Abstract
APSES (Asm1p, Phd1p, Sok2p, Efg1p, and StuAp) family transcription factors play crucial roles in various biological processes of fungi, however, their functional characterization in phytopathogenic fungi is limited. In this study, we explored the role of SsStuA, a typical APSES transcription factor, in the regulation of cell wall integrity (CWI), sclerotia formation and pathogenicity of Sclerotinia sclerotiorum, which is a globally important plant pathogenic fungus. A deficiency of SsStuA led to abnormal phosphorylation level of SsSmk3, the key gene SsAGM1 for UDP-GlcNAc synthesis was unable to respond to cell wall stress, and decreased tolerance to tebuconazole. In addition, ΔSsStuA was unable to form sclerotia but produced more compound appressoria. Nevertheless, the virulence of ΔSsStuA was significantly reduced due to the deficiency of the invasive hyphal growth and increased susceptibility to hydrogen peroxide. We also revealed that SsStuA could bind to the promoter of catalase family genes which regulate the expression of catalase genes. Furthermore, the level of reactive oxygen species (ROS) accumulation was found to be increased in ΔSsStuA. In summary, SsStuA, as a core transcription factor involved in the CWI pathway and ROS response, is required for vegetative growth, sclerotia formation, fungicide tolerance and the full virulence of S. sclerotiorum.
1. Introduction
Sclerotinia sclerotiorum (Lib.) de Bary, a notorious necrotrophic plant pathogenic fungus, has a complete life cycle and infection cycle with extremely wide distribution, causing blight, stem and crown rots of numerous crops [1,2,3,4]. S. sclerotiorum is widely distributed in warm and arid regions and can infect various plants, including oilseed crops such as soybean and rapeseed, as well as ornamental plants such as marigold and tulip [5]. Polymorphism is a remarkable characteristic of S. sclerotiorum which produces compound appressoria, sclerotia, apothecium and ascospore in the process of morphological differentiation [2,4,6]. Therein, compound appressorium and ascospore are an important infection structure, and sclerotia is the core component of the life and infection cycle of S. sclerotiorum [7].
Sclerotia are produced by S. sclerotiorum under unfavorable external environmental conditions, allowing them to ensure extended periods in the field. When conditions become favorable, sclerotia germinate to produce either hyphae or apothecia. Hyphae can directly infect plant leaves, while ascospores released from apothecia discs infect plant tissues, initiating a new infection cycle [8,9]. Sclerotia serve as critical structures for long-term survival and act as the primary inocula of the disease. Once the sclerotia are formed, chemical control of S. sclerotiorum becomes ineffective [10]. Therefore, the spread of the pathogenic fungi would be significantly diminished if sclerotia formation were to be abolished. The formation and development of sclerotia are mediated by complex internal and external factors, including pH, reactive oxygen species (ROS), and nutrient limitation. S. sclerotiorum relies on multiple signaling pathways, such as cAMP signaling, mitogen-activated protein (MAP) kinase pathway, and autophagy, to respond to and regulate these stimuli [11,12,13]. Furthermore, transcription factors (TFs) serve as vital mediators between signal transduction pathways and target gene expression. Fungal-specific TF families are classified into different types according to their conserved domains, including Zn2/Cys6 (Zn cluster), fungal-specific transcription factor domain, APSES, etc. [14]. Among these, APSES TFs play a pivotal role in the life process of various fungi [15].
The APSES family TFs feature a sequence-specific DNA-binding domain known as the APSES domain, which can adopt a bHLH-like structure [16]. Moreover, it has been proposed that APSES domains may be evolved from KilA-N-like precursors found in viruses that were integrated into host cells during early fungal evolution [17]. Within fungi, APSES family TFs such as Asm1p, Phd1p, Sok2p, Efg1p, and StuAp serve as pivotal regulators of various developmental processes [14,18,19]. For example, AnStuA, the first identified APSES TF, is indispensable for asexual reproduction and conidiophore development in Aspergillus nidulans [15]. In Candida albicans, APSES TFs regulate cell differentiation, morphogenesis, and metabolism [20], while Sok2 and Phd1 regulate pseudohyphal differentiation in Saccharomyces cerevisiae by modulating the expression of Flo11, which is crucial for filamentous growth [21]. In A. fumigatus, StuA mutants display impaired asexual reproduction and abnormal conidiophore morphology [22], and RgdA governs mycelial growth and virulence [23]. Similarly, GcStuA, StuA homolog in Glomerella cingulata, is involved in maintaining appressorium turgor pressure and virulence [24]. The StuA homolog Mstu1 in Magnaporthe oryzae is essential for pathogenicity [25], while deletion of FgStuA in Fusarium graminearum results in reduced spore production, pathogenicity, and secondary metabolite production [26]. MoSwi6 interacts with MoMps1 and is necessary for mycelial growth, conidial morphogenesis, and pathogenicity in M. oryzae [27].
In conclusion, the APSES protein family plays crucial roles in biological processes, such as sporulation, cell differentiation, hyphal growth, secondary metabolism, and virulence [15]. However, the regulatory mechanism of APSES TFs on the growth and development of S. sclerotiorum remains unclear. This study focused on characterizing SsStuA as an APSES family TF, which serves as a pivotal transcription factor regulating normal growth, sclerotia formation, and full virulence in S. sclerotiorum. Studying the function of SsStuA aids in further understanding of the underlying molecular mechanisms that regulate the growth and pathogenicity of S. sclerotiorum. This research also offers valuable insights for the prevention and treatment of the diseases caused by this destructive phytopathogen.
2. Materials and Methods
2.1. Culture Condition of Strains
Sclerotinia sclerotiorum UF-1 was used as the wild type (WT) strain [28]. WT, ΔSsStuA and ΔSsStuA-C strains were routinely grown on potato dextrose agar (PDA) (potato 200 g, glucose 20 g, agar 17 g, constant volume to 1 L) in the dark, at 25 °C constant temperature incubator.
2.2. Identification and Sequence Analysis of SsStuA
Phylogenetic analysis of SsStuA was performed by MEGA version 7.0 software using the neighbor-Joining method with 1000 bootstrap replicates. The conserved domains of SsStuA (XP_001590416.1) and its homologs (Botrytis cinerea B05.10/XP_024548075.1, Colletotrichum siamense/KAF4880364.1, Magnaporthe oryzae 70-15/XP_003718315.1, Aspergillus nidulans FGSC A4/XP_050467081.1, Aspergillus fumigatus Af293/XP_755125.1, Cercospora beticola/XP_023454930.1, Fusarium graminearum PH-1/XP_011319067.1, Ustilago maydis/XP_011388143.1) were analyzed by NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 10 March 2023) and Interpro (http://www.ebi.ac.uk/interpro, accessed on 10 March 2023), and the distribution of the conserved domain was visualized by GPS version 2.0 software [29]. The amino acid sequences of the conserved domain in SsStuA and other fungal species underwent alignment using Clustal W version 1.7 [30].
2.3. SsStuA Gene Knockout and Complementation
The knockout strategy for the SsStuA gene is presented in Figure S1A. Knockout fragments were generated using the SPLIT-MARKER PCR method: the upstream (FR1) and downstream fragments (FR2) of SsStuA were amplified using primers SsStuA F1/R1 and SsStuA F2/R2, respectively. Fragments HY and YG were obtained from the pUCATPH vector using primers M13R/NLC37 and M13F/NLC38 [31,32]. Overlapping fragments were fused using primers SsStuA F1/NLC37 and NLC38/SsStuA R2. These two fragments were then transformed in protoplasts of UF-1 of S. sclerotiorum, as described previously [7,33], and transformants’ hyphal tips were selected using hygromycin (100 μg/mL) at least five times.
Complementary fragments were amplified using primers SsStuA CF1/CR1 and SsStuA CF2/CR2. The two fragments were co-transformed into protoplasts of ΔSsStuA mutants together with the geneticin fragment amplified using primers DW69/DW70 from the pII9 vector. The full SsStuA sequence was amplified from wild-type (WT) genomic DNA with the upstream and downstream regions. Hyphal tips of transformants were selected using geneticin (200 μg/mL). ΔSsStuA and ΔSsStuA-C were confirmed via PCR and quantitative real-time PCR. Details of all primers used in this study are presented in Table S1.
2.4. Plasmid Constructs and Transformation
The vector pNAH-ONG containing a GFP tag was used to see the protein expression in the cell. The SsStuA cDNA sequence was amplified using ONG-SsStuA F/R primers and incorporated into the pNAH-ONG vector, which also contains the neomycin phosphotransferase gene, using the ClonExpress II One Step Cloning Kit (Vazyme Biotech Co., Ltd., Nanjing, China). Subsequently, the constructed vector was transformed into the protoplasts of stains UF-1, and transformants’ hyphal tips were purified through multiple rounds of selection (at least five times) with geneticin (200 μg/mL).
2.5. Stress Treatment
Cell wall synthesis inhibitors including Congo Red (CR, 500 μg/mL), sodium dodecyl sulfate (SDS, 0.01%) and Calcofluor White (CFW, 50 μg/mL) were added to PDA to explore whether deficiency of SsStuA affects cell wall formation in S. sclerotiorum. CM medium was composed of 0.1 g KH2PO4, 0.125 g MgSO4·7H2O, 0.075 g NaCl, 0.5 g Ca(NO3)2·4H2O, 5 g glucose, 0.5 g yeast extract, 0.5 g casein hydrolysate, and 10 g agar per 500 mL. MM medium, a modification of CM, lacked 0.5 g yeast extract and 0.5 g casein hydrolysate. Additionally, MM-N medium was formulated by excluding Ca(NO3)2·4H2O from MM, while MM-C medium omitted glucose. Furthermore, MM-P medium was prepared by eliminating KH2PO4 from MM. These various nutrient-deficient media were used in the study. Agar disks (d = 0.5 cm) of wild-type, ΔSsStuA and ΔSsStuA-C strains were inoculated and the culture dishes were cultivated in the dark, at 25 °C constant temperature incubator, the mycelial growth was measured after 2d inoculation and the inhibition rate was calculated as: inhibition rate = (diameter of untreated − diameter of stress treated)/diameter of untreated × 100%.
2.6. Evaluation of ROS and Oxalic Acid Accumulation
To analyze the effects of oxidative stress, wild-type, ΔSsStuA and ΔSsStuA-C strains were inoculated on PDA medium supplemented with different concentrations of H2O2 (0, 5 and 15 mM). Mycelial growth diameters were measured after 2d inoculation at each concentration of H2O2 repeated three times. 3,3′-Diaminobenzidine Tetrahydrochloride solution (DAB) and nitroblue tetrazolium (NBT) staining solution were used for qualitative analysis of H2O2 and O2− accumulation. Mycelial agar disks (d = 0.5 cm) of wild-type, ΔSsStuA and ΔSsStuA-C strains were placed in a 24-well plate, 1mL DAB (1 mg/mL) was added, and the staining was compared with that after 2d inoculation, then photographed. The mycelia of WT, ΔSsStuA and ΔSsStuA-C strains were stained with NBT (0.5 mg/mL) for 20 min, and the staining was compared and photographed. 100 μg/mL bromophenol blue was added to PDA to detect oxalic acid production and photographed after 2 days of inoculation. If the color of the medium has changed from blue to yellow, it indicates that the strain can produce oxalic acid normally.
2.7. Analysis of Sclerotia and Compound Appressoria Formation of S. sclerotiorum
Compound appressoria of WT, ΔSsStuA and ΔSsStuA-C were observed on glass slides by inoculating agar disks (d = 0.7 cm) and cultivated in a humidifier box at 25 °C incubator. After 48 h incubation, taken pictures and calculated the area of compound appressoria using Image J version 1.50i [29]. The morphology of compound appressoria of WT, ΔSsStuA and ΔSsStuA-C strains on glass slides was observed by microscopy. Onion epidermis was used to observe the penetration efficiency difference between the compound appressoria of wild-type, ΔSsStuA and ΔSsStuA-C strains. The onion epidermis was inoculated with agar disks (d = 0.5 cm) of strains and cultivated in a humidifier box, after 12 h and 24 h incubation, the invasion hypha was stained with lactophenol cotton blue (Sigma, St. Louis, MA, USA) for 2 min, then rinsed with distilled water and observed by DIC microscopy. The morphology, number, and weight of sclerotia were photographed and calculated after 2 weeks incubation.
2.8. Pathogenicity Analysis
Pathogenicity analysis of wild-type, ΔSsStuA and ΔSsStuA-C were analyzed on different hosts. Mycelial agar disks (d = 0.5 cm) were inoculated on the leaves and cultivated in a humidifier box at 25 °C constant temperature incubator. After 48 h inoculation, the areas of lesions on wounded and unwounded leaves were calculated by Image J version 1.50i [29].
2.9. Quantitative Real-Time PCR Analysis
The total RNA of strains was extracted by TransZol Up Plus RNA Kit (TransGen Biotech, Beijing, China). The relative expression of genes was measured by Green qPCR SuperMix (TransGen Biotech, Beijing, China). All the primers used in this article are shown in Table S1 and the name was remarked with “Q”. The qRT-PCR was performed by PrimePro48 (Serial NO. R000100174).
2.10. Protein Extraction and Western Blot
Mycelia of wild-type and ΔSsStuA were cultured in PD for 36 h and supplemented with CR (500 μg/mL) as cell wall stress for 2 h, then collected and ground, protein extraction solution (10 mM Tris-HCl, 150 mM NaCl, 0.5 mM EDTA, 1% Triton X-100, 1 mM PMSF, pH 7.5) 1 mL containing 10 μL protease inhibitor and 10 μL phosphatase inhibitor. The lysate inserted in the ice for 20 min and centrifuged at 12,000 rpm for 15 min at 4 °C. The supernate with equal volumes of 2× loading buffer was separated by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene fluoride membrane. The Phospho-p44/42 MAPK antibody (Cell Signaling Technology, Boston, MA, USA) was used at a 1:3000 dilution for detecting the phosphorylated SsSmk1 and SsSmk3. The second antibody was HRP-labeled goat antirabbit IgG (1:5000) and Coomassie Brilliant Blue (CBB) was used as a control.
2.11. Yeast One-Hybrid
The cDNA sequence of SsStuA was amplified by AD-SsStuAF/AD-SsStuAR and cloned into pGADT7 though ClonExpress II One Step Cloning Kit. The promoters of Sscle_01g011570, Sscle_04g037170, Sscle_15g107280, Sscle_05g044180, Sscle_08g064900, Sscle_15g104430, Sscle_05g047950 and Sscle_03g026200 were amplified and cloned into pHIS. pGADT7-SsStuA and pHIS-X were transformed into yeast Y187 strain. 3-amino-1,2,4-triazole (3-AT, 30 mM) was used to inhibit self-activation of pHIS-X.
3. Results
3.1. SsStuA Is a Critical APSES TF Regulating Sclerotia Formation in S. sclerotiorum
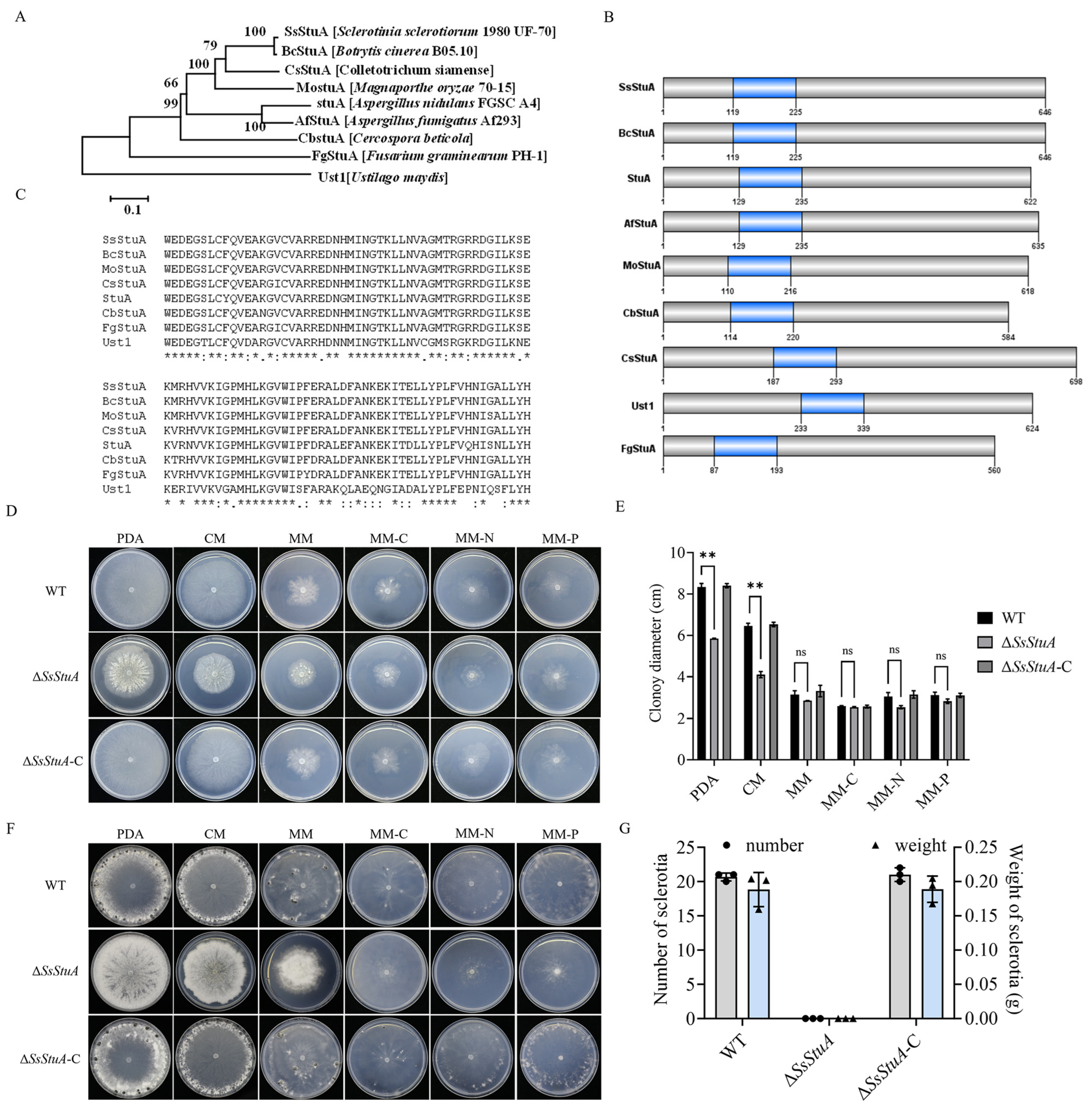
SsStuA (Sscle_10g075100) was grouped together with the Botrytis cinerea BcStuA sequence, which revealed 97.83% similarity (Figure 1A). SsStuA and its homologous proteins from other fungal species contained a conserved domain named KilA-N domain (pfam04383) (Figure 1B,C). In order to clarify the biological function of SsStuA in S. sclerotiorum, split-marker PCR was used to delete SsStuA, and ΔSsStuA was then complemented by transformation with a copy of SsStuA (Figure S1). There were obvious differences in the morphology between UF-1 and ΔSsStuA on PDA (Figure 1D), and the radial growth of ΔSsStuA was reduced (Figure 1E). Significantly, only deep and dense colonies of ΔSsStuA were observed after 14 days of culture on PDA, CM, and MM, indicating that the deficiency of SsStuA abolishes the formation of sclerotia (Figure 1F,G). The results indicate that SsStuA performs a vital function in the vegetative growth and sclerotia formation of S. sclerotiorum.
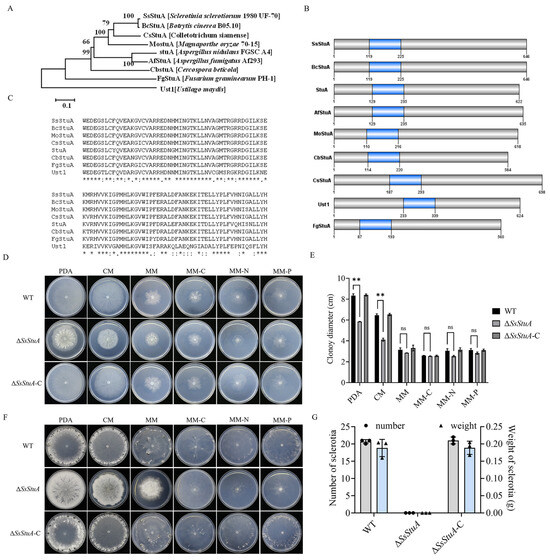
Figure 1.
SsStuA performed a critical role in vegetative growth and sclerotia formation of S. sclerotiorum. (A) Phylogenetic analysis of SsStuA. The sequences of SsStuA and its homologs were aligned by ClustalW, and MEGA 7 was used to construct the phylogenetic tree by using the neighbor-Joining method with 1000 bootstrap replicates. (B) Analysis of conserved domains of SsStuA. Conserved domains were analyzed by interpro and visualized by GPS 2.0. (C) The conserved sites in the domain of SsStuA. The conserved sites were labeled with an asterisk “*”. (D) Colony morphology of ΔSsStuA, WT and ΔSsStuA-C. (E) Colony diameters were measured after 48 h incubation. (F,G) Sclerotia formation of ΔSsStuA, WT and ΔSsStuA-C (Gray and blue represent the number and weight of sclerotia in (G), respectively). Error bars mean standard deviations (SDs) and significant differences between the WT and ΔSsStuA were performed by t test (**, p < 0.01), ns means that there was no significance between WT and ΔSsStuA.
3.2. SsStuA Is Involved in Cell Wall Integrity of S. sclerotiorum
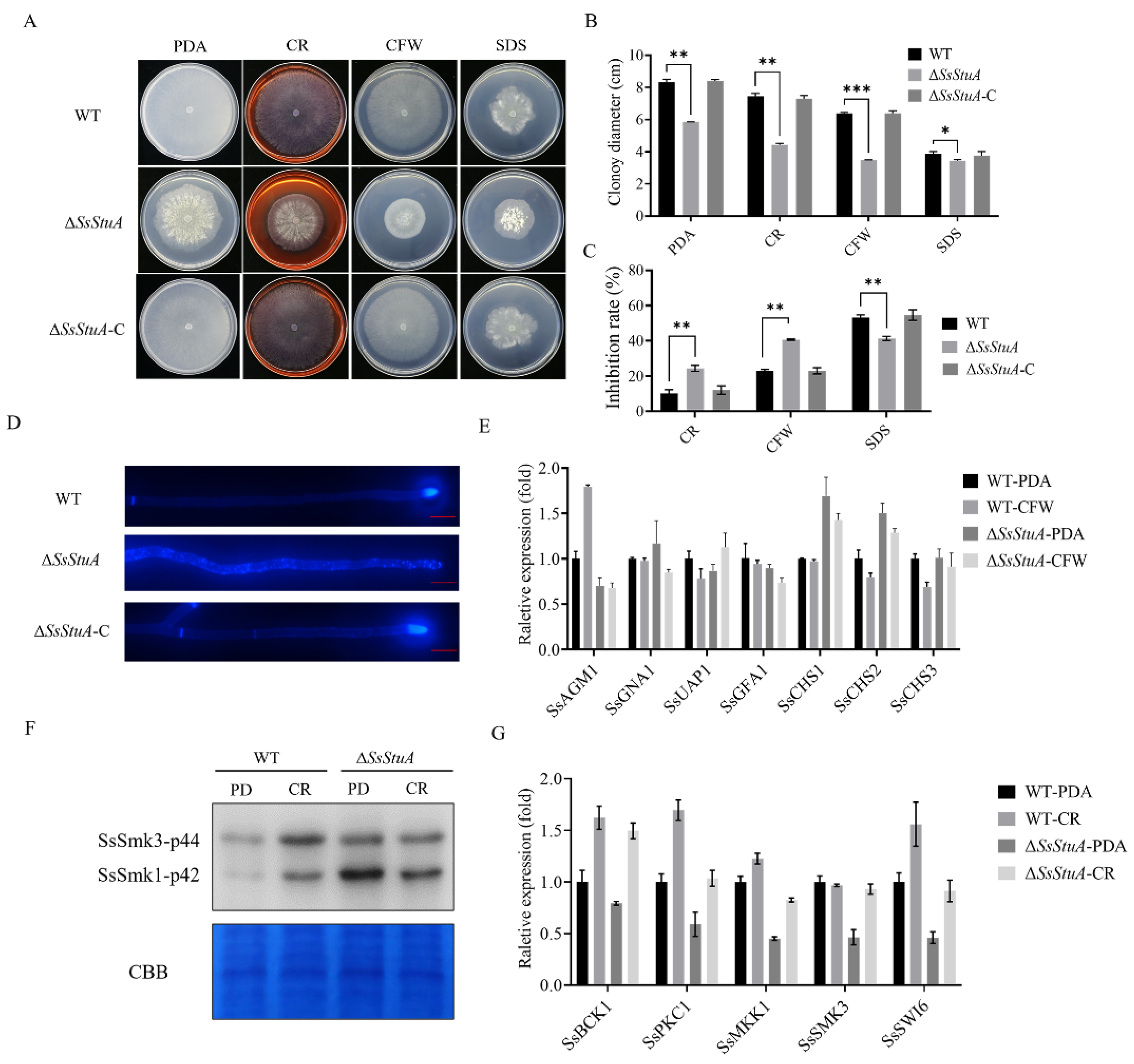
Since ΔSsStuA showed a compact colony on PDA, we suspected that SsStuA might affect the cell wall integrity. The CWI (cell wall integrity) signaling pathway provides a pivotal role in responding to diverse stress in pathogenic fungi [34,35]. Cell wall inhibitors (CR and CFW) limited the growth of ΔSsStuA (Figure 2A–C) and CFW staining hyphae exhibited scattered and distributed blue fluorescence (Figure 2D), which suggest that SsStuA is involved in cell wall integrity of S. sclerotiorum. UDP-GlcNAc is the precursor of chitin which is the main component of the fungal cell wall. Therefore, we assessed the expression level of SsAGM1 under CFW treatment. SsAGM1 responded to CFW treatment and was up-regulated in WT, but there was no significant change in ΔSsStuA (Figure 2E). These results suggest that the chitin synthesis process in ΔSsStuA is affected. The genes associated with chitin synthesis (SsCHS1 and SsCHS2) exhibited significantly elevated expression levels in ΔSsStuA (Figure 2E), potentially accounting for the dense colony phenotype in ΔSsStuA.
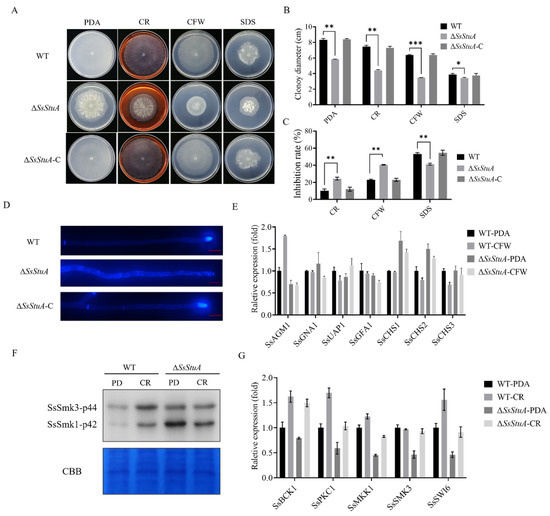
Figure 2.
SsStuA contributes to cell integrity of S. sclerotiorum. (A) Colony morphology of ΔSsStuA, WT and ΔSsStuA-C under different cell wall inhibitors. (B) Colony diameters of ΔSsStuA, WT and ΔSsStuA-C. (C) The inhibition rate of hyphal growth under diverse cell wall inhibitors. Inhibition rate = (diameter of untreated − diameter of stress treated)/(diameter of untreated × 100%). (D) Deficiency of SsStuA caused defective cell wall. ΔSsStuA, WT and ΔSsStuA-C were stained with Calcofluor White (CFW) and observed by fluorescence microscope. Scale bar = 50 μm. (E) Relative expression of genes involved in UDP-GlcNAc synthesis pathway and chitin synthesis. (F) Phosphorylation level of SsSmk3 in WT and ΔSsStuA under CR treatment. (G) Relative expression of CWI pathway genes in WT and ΔSsStuA under CR treatment. Error bars represent the SDs and significant differences between the WT and ΔSsStuA were performed by t test (*, p < 0.05; **, p < 0.01, ***, p < 0.001).
Moreover, the cell wall integrity pathway is regulated by MAPK-SsSmk3. After treatment with CR, the phosphorylation level of SsSmk3 significantly increased in wild type, but there was no significant change in ΔSsStuA (Figure 2F). The expression levels of CWI pathway related genes (SsPKC1, SsMKK1, SsSMK3 and SsSWI6) in ΔSsStuA were decreased remarkably compared with that of WT (Figure 2G). In summary, transcription factor SsStuA is involved in the CWI pathway.
3.3. ΔSsStuA Is Sensitive to Tebuconazole
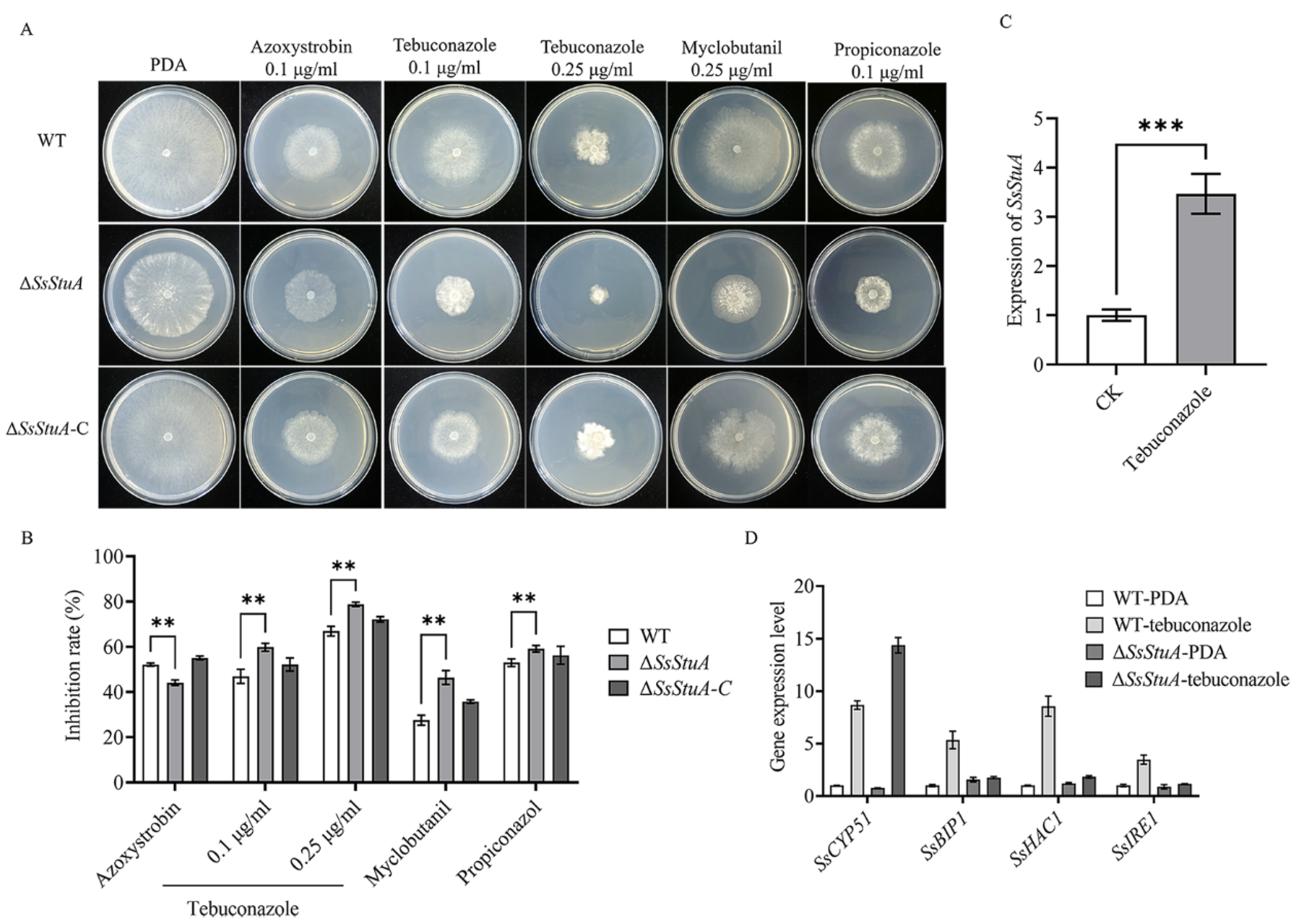
Tebuconazole is a triazole fungicide belonging to DMI class, which is an ergosterol inhibitor of phytopathogens that mainly affects the formation of fungal cell wall [36,37]. The deletion of SsStuA caused defects in the CWI pathway, which led to the question of whether ΔSsStuA was sensitive to fungicides. Thus, different fungicides were added to explore whether the SsStuA affects the fungicide resistance of S. sclerotiorum. The inhibition rate of tebuconazole, myclobutanil and propiconazole was all increased in ΔSsStuA (Figure 3A,B), and the expression level of SsStuA also increased under tebuconazole treatment (Figure 3C). CYP51 is the target of tebuconazole [38,39], therefore, the expression of SsCYP51 was measured in WT and ΔSsStuA. The results showed that SsCYP51 was up-regulated in ΔSsStuA, which indicate that SsCYP51 is not the only target of tebuconazole in S. sclerotiorum (Figure 3D). Tebuconazole triggers endoplasmic reticulum (ER) stress by activating the unfolded protein response (UPR) [40]. To determine whether SsStuA is important for the UPR, the expression levels of SsBIP1, SsHAC1 and SsIRE1 were measured. Compared to WT, the deficiency of SsStuA caused no obvious change of SsBIP1, SsHAC1 and SsIRE1 under tebuconazole treatment (Figure 3D), demonstrating that SsStuA is essential for the UPR.
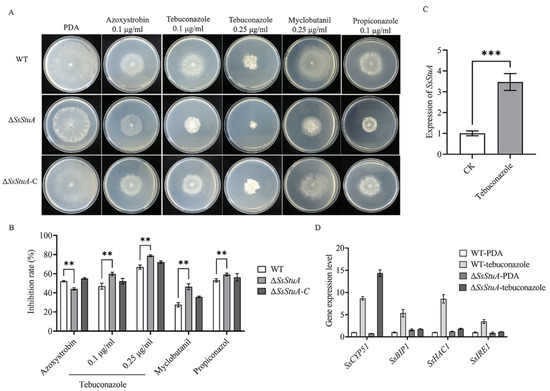
Figure 3.
ΔSsStuA increased sensitivity to tebuconazole. (A) Sensitivity of ΔSsStuA to fungicides. (B) Inhibition rate of fungicides on the ΔSsStuA. Inhibition rate = (diameter of untreated − diameter of fungicide treated)/(diameter of untreated × 100%). (C) The expression of SsStuA under tebuconazole treatment. (D) Gene expression of WT and ΔSsStuA under tebuconazole treatment. Error bars represent the SDs and significant differences were performed by t test (**, p < 0.01, ***, p < 0.001).
3.4. ΔSsStuA Mutant Shows Attenuation of Pathogenicity
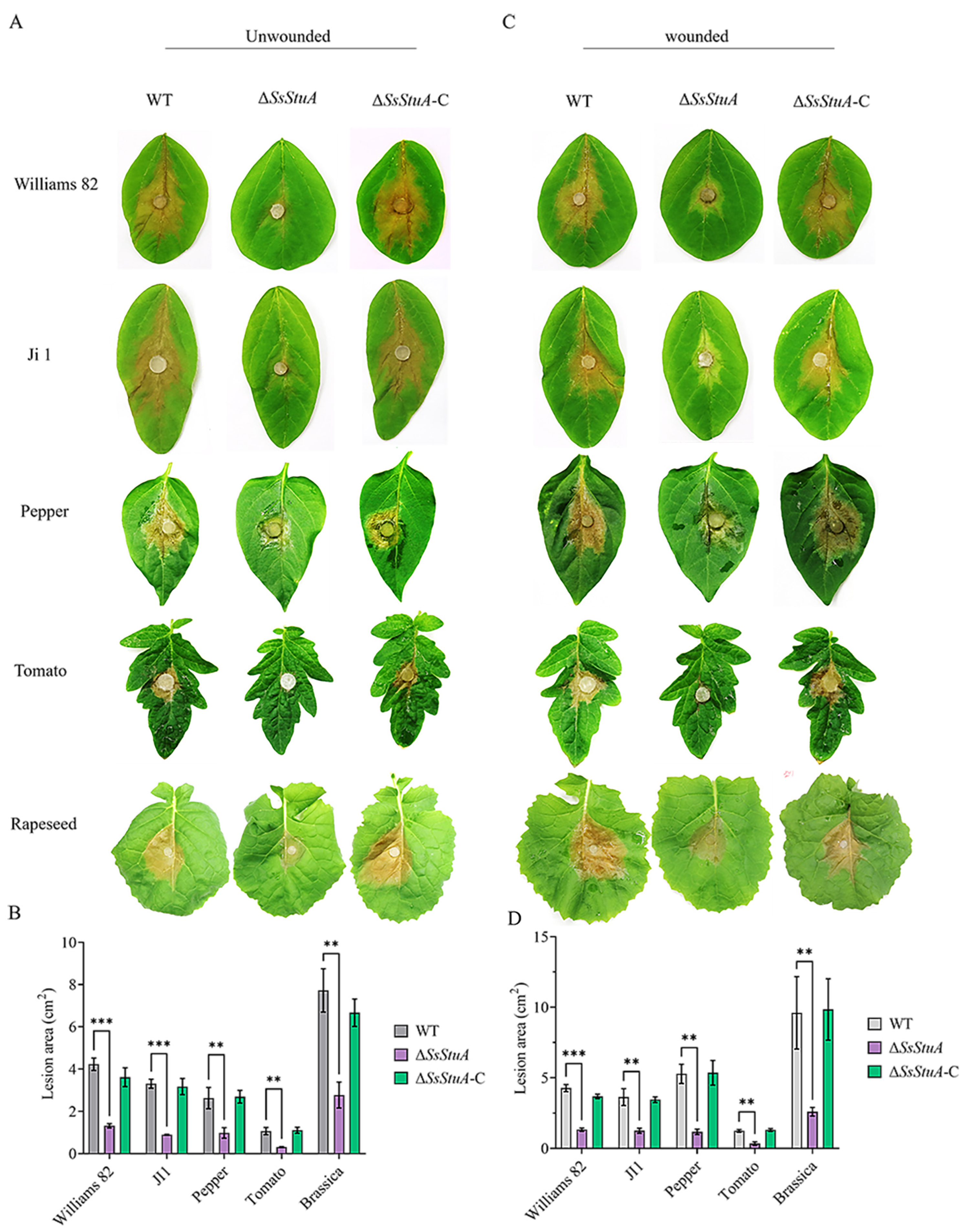
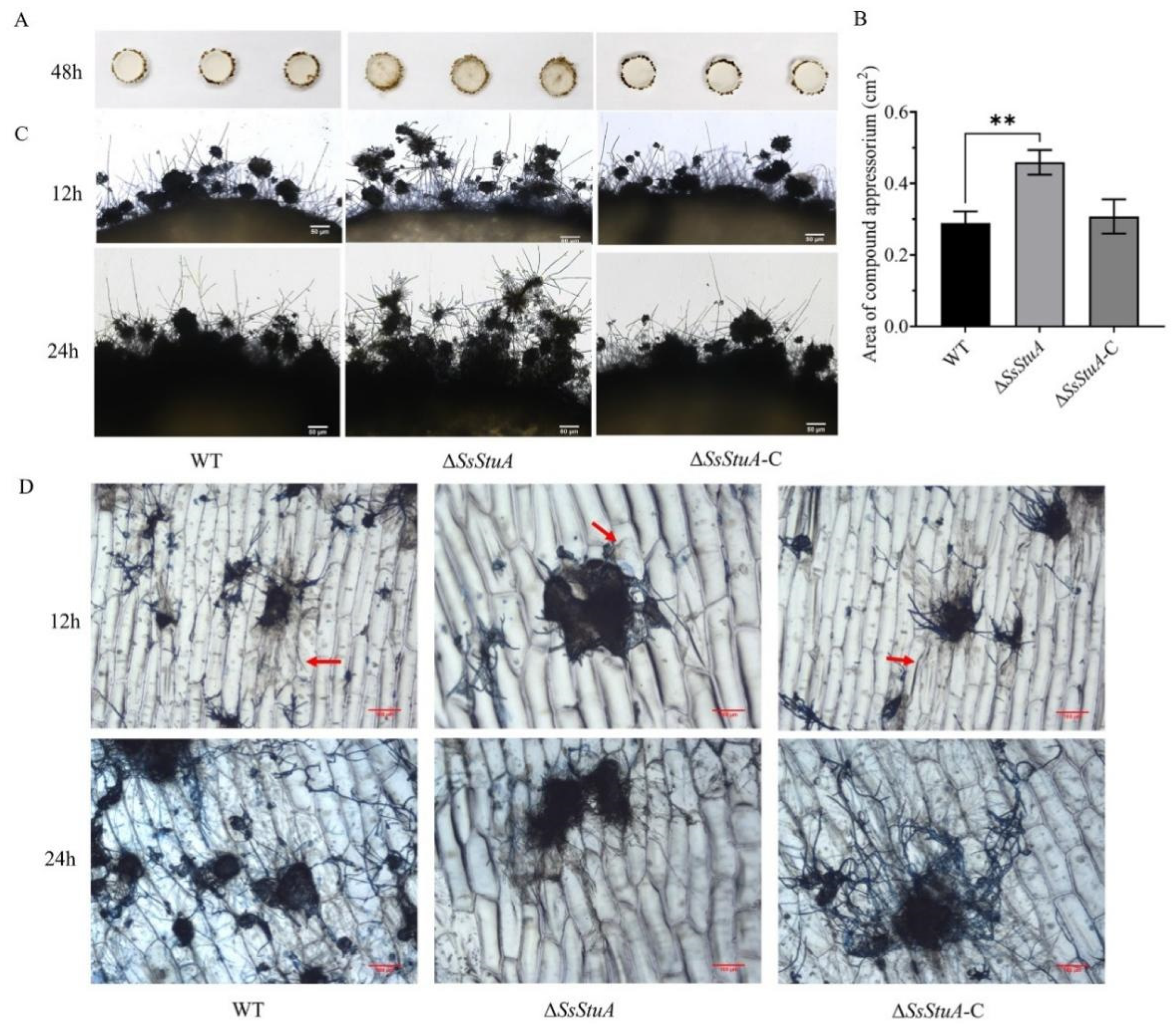
The pathogenicity of ΔSsStuA was obviously decreased on unwounded leaves (Figure 4A,B), ΔSsStuA inoculated on wounded leaves also caused defects in pathogenicity (Figure 4C,D). Compound appressorium is a prerequisite for successful penetration of the host organism by S. sclerotiorum. Therefore, the formation and development of compound appressoria were investigated on glass slide, and obviously numerous compound appressoria were formed from the agar disk of ΔSsStuA (Figure 5A,B). We further observed the morphology of compound appressoria, but there was no distinction between ΔSsStuA and WT (Figure 5C). In order to compare the penetration capacity of compound appressoria, WT, ΔSsStuA and ΔSsStuA-C were inoculated on onion epidermis respectively and stained with lactophenol cotton blue to observe the growth situation of the invasive hyphae (IH). The unstained IH of ΔSsStuA showed retarded growth than that of WT and ΔSsStuA-C (Figure 5D). The results indicate that SsStuA probably negatively regulates the formation of compound appressoria but positively regulates the growth of IH in plant tissue, and the deletion of SsStuA perhaps affects the effects of other pathogenic factors besides compound appressoria.
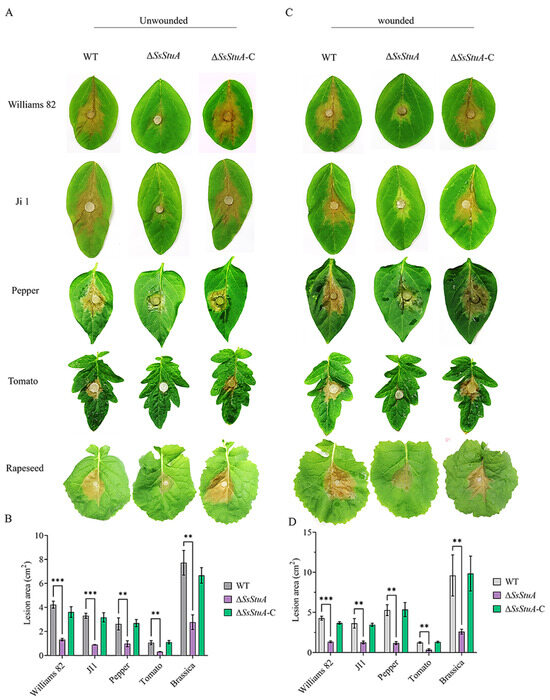
Figure 4.
SsStuA influenced the full virulence of S. sclerotiorum. (A) Unwounded inoculation of ΔSsStuA, WT, and ΔSsStuA-C on different host plants. (B) Analysis of the lesion area under unwounded inoculation. (C) Wounded inoculation of ΔSsStuA, WT, and ΔSsStuA-C on different host plants. (D) Analysis of the lesion area under wounded inoculation. The lesion area was calculated by Image J. Error bars represent the SDs and significant differences between the WT and ΔSsStuA were performed by t test (**, p < 0.01, ***, p < 0.001).
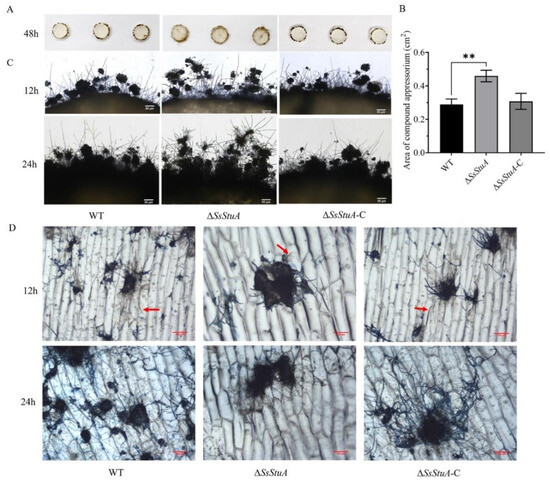
Figure 5.
Compound appressoria were abnormal in ΔSsStuA. (A) Compound appressoria production of strains on glass slides. Photographed at 2 days post-inoculation. (B) The area of compound appressoria produced by ΔSsStuA was larger than that of WT and ΔSsStuA-C. Area of compound appressoria was calculated by Image J. Error bars represent the SDs and significant difference was performed by t test (**, p < 0.01). (C) Morphology of compound appressoria on glass slides. Scale bar = 50 μm. (D) The invasive hyphae of ΔSsStuA showed retarded growth in onion epidermis. The compound appressoria were stained in blue by lactophenol blue, the red arrow points to the invasive hyphae. Scale bar = 100 μm.
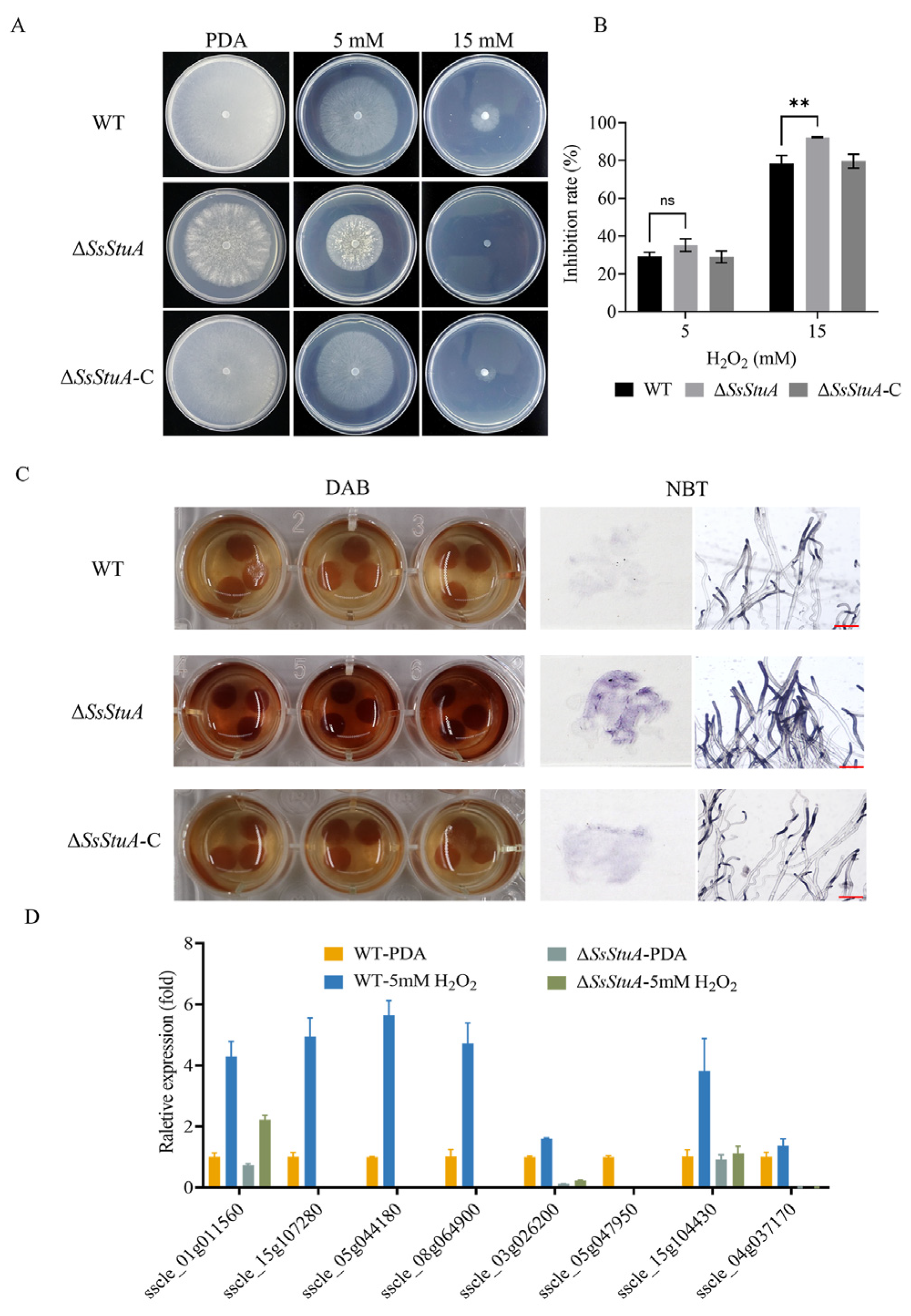
3.5. ΔSsStuA Was Hypersusceptible to Hydrogen Peroxide
The development of invasive hyphae in host tissue could respond to complex signals. Hypersensitive reaction is one of the earliest responses of plants to pathogens, which could accumulate a lot of reactive oxygen species [41]. To determine the mechanism of the reduced virulence of ΔSsStuA, we examined the susceptibility of ΔSsStuA to hydrogen peroxide. ΔSsStuA was more hypersusceptible to hydrogen peroxide than that of WT and ΔSsStuA-C strains (Figure 6A,B). Subsequently, we used DAB and NBT staining to detect ROS production during hyphal growth in ΔSsStuA. Compared with WT strain, the dark-brown coloration and dark-blue formazan precipitates of ΔSsStuA suggest an increased accumulation of ROS in ΔSsStuA under the same growth condition (Figure 6C). Moreover, the expression of catalase family genes was dramatically decreased in ΔSsStuA (Figure 6D), indicating that SsStuA is required for the normal expression of genes associated with reactive oxygen scavenging enzymes.
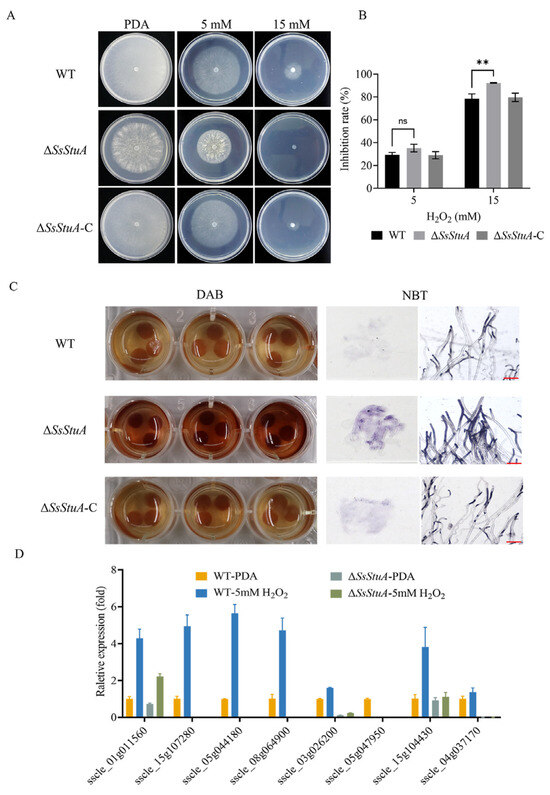
Figure 6.
ΔSsStuA was hypersusceptible to H2O2. (A) The morphological phenotype of WT, ΔSsStuA and ΔSsStuA-C in PDA medium supplemented with 0, 5 and 15 mM H2O2 concentrations. (B) The hyphal growth of WT, ΔSsStuA and ΔSsStuA-C at 0, 5 and 15 mM H2O2 concentrations. Colony diameters were measured at 2 days post-inoculation. (C) Detection of ROS accumulation in hyphae of strains. DAB and NBT were used to detect ROS accumulation. Scale bar = 50 μm. (D) The relative gene expression of CAT family genes in WT and ΔSsStuA. Error bars represent the SDs and significant difference was performed by t test (**, p < 0.01), ns means that there was no significant difference between WT and ΔSsStuA.
3.6. SsStuA Transcriptional Activation of Catalase Gene Expression
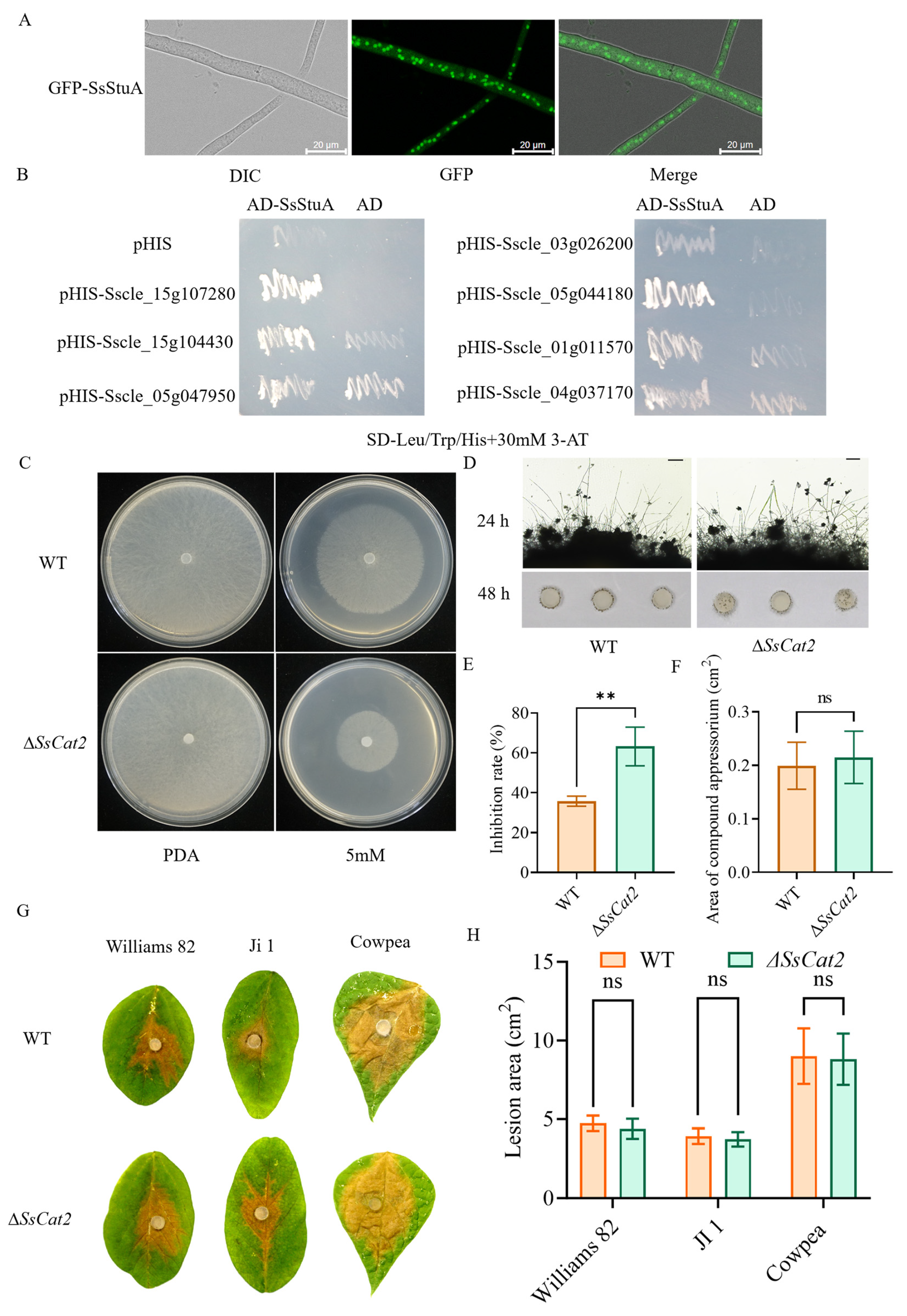
SsStuA was observed to be localized in the nucleus (Figure 7A). Furthermore, there was a notable increase in the accumulation of reactive oxygen species (ROS) in ΔSsStuA, coupled with a down-regulation of ROS metabolism genes. A yeast one-hybrid (Y1H) experiment was performed to elucidate the reason behind the significant down-regulation of catalase (CAT) family genes in ΔSsStuA. The yeast strains harboring pGADT7-SsStuA and pHIS-X (Sscle_01g011570, Sscle_04g037170, Sscle_15g107280, Sscle_05g044180, Sscle_15g104430, Sscle_05g047950, and Sscle_03g026200) exhibited normal growth on SD-Leu/Trp/His medium supplemented with 30 mM 3-AT, while the strains containing only pGADT7 and pHIS-X failed to survive (Figure 7B). This Y1H result preliminarily confirms that SsStuA possesses transcriptional activation activity and can bind to the promoters of CAT family genes. In order to determine whether CAT genes participated in the active oxygen metabolism of S. sclerotiorum, the SsCat2 gene was knocked out. Further analysis of hydrogen peroxide sensitivity and pathogenicity showed that after SsCat2 deletion, the sensitivity to hydrogen peroxide increased (Figure 7C,E), however, the compound appressoria and pathogenicity of ΔSsCat2 had no obvious difference with WT (Figure 7D,F–H). The results indicate that the transcription of CAT genes is regulated by SsStuA and the function of CAT family genes is redundant.
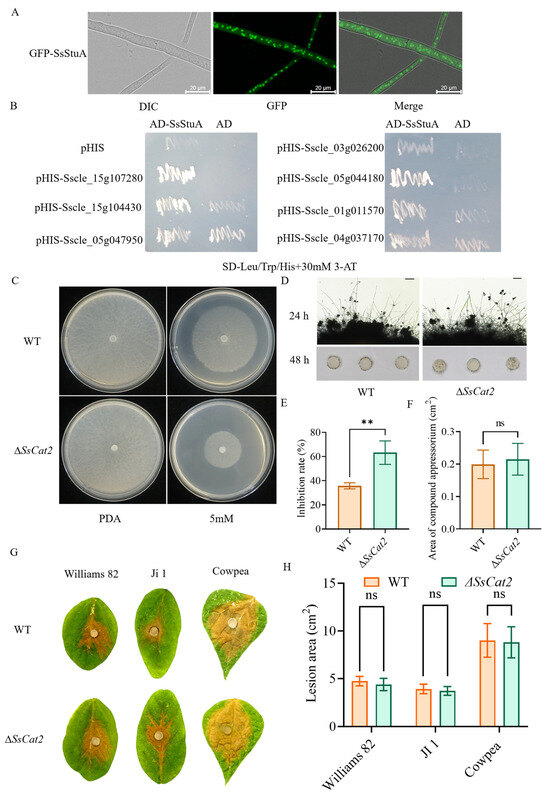
Figure 7.
SsStuA transcriptionally activates catalase genes expression. (A) GFP-SsStuA was localized in the nucleus of S. sclerotiorum. (B) SsStuA binds to the promoters of the CAT family genes. (C,E) ΔSsCat2 was hypersusceptible to H2O2. (D,F) The compound appressoria of ΔSsCat2 did not differ from the wild type. Scale bar = 50 μm. (G,H) Pathogenicity between WT and ΔSsCat2 does not have any obvious differences. Error bars represent the SDs and significant difference was performed by t test (**, p < 0.01), ns means that there was no significant difference between WT and ΔSsCat2.
4. Discussion
Transcription factors (TFs) play a crucial role in signal transduction, serving as the vital link between signal reception and the expression of target genes. They regulate gene expression within cells, ultimately influencing cell function [14]. StuA belongs to the APSES protein family, which is unique to fungi and has been identified as a key regulator of cellular development and other biological processes [15,16,42]. Here, we found that SsStuA as a core transcription factor, plays vital roles in regulating CWI pathway, ROS response, tolerance to tebuconazole, sclerotia formation and the full virulence of S. sclerotiorum.
Sclerotia, as the initial infection source of stem rot produced by S. sclerotiorum, play an important role during the disease cycle [4,8]. We reported that the formation of sclerotia was abolished, and the colony showed a deeper color with increasing culture time after SsStuA deletion. Comparably, in Verticillium dahliae, Vst1 is involved in the production of microsclerotia [42]. In A. nidulans, StuA was required for correct differentiation and spatial organization of cell pattern in the complex conidiophore [43,44], and MrStuA regulated sporulation as well as vegetative growth and virulence in Metarhizium robertsii [45]. In the current study, we demonstrated that SsStuA plays an indispensable role in the asexual structure formation of S. sclerotiorum, indicating its function is conserved in ascomycetes.
Compound appressoria and Oxalic acid (OA) are the important factors that affect the full virulence of S. sclerotiorum [10,46,47]. Interestingly, the mechanism of sclerotia development is obviously opposite to that of compound appressoria in ΔSsStuA. ΔSsStuA produced more compound appressoria than the wild type and generated oxalic acid normally (Figure S2). However, the pathogenicity of ΔSsStuA was obviously decreased. Analyzing the penetrability of the compound appressorium between ΔSsStuA and WT, the IH of ΔSsStuA were grew slowly, which made the lesion area of the SsStuA mutant significantly lower than that of the wild type and ΔSsStuA-C. These results indicate that compound appressorium formation does not always dominate the pathogenicity. Furthermore, StuA had some distinctions between different ascomycetes. Mstu1 did not affect the number of appressoria but was deficient in the appressorium of Magnaporthe grisea [25]. Vst1 is dispensable for pathogenicity in V. dahliae and V. nonalfalfae [42]. Additionally, the pathogenicity of phytopathogenic fungus is also related to the external environment. Plants produce reactive oxygen species after inoculation with pathogens [48,49]. ΔSsStuA was more sensitive to reactive oxygen species than wild type and ΔSsStuA-C. The results further indicate that SsStuA could bind to the promoters of the CAT family genes and regulate the expression level of the CAT family genes. However, deficiency of SsCAT2 gene does not influence the full virulence of S. sclerotiorum, further suggesting that the influence of SsStuA on pathogenicity is multifactorial.
The prevention and control of sclerotinia disease caused by S. sclerotiorum was still challenging. Results showed that ΔSsStuA was sensitive to tebuconazole, which is an ergosterol inhibitor that affects the formation of fungal cell wall by inhibiting the demethylation reaction of ergosterol intermediates. Cell wall integrity provides a pivotal role in the response and adaptation to various stresses. SsAGM1, the main gene that regulates uridine diphosphate-N-acetylglucosamine synthesis, was down-regulated in ΔSsStuA. Additionally, the chitin distributed randomly in the hyphae of ΔSsStuA, suggesting that SsStuA is involved in the CWI pathway. CYP51 has been reported as the target of tebuconazole, however, SsCYP51 was up-regulated in ΔSsStuA which suggests that there is a new target of tebuconazole in S. sclerotiorum, and SsStuA, a fungus-specific TF, can be developed as a new drug target for disease control.
In summary, we identified the APSES transcription factor SsStuA, as a key TF, affecting cell wall integrity, tolerance to fungicide and ROS metabolism, thereby regulating mycelial growth, sclerotium production and full virulence of S. sclerotiorum.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof10040238/s1, Figure S1. Identification of mutants by PCR and qPCR. (A) The knockout strategy of SsStuA gene. (B) Knockout of SsStuA was verified by PCR. (C) The knockout and complementation mutants of SsStuA were verified by PCR. Error bars represent the SDs and significant difference was performed by t test (***, p < 0.001). Figure S2. ΔSsStuA produced oxalic acid normally. Photographs were taken at 2 days post-inoculation. Table S1. Primers used in this paper.
Author Contributions
Data curation, M.L. and W.J.; Funding acquisition, H.P.; Methodology, W.J., M.L., X.L., J.H., X.Z., S.S. and J.L.; Project administration, H.P. and Y.Z.; Software, W.J., T.L. and J.Z.; Writing—original draft, W.J. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Key Research and Development Program of China (2023YFD1401000), Jilin Province Science and Technology Development Plan Program (20240303020NC), and Graduate Innovation Fund of Jilin University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
Thanks Airong Wang (Fujian Agriculture and Forestry University) for providing the vector “pNAH-ONG”.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Baniasadi, F.; Bonjar, G.H.S.; Baghizadeh, A.; Nik, A.K.; Jorjandi, M.; Aghighi, S.; Farokhi, P.R. Biological Control of Sclerotinia sclerotiorum, Causal Agent of Sunflower Head and Stem Rot Disease, by Use of Soil borne Actinomycetes Isolates. Am. J. Agric. Biol. Sci. 2009, 4, 146–151. [Google Scholar] [CrossRef]
- Willbur, J.; McCaghey, M.; Kabbage, M.; Smith, D.L. An overview of the Sclerotinia sclerotiorum pathosystem in soybean: Impact, fungal biology, and current management strategies. Trop. Plant Pathol. 2018, 44, 3–11. [Google Scholar] [CrossRef]
- Ni, Y.; Guo, Y.J.; Wang, J.; Xia, R.E.; Wang, X.Q.; Ash, G.; Li, J.N. Responses of physiological indexes and leaf epicuticular waxes of Brassica napus to Sclerotinia sclerotiorum infection. Plant Pathol. 2013, 63, 174–184. [Google Scholar] [CrossRef]
- Xu, L.; Li, G.; Jiang, D.; Chen, W. Sclerotinia sclerotiorum: An Evaluation of Virulence Theories. Annu. Rev. Phytopathol. 2018, 56, 311–338. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Sultana, F.; Li, W.; Tran, L.-S.P.; Mostofa, M.G. Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the Pathogenomic Features of a Global Pathogen. Cells 2023, 12, 1063. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Jiao, W.; Yu, H.; Chen, X.; Xiao, K.; Jia, D.; Wang, F.; Zhang, Y.; Pan, H. The SsAtg1 Activating Autophagy Is Required for Sclerotia Formation and Pathogenicity in Sclerotinia sclerotiorum. J. Fungi 2022, 8, 1314. [Google Scholar] [CrossRef]
- Erental, A.; Dickman, M.B.; Yarden, O. Sclerotial development in Sclerotinia sclerotiorum: Awakening molecular analysis of a “Dormant” structure. Fungal Bio Rev. 2008, 22, 6–16. [Google Scholar] [CrossRef]
- Jiang, D.; Fu, Y.; Guoqing, L.; Ghabrial, S.A. Viruses of the Plant Pathogenic Fungus Sclerotinia sclerotiorum. Adv. Virus Res. 2013, 86, 215–248. [Google Scholar] [CrossRef]
- Xia, S.; Xu, Y.; Hoy, R.; Zhang, J.; Qin, L.; Li, X. The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis. Pathogens 2019, 9, 27. [Google Scholar] [CrossRef]
- Chen, C.; Dickman, M.B. cAMP blocks MAPK activation and sclerotial development via Rap-1 in a PKA-independent manner in Sclerotinia sclerotiorum. Mol. Microbiol. 2004, 55, 299–311. [Google Scholar] [CrossRef]
- Chen, C.; Harel, A.; Gorovoits, R.; Yarden, O.; Dickman, M.B. MAPK regulation of sclerotial development in Sclerotinia sclerotiorum is linked with pH and cAMP sensing. Mol. Plant Microbe Interact. 2004, 17, 404–413. [Google Scholar] [CrossRef]
- Xu, T.; Li, J.; Yu, B.; Liu, L.; Zhang, X.; Liu, J.; Pan, H.; Zhang, Y. Transcription Factor SsSte12 Was Involved in Mycelium Growth and Development in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 2476. [Google Scholar] [CrossRef]
- Shelest, E. Transcription factors in fungi. FEMS Microbiol. Lett. 2008, 286, 145–151. [Google Scholar] [CrossRef]
- Zhao, Y.; Su, H.; Zhou, J.; Feng, H.; Zhang, K.-Q.; Yang, J. The APSES family proteins in fungi: Characterizations, evolution and functions. Fungal Genet. Biol. 2015, 81, 271–280. [Google Scholar] [CrossRef]
- Dutton, J.R.; Johns, S.; Miller, B.L. StuAp is a sequence-specific transcription factor that regulates developmental complexity in Aspergillus nidulans. EMBO J. 1997, 16, 5710–5721. [Google Scholar] [CrossRef]
- Iyer, L.M.; Koonin, E.V.; Aravind, L. Extensive domain shuffling in transcription regulators of DNA viruses and implications for the origin of fungal APSES transcription factors. Genome Biol. 2002, 3, RESEARCH0012. [Google Scholar] [CrossRef]
- Soyer, J.L.; Hamiot, A.; Ollivier, B.; Balesdent, M.H.; Rouxel, T.; Fudal, I. The APSES transcription factor LmStuA is required for sporulation, pathogenic development and effector gene expression in Leptosphaeria maculans. Mol. Plant Pathol. 2015, 16, 1000–1005. [Google Scholar] [CrossRef]
- Tebarth, B.; Doedt, T.; Krishnamurthy, S.; Weide, M.; Monterola, F.; Dominguez, A.; Ernst, J.F. Adaptation of the Efg1p morphogenetic pathway in Candida albicans by negative autoregulation and PKA-dependent repression of the EFG1 gene. J. Mol. Biol. 2003, 329, 949–962. [Google Scholar] [CrossRef]
- Doedt, T.; Krishnamurthy, S.; Bockmühl, D.P.; Tebarth, B.; Stempel, C.; Russell, C.L.; Brown, A.J.; Ernst, J.F. APSES Proteins Regulate Morphogenesis and Metabolism in Candida albicans. Mol. Biol. Cell 2004, 15, 3167–3180. [Google Scholar] [CrossRef]
- Shenhar, G.; Kassir, Y. A positive regulator of mitosis, Sok2, functions as a negative regulator of meiosis in Saccharomyces cerevisiae. Mol. Cell Biol. 2001, 21, 1603–1612. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Doedt, T.; Chiang, L.Y.; Kim, H.S.; Chen, D.; Nierman, W.C.; Filler, S.G. The Aspergillus fumigatus StuA protein governs the up-regulation of a discrete transcriptional program during the acquisition of developmental competence. Mol. Biol. Cell 2005, 16, 5866–5879. [Google Scholar] [CrossRef]
- Jun, S.C.; Choi, Y.H.; Lee, M.W.; Yu, J.H.; Shin, K.S. The Putative APSES Transcription Factor RgdA Governs Growth, Development, Toxigenesis, and Virulence in Aspergillus fumigatus. mSphere 2020, 5, e00998-20. [Google Scholar] [CrossRef]
- Tong, X.; Zhang, X.; Plummer, K.M.; Stowell, K.M.; Sullivan, P.A.; Farley, P.C. GcSTUA, an APSES transcription factor, is required for generation of appressorial turgor pressure and full pathogenicity of Glomerella cingulata. Mol. Plant Microbe Interact. 2007, 20, 1102–1111. [Google Scholar] [CrossRef]
- Nishimura, M.; Fukada, J.; Moriwaki, A.; Fujikawa, T.; Ohashi, M.; Hibi, T.; Hayashi, N. Mstu1, an APSES transcription factor, is required for appressorium-mediated infection in Magnaporthe grisea. Biosci. Biotechnol. Biochem. 2009, 73, 1779–1786. [Google Scholar] [CrossRef]
- Lysøe, E.; Pasquali, M.; Breakspear, A.; Kistler, H.C. The transcription factor FgStuAp influences spore development, pathogenicity, and secondary metabolism in Fusarium graminearum. Mol. Plant Microbe Interact. 2011, 24, 54–67. [Google Scholar] [CrossRef]
- Qi, Z.; Wang, Q.; Dou, X.; Wang, W.; Zhao, Q.; Lv, R.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. MoSwi6, an APSES family transcription factor, interacts with MoMps1 and is required for hyphal and conidial morphogenesis, appressorial function and pathogenicity of Magnaporthe oryzae. Mol. Plant Pathol. 2012, 13, 677–689. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Zhang, Y.; Yu, P.L.; Pan, H.; Rollins, J.A. Introduction of Large Sequence Inserts by CRISPR-Cas9 To Create Pathogenicity Mutants in the Multinucleate Filamentous Pathogen Sclerotinia sclerotiorum. mBio 2018, 9, e00567-18. [Google Scholar] [CrossRef]
- Jiao, W.; Yu, H.; Cong, J.; Xiao, K.; Zhang, X.; Liu, J.; Zhang, Y.; Pan, H. Transcription factor SsFoxE3 activating SsAtg8 is critical for sclerotia, compound appressoria formation, and pathogenicity in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2022, 23, 204–217. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Jurick, W.M.; Rollins, J.A. Deletion of the adenylate cyclase (sac1) gene affects multiple developmental pathways and pathogenicity in Sclerotinia sclerotiorum. Fungal Genet. Biol. 2007, 44, 521–530. [Google Scholar] [CrossRef]
- Goswami, R.S. Targeted gene replacement in fungi using a split-marker approach. Methods Mol. Biol. 2012, 835, 255–269. [Google Scholar] [CrossRef]
- Rollins, J.A. The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant Microbe Interact. 2003, 16, 785–795. [Google Scholar] [CrossRef]
- Sanchez-Adria, I.E.; Sanmartin, G.; Prieto, J.A.; Estruch, F.; Randez-Gil, F. Slt2 Is Required to Activate ER-Stress-Protective Mechanisms through TORC1 Inhibition and Hexosamine Pathway Activation. J Fungi 2022, 8, 92. [Google Scholar] [CrossRef]
- Yin, Z.; Feng, W.; Chen, C.; Xu, J.; Li, Y.; Yang, L.; Wang, J.; Liu, X.; Wang, W.; Gao, C.; et al. Shedding light on autophagy coordinating with cell wall integrity signaling to govern pathogenicity of Magnaporthe oryzae. Autophagy 2020, 16, 900–916. [Google Scholar] [CrossRef]
- Kang, Z.; Huang, L.; Krieg, U.; Mauler-Machnik, A.; Buchenauer, H. Effects of tebuconazole on morphology, structure, cell wall components and trichothecene production of Fusarium culmorum in vitro. Pest. Manag. Sci. 2001, 57, 491–500. [Google Scholar] [CrossRef]
- Fan, K.; Fu, L.; Liu, H.; Qu, J.; Zhang, G.; Zhang, S.; Qiao, K. Reduced Sensitivity to Tebuconazole in Botryosphaeria dothidea Isolates Collected from Major Apple Production Areas of China. Plant Dis. 2022, 106, 2817–2822. [Google Scholar] [CrossRef]
- Moore, L.C.; Brenneman, T.B.; Waliullah, S.; Bock, C.H.; Ali, M.E. Multiple Mutations and Overexpression in the CYP51A and B Genes Lead to Decreased Sensitivity of Venturia effusa to Tebuconazole. Curr. Issues Mol. Biol. 2022, 44, 670–685. [Google Scholar] [CrossRef]
- Wei, L.-l.; Chen, W.-c.; Zhao, W.-c.; Wang, J.; Wang, B.-r.; Li, F.-j.; Wei, M.-d.; Guo, J.; Chen, C.-j.; Zheng, J.-q.; et al. Mutations and Overexpression of CYP51 Associated with DMI-Resistance in Colletotrichum gloeosporioides from Chili. Plant Dis. 2020, 104, 668–676. [Google Scholar] [CrossRef]
- Ben Othmène, Y.; Monceaux, K.; Belhadef, A.; Karoui, A.; Ben Salem, I.; Boussabbeh, M.; Abid-Essefi, S.; Lemaire, C. Triazole fungicide tebuconazole induces apoptosis through ROS-mediated endoplasmic reticulum stress pathway. Env. Toxicol. Pharmacol. 2022, 94, 103919. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef]
- Sarmiento-Villamil, J.L.; García-Pedrajas, N.E.; Baeza-Montañez, L.; García-Pedrajas, M.D. The APSES transcription factor Vst1 is a key regulator of development in microsclerotium- and resting mycelium-producing Verticillium species. Mol. Plant Pathol. 2018, 19, 59–76. [Google Scholar] [CrossRef]
- Miller, K.Y.; Wu, J.; Miller, B.L. StuA is required for cell pattern formation in Aspergillus. Genes. Dev. 1992, 6, 1770–1782. [Google Scholar] [CrossRef]
- Clutterbuck, A.J. A mutational analysis of conidial development in Aspergillus nidulans. Genetics 1969, 63, 317–327. [Google Scholar] [CrossRef]
- Yang, W.; Wu, H.; Wang, Z.; Sun, Q.; Qiao, L.; Huang, B. The APSES Gene MrStuA Regulates Sporulation in Metarhizium robertsii. Front. Microbiol. 2018, 9, 1208. [Google Scholar] [CrossRef]
- Liang, X.; Rollins, J.A. Mechanisms of Broad Host Range Necrotrophic Pathogenesis in Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1128–1140. [Google Scholar] [CrossRef]
- Kim, K.S.; Min, J.Y.; Dickman, M.B. Oxalic acid is an elicitor of plant programmed cell death during Sclerotinia sclerotiorum disease development. Mol. Plant Microbe Interact. 2008, 21, 605–612. [Google Scholar] [CrossRef]
- Govrin, E.M.; Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol. 2000, 10, 751–757. [Google Scholar] [CrossRef]
- Rana, K.; Ding, Y.; Banga, S.S.; Liao, H.; Zhao, S.; Yu, Y.; Qian, W. Sclerotinia sclerotiorum Thioredoxin1 (SsTrx1) is required for pathogenicity and oxidative stress tolerance. Mol. Plant Pathol. 2021, 22, 1413–1426. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).