. Res. 2017, 56, 122–129. [Google Scholar] [CrossRef]
- Beaurepaire, A.; Arredondo, D.; Genchi-García, M.L.; Castelli, L.; Reynaldi, F.J.; Antunez, K.; Invernizzi, C.; Mondet, F.; Le Conte, Y.; Dalmon, A. Genetic diversification of an invasive honey bee ectoparasite across sympatric and allopatric host populations. Infect. Genet. Evol. 2022, 103, 105340. [Google Scholar] [CrossRef]
- Seeley, T.D.; Smith, M.L. Crowding honeybee colonies in apiaries can increase their vulnerability to the deadly ectoparasite Varroa destructor. Apidologie 2015, 46, 716–727. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Apiary S | ||||||||
| Date | 10 Feb. | 25 Feb. | 22 Mar. | 20 Apr. | 13 Jun. | 9 Aug. | 10 Oct. | |
| Population | M-S | 8.9 ± 1.7 a | 7.7 ± 1.3 a | 4.0 ± 1.8 a | 4.1 ± 1.6 a | 1.8 ± 1.3 a | - | - |
| M-R | 9.3 ± 2.2 a | 9.4 ± 1.7 b | 9.5 ± 1.9 b | 9.2 ± 1.8 b | 5.7 ± 2.0 b | 2.4 ± 1.3 | 3.9 ± 2.2 | |
| Brood | M-S | 4.3 ± 0.7 a | 3.8 ± 1.6 a | 2.5 ± 1.5 a | 2.2 ± 0.9 a | 0.1 ± 0.1 a | - | - |
| M-R | 5.0 ± 1.4 a | 5.3 ± 1.1 b | 4.4 ± 1.4 b | 2.2 ± 0.9 a | 0.5 ± 0.5 b | 0.5 ± 0.3 | 2.3 ± 1.5 | |
| Mortality (%) | M-S | 0 a | 0 a | 10 a | 60 a | 75 a | 100 a | 100 a |
| M-R | 0 a | 0 a | 0 a | 0 b | 0 b | 28 b | 61 b | |
| Apiary R | ||||||||
| Date | 2 Fe b. | 14 Mar. | 12 Apr. | 18 May. | 21 Jun. | 29 Aug. | 12 Oct. | |
| Population | M-S | 9.6 ± 1.7 a | 6.7 ± 1.9 a | 7.4 ± 2.8 a | 5.1 ± 2.6 a | 3.6 ± 1.7 a | - | - |
| M-R | 9.7 ± 2.1 a | 7.3 ± 1.3 a | 9.0 ± 1.1 a | 8.4 ± 1.9 b | 6.1 ± 2.1 b | 6.1 ± 2.5 | 8.6 ± 3.0 | |
| Brood | M-S | 5.4 ± 1.3 a | 4.3 ± 1.7 a | 3.2 ± 1.8 a | 1.8 ± 1.6 a | 1.5 ± 1.3 a | - | - |
| M-R | 4.3 ± 0.9 b | 3.7 ± 0.7 a | 3.8 ± 1.1 a | 1.6 ± 0.6 a | 1.0 ± 0.6 a | 2.6 ± 1.2 | 4.2 ± 1.9 | |
| Mortality (%) | M-S | 0 a | 0 a | 5 a | 21 a | 74 a | 100 a | 100 a |
| M-R | 0 a | 5 a | 10 a | 10 a | 10 b | 15 b | 35 b | |
| Colonies | Mite-Susceptible | Mite-Resistant | ||
|---|---|---|---|---|
| Apiary S | Drones | Inspected cells | 209 (6) | 871 (14) |
| Mite-infested cells | 171 | 298 | ||
| Total mites | 497 | 468 | ||
| Abundance | 2.4 | 0.5 | ||
| Intensity | 2.9 | 1.6 | ||
| Prevalence (%) | 81.8 | 34.2 | ||
| Workers | Inspected cells | 499 (6) | 4005 (14) | |
| Mite-infested cells | 185 | 299 | ||
| Total mites | 237 | 327 | ||
| Abundance | 0.5 | 0.1 | ||
| Intensity | 1.28 | 1.09 | ||
| Prevalence (%) | 37.1 | 7.5 | ||
| Mites in drone/worker cells | 5,0 | 6,5 | ||
| Apiary R | Drones | Inspected cells | 81 (3) | 654 (7) |
| Mite-infested cells | 66 | 144 | ||
| Total mites | 143 | 239 | ||
| Abundance | 1.8 | 0.4 | ||
| Intensity | 2.17 | 1.66 | ||
| Prevalence (%) | 81.5 | 22.0 | ||
| Workers | Inspected cells | 765 (3) | 2130 (7) | |
| Mite-infested cells | 84 | 45 | ||
| Total mites | 112 | 46 | ||
| Abundance | 0.1 | 0.02 | ||
| Intensity | 1.33 | 1.02 | ||
| Prevalence (%) | 11.0 | 2.1 | ||
| Mites in drone/worker cells | 12.1 | 16.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendoza, Y.; Santos, E.; Clavijo-Baquett, S.; Invernizzi, C. A Reciprocal Transplant Experiment Confirmed Mite-Resistance in a Honey Bee Population from Uruguay. Vet. Sci. 2022, 9, 596. https://doi.org/10.3390/vetsci9110596
Mendoza Y, Santos E, Clavijo-Baquett S, Invernizzi C. A Reciprocal Transplant Experiment Confirmed Mite-Resistance in a Honey Bee Population from Uruguay. Veterinary Sciences. 2022; 9(11):596. https://doi.org/10.3390/vetsci9110596
Chicago/Turabian StyleMendoza, Yamandú, Estela Santos, Sabrina Clavijo-Baquett, and Ciro Invernizzi. 2022. "A Reciprocal Transplant Experiment Confirmed Mite-Resistance in a Honey Bee Population from Uruguay" Veterinary Sciences 9, no. 11: 596. https://doi.org/10.3390/vetsci9110596
APA StyleMendoza, Y., Santos, E., Clavijo-Baquett, S., & Invernizzi, C. (2022). A Reciprocal Transplant Experiment Confirmed Mite-Resistance in a Honey Bee Population from Uruguay. Veterinary Sciences, 9(11), 596. https://doi.org/10.3390/vetsci9110596