
Hidden Treasure: Halophilic Fungi as a Repository of Bioactive Lead Compounds




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Physiology of Halophilicity in Fungi
3. Ecology and Biodiversity of Halophilic Fungi
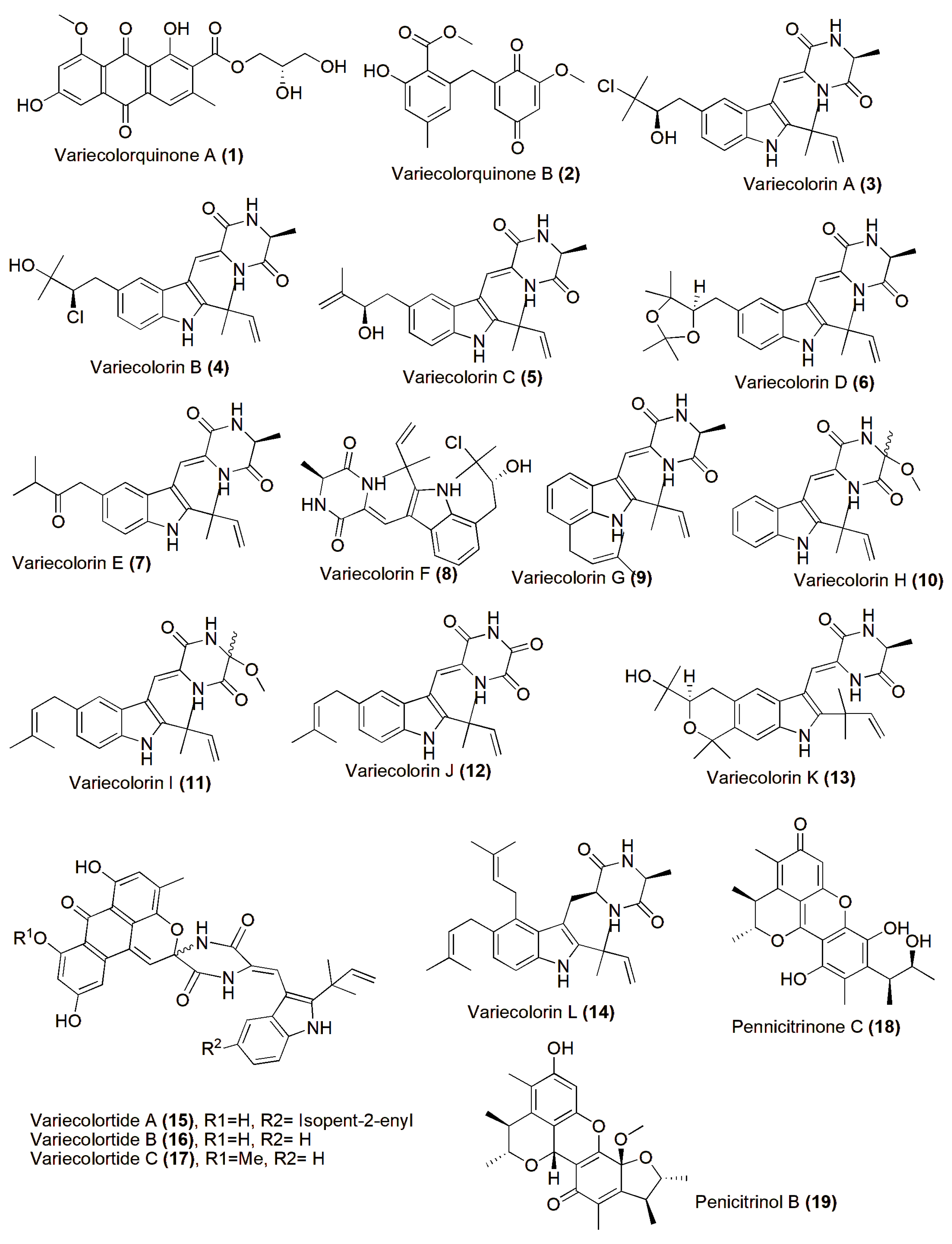
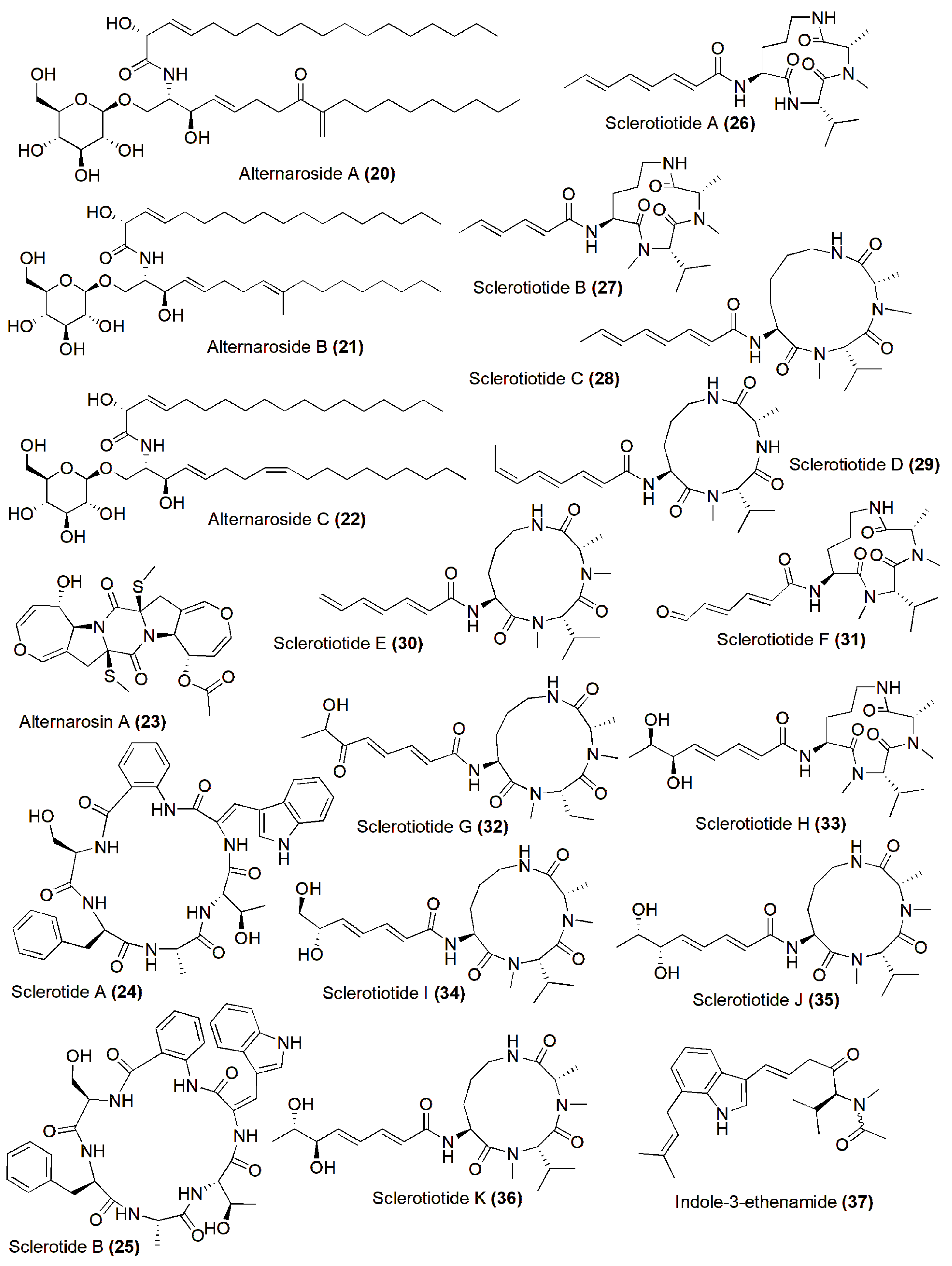
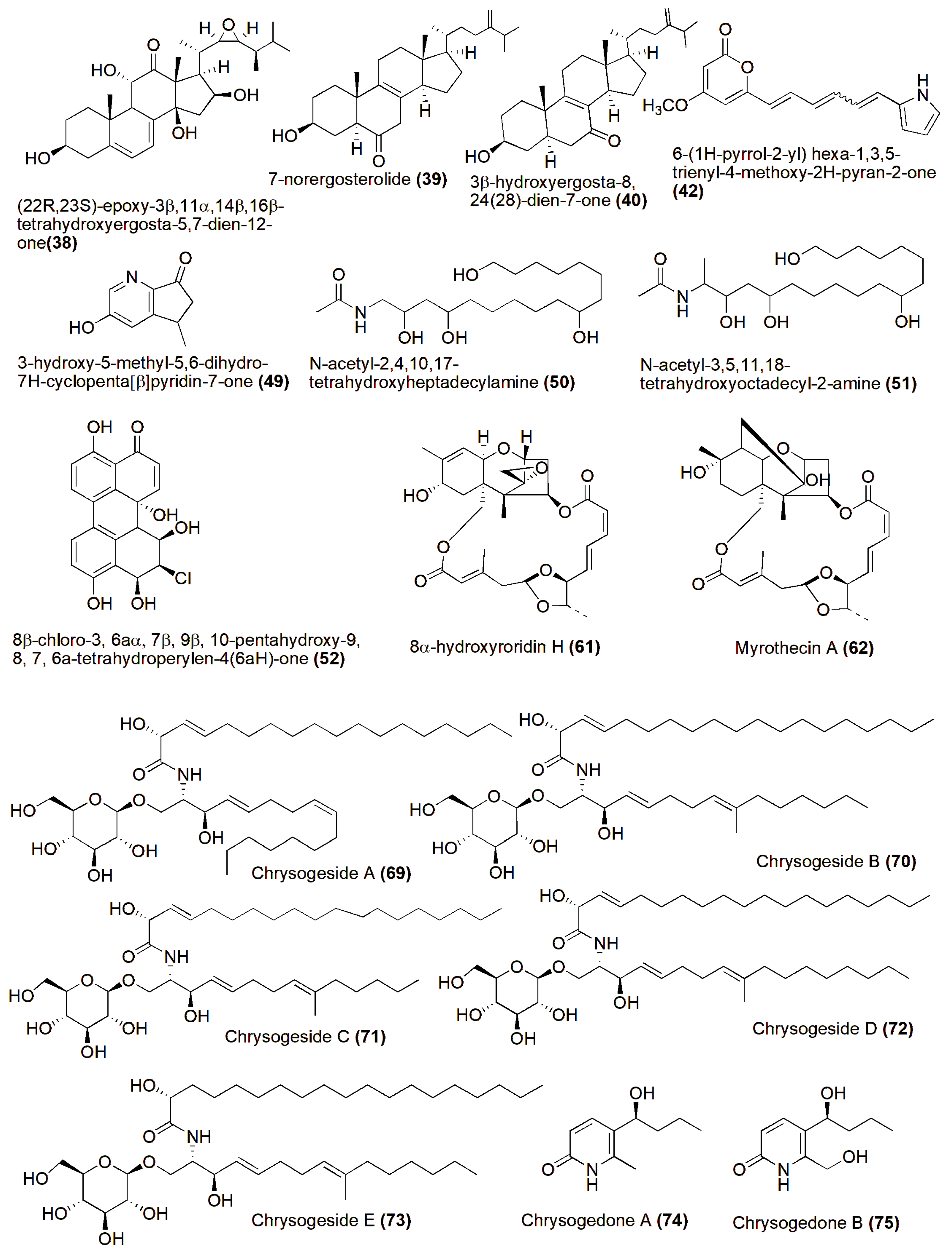
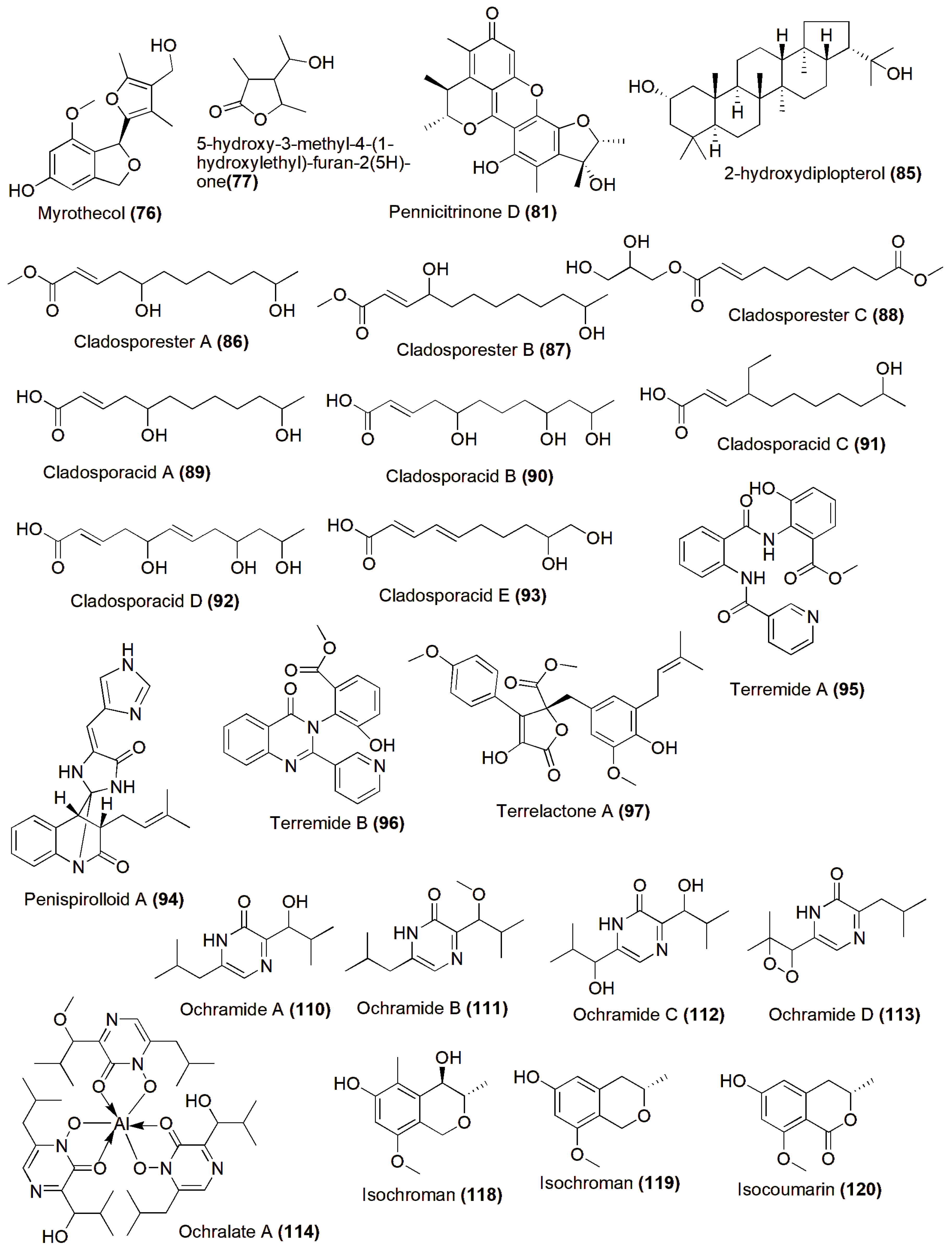
4. Role in Production of Therapeutic Compounds
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chavan, P.; Agrawal, S. Uncovering the Desert Fungal Enigma: An Attractive Resource for Biopharmaceuticals. In Fungi Bioactive Metabolites: Integration of Pharmaceutical Applications; Springer: Berlin/Heidelberg, Germany, 2024; pp. 517–536. [Google Scholar]
- Rodriguez-Valera, F. Introduction to saline environments. In The Biology of Halophilic Bacteria; CRC Press: Boca Raton, FL, USA, 2020; pp. 1–23. [Google Scholar]
- Buchalo, A.S.; Nevo, E.; Wasser, S.P.; Oren, A.; Molitoris, H.P. Fungal life in the extremely hypersaline water of the Dead Sea: First records. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 1461–1465. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Zalar, P.; de Hoog, S.; Plemenitaš, A. Hypersaline waters in salterns–natural ecological niches for halophilic black yeasts. FEMS Microbiol. Ecol. 2000, 32, 235–240. [Google Scholar]
- Butinar, L.; Santos, S.; Spencer-Martins, I.; Oren, A.; Gunde-Cimerman, N. Yeast diversity in hypersaline habitats. FEMS Microbiol. Lett. 2005, 244, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Rampelotto, P.H. Extremophiles and extreme environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Nonribosomal peptides from marine microbes and their antimicrobial and anticancer potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Marine fungi: An untapped bioresource for future cosmeceuticals. Phytochem. Lett. 2018, 23, 15–20. [Google Scholar] [CrossRef]
- Agrawal, S.; Deshmukh, S.K.; Reddy, M.S.; Prasad, R.; Goel, M. Endolichenic fungi: A hidden source of bioactive metabolites. S. Afr. J. Bot. 2020, 134, 163–186. [Google Scholar] [CrossRef]
- Agrawal, S.; Samanta, S.; Deshmukh, S.K. The antidiabetic potential of endophytic fungi: Future prospects as therapeutic agents. Biotechnol. Appl. Biochem. 2022, 69, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Agrawal, S.; Prakash, V.; Gupta, M.K.; Reddy, M.S. Anti-infectives from mangrove endophytic fungi. S. Afr. J. Bot. 2020, 134, 237–263. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Agrawal, S.; Gupta, M.K.; Patidar, R.K.; Ranjan, N. Recent advances in the discovery of antiviral metabolites from fungi. Curr. Pharm. Biotechnol. 2022, 23, 495–537. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Agrawal, S.; Gupta, M.K. Fungal Metabolites: A Potential Source of Antidiabetic Agents with Particular Reference to PTP1B Inhibitors. Curr. Pharm. Biotechnol. 2023, 24, 927–945. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Gupta, M.K.; Agrawal, S. Antidiabetic Agents from Fungi with Special Reference to α-Glucosidase Inhibitors. Recent Pat. Biotechnol. 2023, 17, 24–61. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.-Y.; Chen, S.-A.; Hsueh, Y.-P. The high osmolarity glycerol (HOG) pathway functions in osmosensing, trap morphogenesis and conidiation of the nematode-trapping fungus Arthrobotrys oligospora. J. Fungi 2020, 6, 191. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Gunde-Cimerman, N. Understanding Fungi in Glacial and Hypersaline Environments. Annu. Rev. Microbiol. 2023, 77, 89–109. [Google Scholar] [CrossRef] [PubMed]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to high salt concentrations in halotolerant/halophilic fungi: A molecular perspective. Front. Microbiol. 2014, 5, 199. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.; Cannon, P.; David, J.; Stalpers, J. Ainsworth and Bisbys Dictionary of the Fun. In Ainsworth and Bisbys Dictionary of the Fun; CABI: Wallingford, UK, 2001. [Google Scholar]
- Raol, G.G.; Raol, B.; Prajapati, V.S.; Bhavsar, N.H. Utilization of agro-industrial waste for β-galactosidase production under solid state fermentation using halotolerant Aspergillus tubingensis GR1 isolate. 3 Biotech 2015, 5, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Ruginescu, R.; Gomoiu, I.; Popescu, O.; Cojoc, R.; Neagu, S.; Lucaci, I.; Batrinescu-Moteau, C.; Enache, M. Bioprospecting for novel halophilic and halotolerant sources of hydrolytic enzymes in brackish, saline and hypersaline lakes of Romania. Microorganisms 2020, 8, 1903. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, L.K.; Jitprasitporn, N.; Che-Alee, N. Isolation and characterization of halophilic and halotolerant fungi from man-made solar salterns in Pattani Province, Thailand. PLoS ONE 2023, 18, e0281623. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.S.; Gonsalves, V.; Nazareth, S.W. Isolation and salt tolerance of halophilic fungi from mangroves and solar salterns in Goa, India. Indian J. Geo-Mar. Sci. 2012, 41, 164–172. [Google Scholar]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.; Heitman, J.; Hom, E.F.; Ianiri, G.; Jones, A.C. Fungi in the marine environment: Open questions and unsolved problems. MBio 2019, 10, 10-1128. [Google Scholar] [CrossRef]
- Chung, D.; Kim, H.; Choi, H.S. Fungi in salterns. J. Microbiol. 2019, 57, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Anastasiou, C. Fungi from salt lakes. I. A new species of Clavariopsis. Mycologia 1961, 53, 11–16. [Google Scholar] [CrossRef]
- Oren, A.; Gunde-Cimerman, N. Fungal life in the Dead Sea. In Biology of Marine Fungi; Springer: Berlin/Heidelberg, Germany, 2011; pp. 115–132. [Google Scholar]
- Chamekh, R.; Deniel, F.; Donot, C.; Jany, J.-L.; Nodet, P.; Belabid, L. Isolation, identification and enzymatic activity of halotolerant and halophilic fungi from the Great Sebkha of Oran in Northwestern of Algeria. Mycobiology 2019, 47, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Lenassi, M.; Gunde-Cimerman, N.; Plemenitaš, A. Fungal adaptation to extremely high salt concentrations. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 77, pp. 71–96. [Google Scholar]
- Cantrell, S.A.; Casillas-Martínez, L.; Molina, M. Characterization of fungi from hypersaline environments of solar salterns using morphological and molecular techniques. Mycol. Res. 2006, 110, 962–970. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Zalar, P.; Petrovič, U.; Turk, M.; Kogej, T.; de Hoog, G.S.; Plemenitaš, A. Fungi in salterns. In Halophilic Microorganisms; Springer: Berlin/Heidelberg, Germany, 2004; pp. 103–113. [Google Scholar]
- Gostinčar, C.; Turk, M.; Plemenitaš, A.; Gunde-Cimerman, N. The expressions of Δ9-, Δ12-desaturases and an elongase by the extremely halotolerant black yeast Hortaea werneckii are salt dependent. FEMS Yeast Res. 2009, 9, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Zalar, P.; Sybren de Hoog, G.; Schroers, H.-J.; Frank, J.M.; Gunde-Cimerman, N. Taxonomy and phylogeny of the xerophilic genus Wallemia (Wallemiomycetes and Wallemiales, cl. et ord. nov.). Antonie Van Leeuwenhoek 2005, 87, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Kogej, T.; Galinski, E.A.; Ramos, J.; Gunde-Cimerman, N. Osmoadaptation strategy of the most halophilic fungus, Wallemia ichthyophaga, growing optimally at salinities above 15% NaCl. Appl. Environ. Microbiol. 2014, 80, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Life in hypersaline environments. In Their World: A Diversity of Microbial Environments; Springer: Berlin/Heidelberg, Germany, 2016; pp. 301–339. [Google Scholar] [CrossRef]
- Gocheva, Y.; Angelova, M.; Krumova, E. Potential of Halotolerant and Halophilic Fungi as a Source of New Extracellular Enzymes and Antimicrobial Compounds. Acta Microbiol. Bulg 2021, 37, 57–68. [Google Scholar]
- Wang, W.; Zhu, T.; Tao, H.; Lu, Z.; Fang, Y.; Gu, Q.; Zhu, W. Two new cytotoxic quinone type compounds from the halotolerant fungus Aspergillus variecolor. J. Antibiot. 2007, 60, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-L.; Lu, Z.-Y.; Tao, H.-W.; Zhu, T.-J.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Isoechinulin-type alkaloids, variecolorins A–L, from halotolerant Aspergillus variecolor. J. Nat. Prod. 2007, 70, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.L.; Zhu, T.J.; Tao, H.W.; Lu, Z.Y.; Fang, Y.C.; Gu, Q.Q.; Zhu, W.M. Three novel, structurally unique spirocyclic alkaloids from the halotolerant B-17 fungal strain of Aspergillus variecolor. Chem. Biodivers. 2007, 4, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-Y.; Lin, Z.-J.; Wang, W.-L.; Du, L.; Zhu, T.-J.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Citrinin dimers from the halotolerant fungus Penicillium citrinum B-57. J. Nat. Prod. 2008, 71, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Y.; Tao, H.; Peng, X.; Liu, P.; Zhu, W. Cerebrosides of the halotolerant fungus Alternaria raphani isolated from a sea salt field. J. Nat. Prod. 2009, 72, 1695–1698. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhu, H.; Hong, K.; Wang, Y.; Liu, P.; Wang, X.; Peng, X.; Zhu, W. Novel cyclic hexapeptides from marine-derived fungus, Aspergillus sclerotiorum PT06-1. Org. Lett. 2009, 11, 5262–5265. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xu, Z.; Wang, Y.; Hong, K.; Liu, P.; Zhu, W. Cyclic tripeptides from the halotolerant fungus Aspergillus sclerotiorum PT06-1. J. Nat. Prod. 2010, 73, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, J.-K.; Qu, H.-J.; Liu, P.-P.; Wang, Y.; Zhu, W.-M. A new cytotoxic indole-3-ethenamide from the halotolerant fungus Aspergillus sclerotiorum PT06-1. J. Antibiot. 2011, 64, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wang, Y.; Wang, J.; Liu, P.; Li, J.; Zhu, W. Antimicrobial ergosteroids and pyrrole derivatives from halotolerant Aspergillus flocculosus PT05-1 cultured in a hypersaline medium. Extremophiles 2013, 17, 963–971. [Google Scholar] [CrossRef]
- Peng, X.-P.; Wang, Y.; Liu, P.-P.; Hong, K.; Chen, H.; Yin, X.; Zhu, W.-M. Aromatic compounds from the halotolerant fungal strain of Wallemia sebi PXP-89 in a hypersaline medium. Arch. Pharmacal Res. 2011, 34, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, S.; Zhu, J.; Pan, H.; Bai, J.; Li, Z.; Guan, L.; Liu, G.; Yuan, C.; Wu, X. Two new amides from a halotolerant fungus, Myrothecium sp. GS-17. J. Antibiot. 2015, 68, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-Y.; Zhan-Lin, L.; Jiao, B.; Yu, W.; Zhang, L.-M.; Xin, W.; Hui-Ming, H. A new perylenequinone from a halotolerant fungus, Alternaria sp. M6. Chin. J. Nat. Med. 2012, 10, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Li, Z.L.; Guan, L.P.; Wu, X.; Pan, H.Q.; Bai, J.; Hua, H.M. Structure determination of two new trichothecenes from a halotolerant fungus Myrothecium sp. GS-17 by NMR spectroscopy. Magn. Reson. Chem. 2012, 50, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wang, Y.; Sun, K.; Liu, P.; Yin, X.; Zhu, W. Cerebrosides and 2-pyridone alkaloids from the halotolerant fungus Penicillium chrysogenum grown in a hypersaline medium. J. Nat. Prod. 2011, 74, 1298–1302. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, J.; Zhang, S.-Y.; Li, Z.-L.; Guan, L.-P.; Pan, H.-Q.; Wu, X.; Bai, J.; Hua, H.-M. Polyketides from the halotolerant fungus Myrothecium sp. GS-17. Molecules 2013, 18, 15126–15133. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.-H.; Wang, W.-L.; Zhang, Y.-P.; Xie, H.; Gu, Q.-Q.; Zhu, W.-M. Pennicitrinone D, a new citrinin dimer from the halotolerant fungus Penicillium notatum B-52. J. Antibiot. 2009, 62, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-L.; Liu, P.-P.; Zhang, Y.-P.; Li, J.; Tao, H.-W.; Gu, Q.-Q.; Zhu, W.-M. 2-Hydroxydiplopterol, a new cytotoxic pentacyclic triterpenoid from the halotolerant fungus Aspergillus variecolor B-17. Arch. Pharmacal Res. 2009, 32, 1211–1214. [Google Scholar] [CrossRef]
- Peng, X.; Wang, Y.; Zhu, G.; Zhu, W. Fatty acid derivatives from the halotolerant fungus Cladosporium cladosporioides. Magn. Reson. Chem. 2018, 56, 18–24. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Liu, Z.; Yang, J.; Fu, P.; Peng, J.; Zhu, W.-M.; Qi, S.-H. A novel antifouling alkaloid from halotolerant fungus Penicillium sp. OUCMDZ-776. Tetrahedron Lett. 2012, 53, 2280–2283. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, J.; Liu, P.; Wang, W.; Zhu, W. Three new compounds from Aspergillus terreus PT06-2 grown in a high salt medium. Mar. Drugs 2011, 9, 1368–1378. [Google Scholar] [CrossRef]
- Peng, X.; Wang, Y.; Zhu, T.; Zhu, W. Pyrazinone derivatives from the coral-derived Aspergillus ochraceus LCJ11-102 under high iodide salt. Arch. Pharmacal Res. 2018, 41, 184–191. [Google Scholar] [CrossRef]
- Orfali, R.S.; Aly, A.H.; Ebrahim, W.; Proksch, P. Isochroman and isocoumarin derivatives from hypersaline lake sediment-derived fungus Penicillium sp. Phytochem. Lett. 2015, 13, 234–238. [Google Scholar] [CrossRef]
- Shankar, A.; Sharma, K.K. Fungal secondary metabolites in food and pharmaceuticals in the era of multi-omics. Appl. Microbiol. Biotechnol. 2022, 106, 3465–3488. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, J.; Jiang, H.; Wang, G.; Wang, Y. Deep sequencing of the Sanghuangporus vaninii transcriptome reveals dynamic landscapes of candidate genes involved in the biosynthesis of active compounds. Arch. Microbiol. 2021, 203, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.; Hilário, S.; Van de Peer, Y.; Esteves, A.C.; Alves, A. Genomic and metabolomic analyses of the marine fungus Emericellopsis cladophorae: Insights into saltwater adaptability mechanisms and its biosynthetic potential. J. Fungi 2021, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Gómez, I.; Valdés-Muñoz, G.; Moreno-Ulloa, A.; Pérez-Llano, Y.; Moreno-Perlín, T.; Silva-Jiménez, H.; Sánchez-Carbente, M.d.R.; Folch-Mallol, J.L.; Gunde-Cimerman, N.; Lago-Lestón, A. Surviving in the brine: A multi-omics approach for understanding the physiology of the halophile fungus Aspergillus sydowii at saturated NaCl concentration. Front. Microbiol. 2022, 13, 840408. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrawal, S.; Chavan, P.; Dufossé, L. Hidden Treasure: Halophilic Fungi as a Repository of Bioactive Lead Compounds. J. Fungi 2024, 10, 290. https://doi.org/10.3390/jof10040290
Agrawal S, Chavan P, Dufossé L. Hidden Treasure: Halophilic Fungi as a Repository of Bioactive Lead Compounds. Journal of Fungi. 2024; 10(4):290. https://doi.org/10.3390/jof10040290
Chicago/Turabian StyleAgrawal, Shivankar, Pruthviraj Chavan, and Laurent Dufossé. 2024. "Hidden Treasure: Halophilic Fungi as a Repository of Bioactive Lead Compounds" Journal of Fungi 10, no. 4: 290. https://doi.org/10.3390/jof10040290
APA StyleAgrawal, S., Chavan, P., & Dufossé, L. (2024). Hidden Treasure: Halophilic Fungi as a Repository of Bioactive Lead Compounds. Journal of Fungi, 10(4), 290. https://doi.org/10.3390/jof10040290