
Aspergillus nidulans gfdB, Encoding the Hyperosmotic Stress Protein Glycerol-3-phosphate Dehydrogenase, Disrupts Osmoadaptation in Aspergillus wentii
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Reverse-Transcription Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Assays
2.3. High Throughput RNA Sequencing
2.4. Evaluation of the Transcriptome Data
3. Results
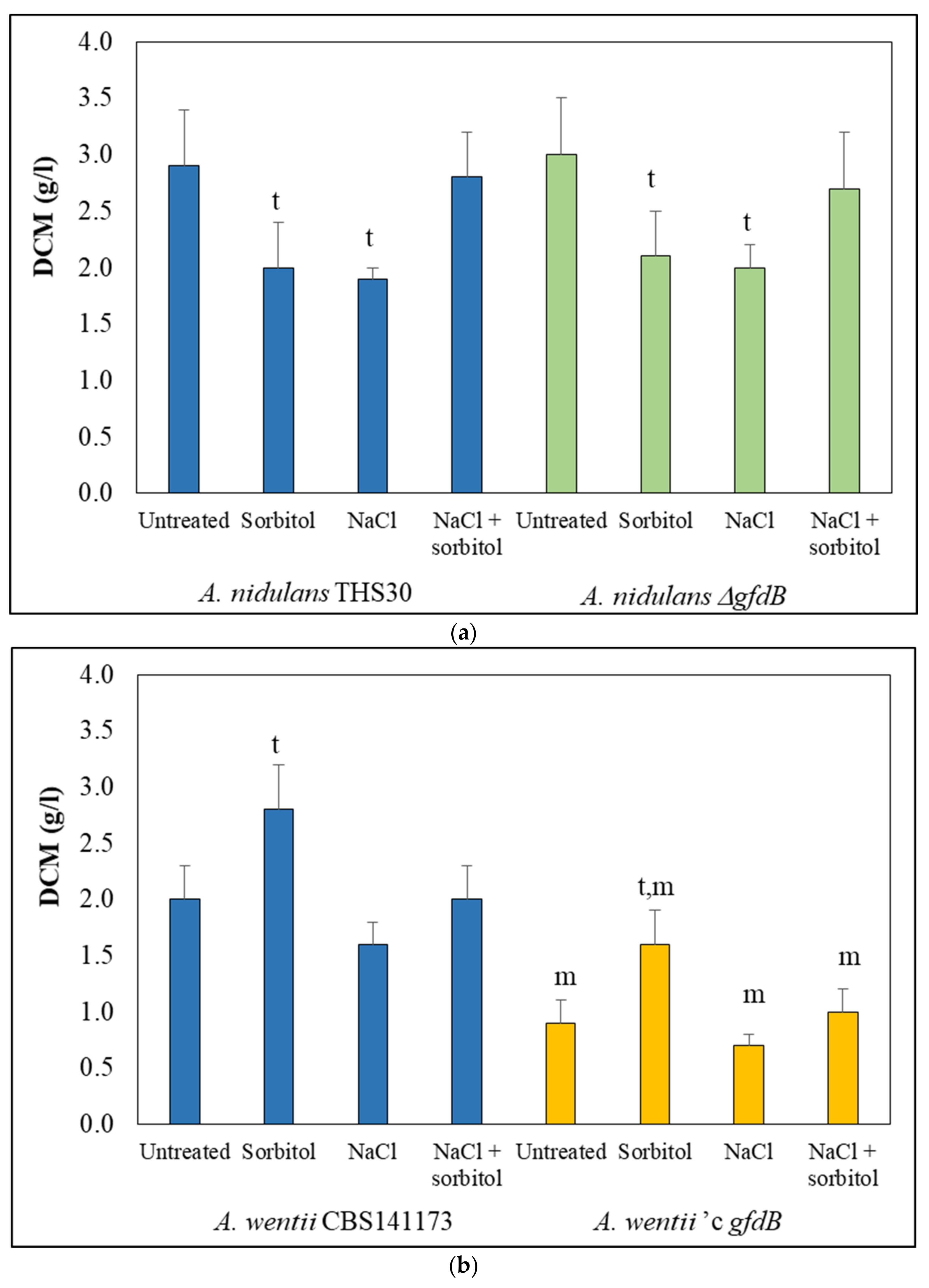
3.1. Deletion of gfdB in A. nidulans Produced Negligible Effect on Osmotolerance, While Expression of A. nidulans gfdB in A. wentii Reduced Growth, Irrespectively of the Osmolarity
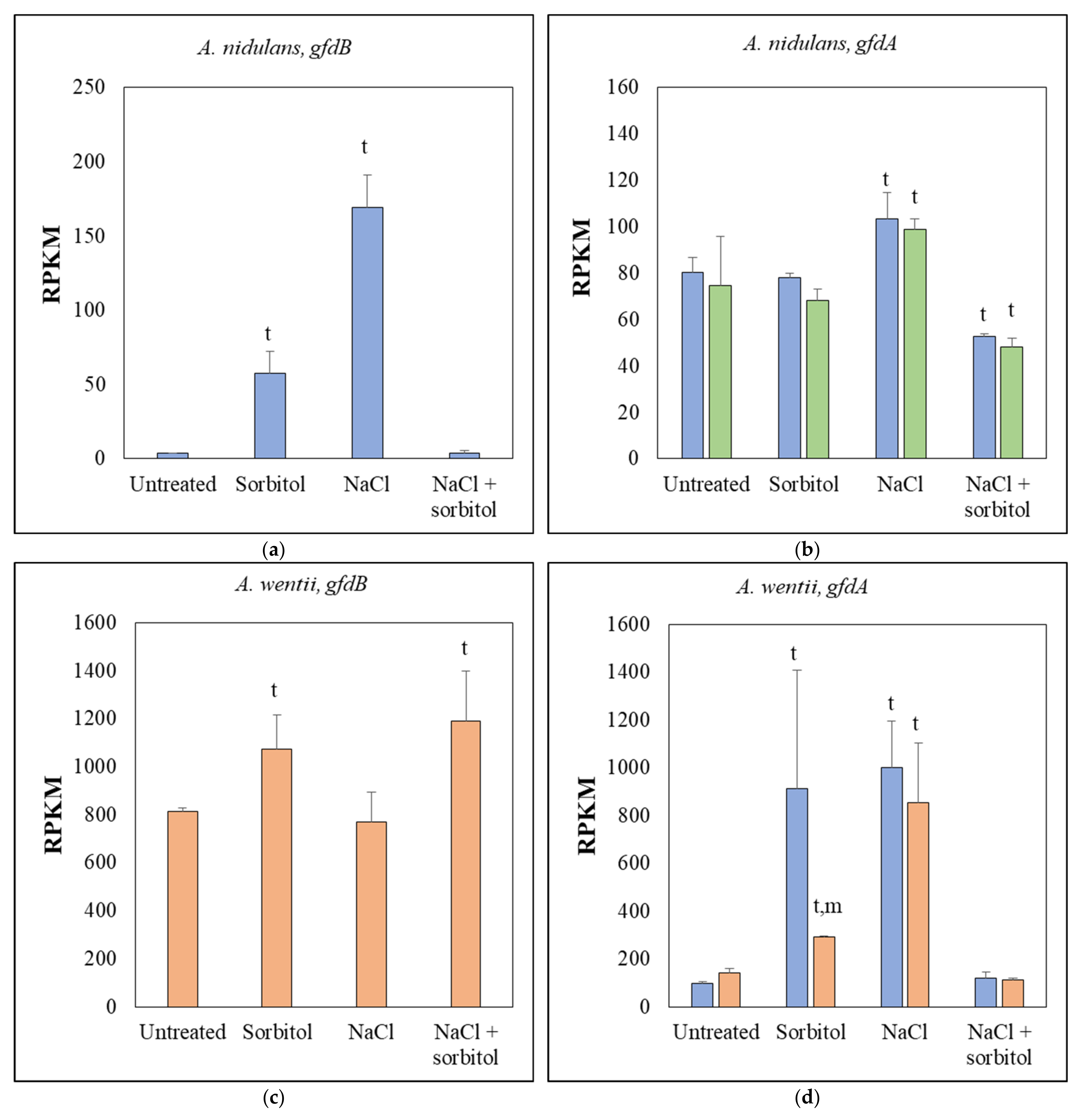
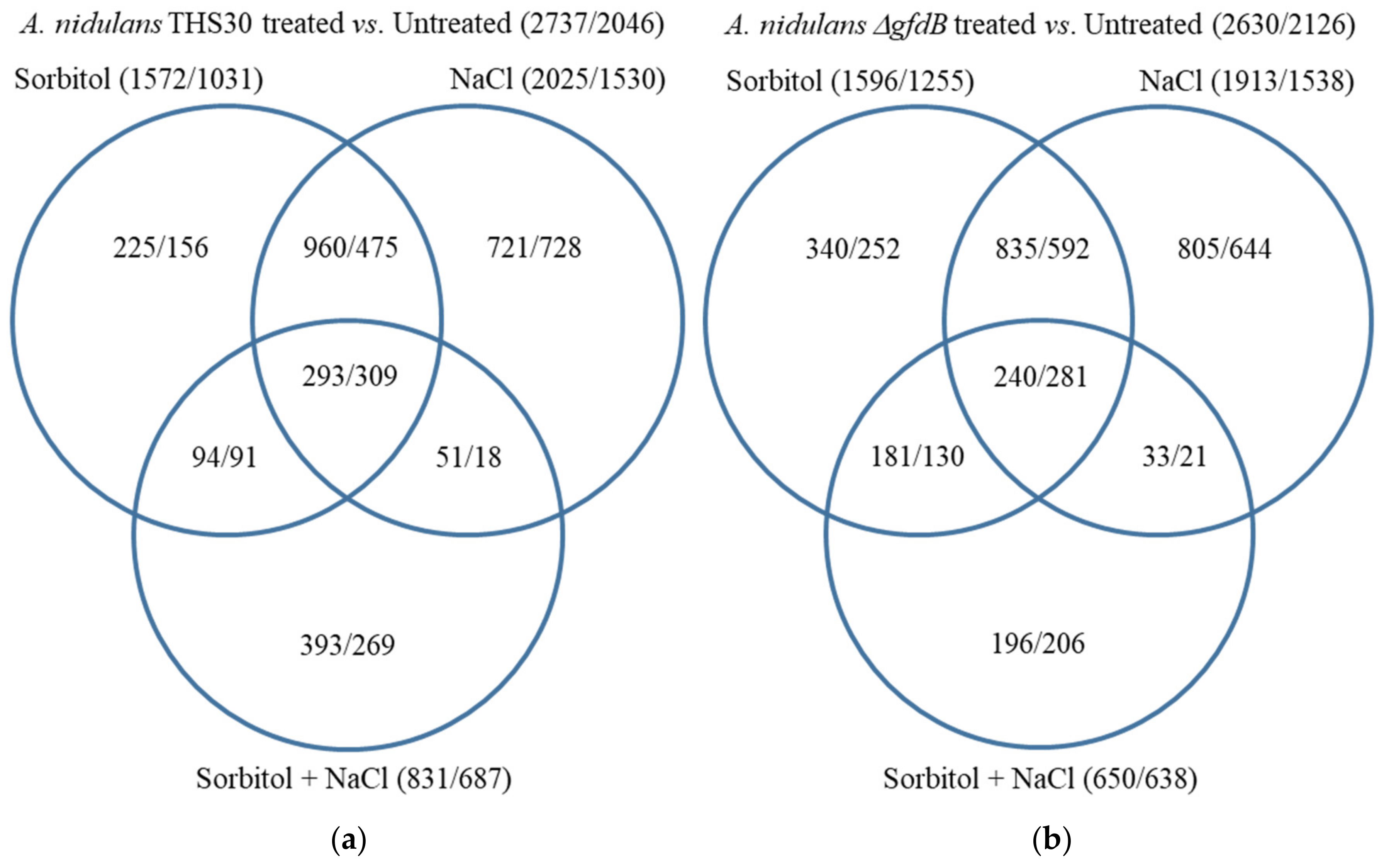
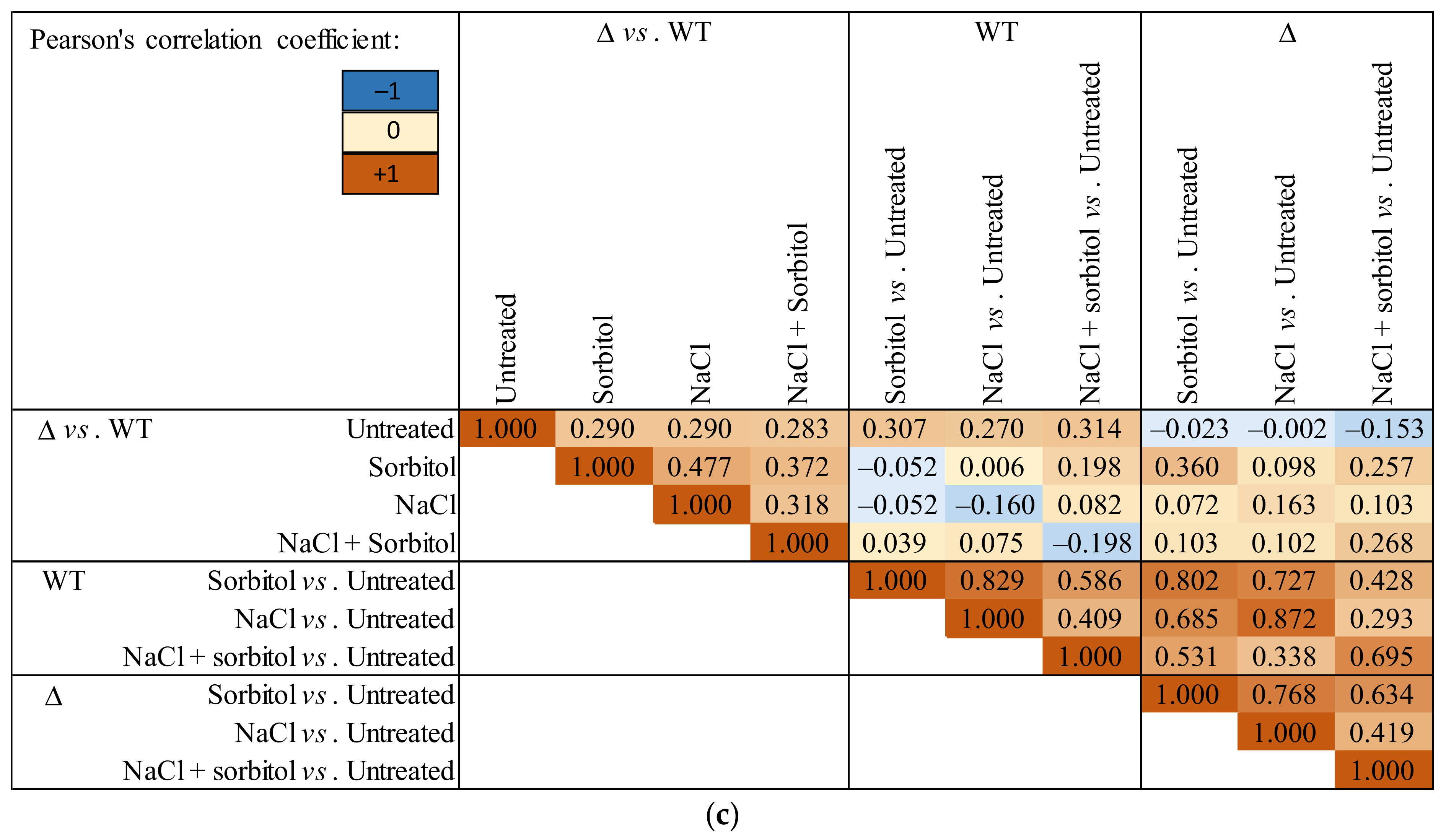
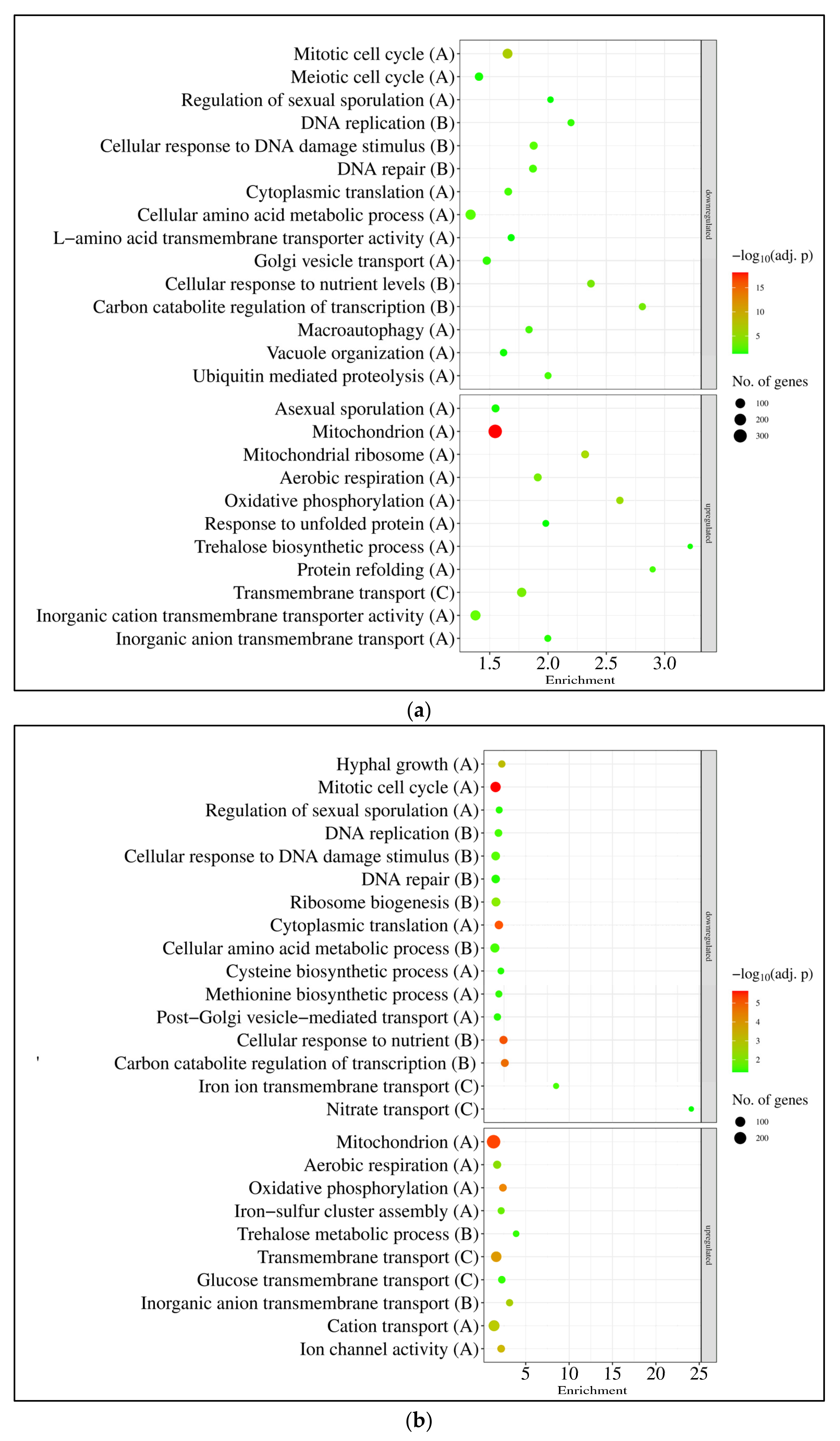
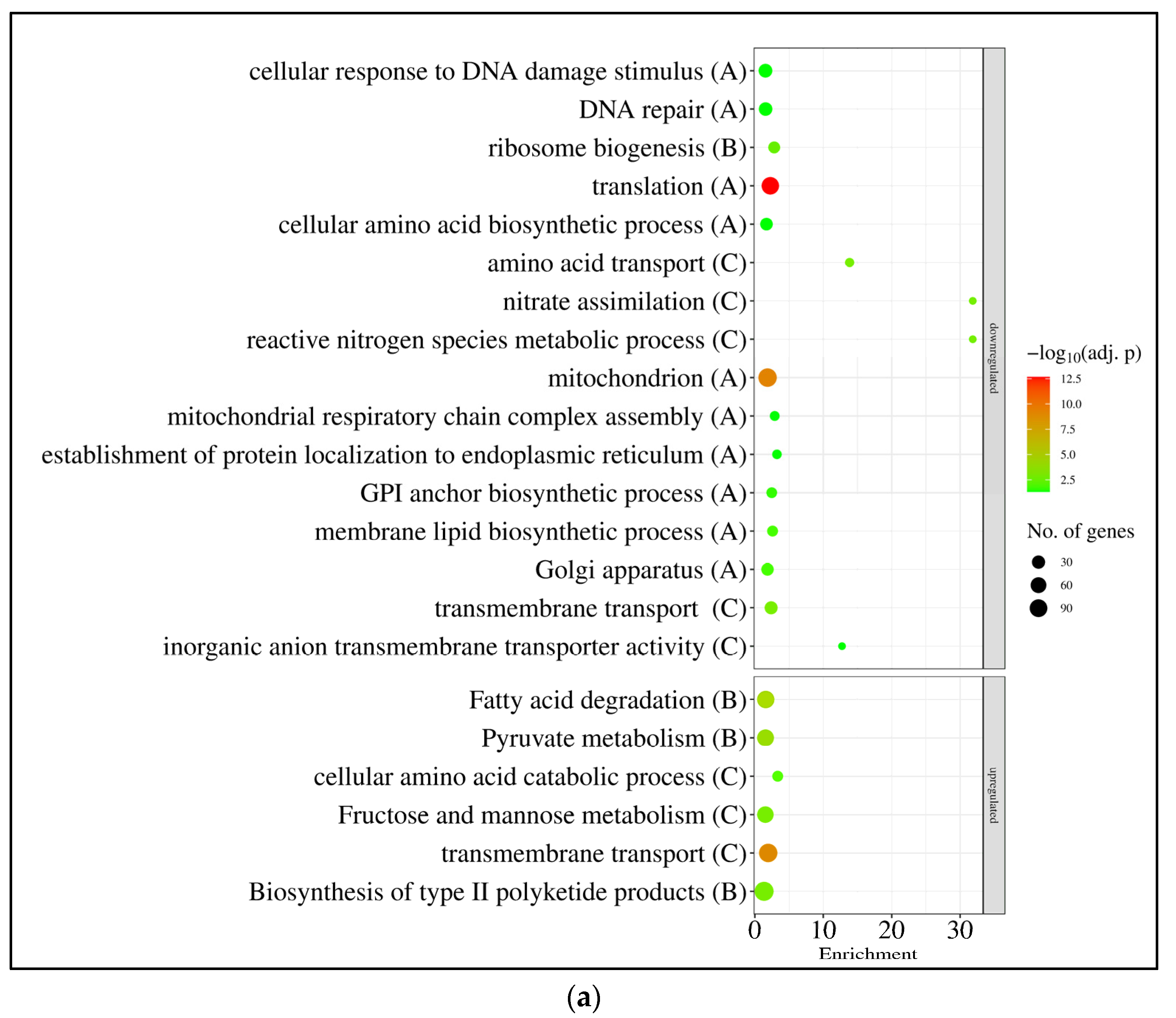
3.2. RNAseq Data Revealed That the Absence of GfdB Caused Only Minor Changes in the Osmotic Stress Response of A. nidulans
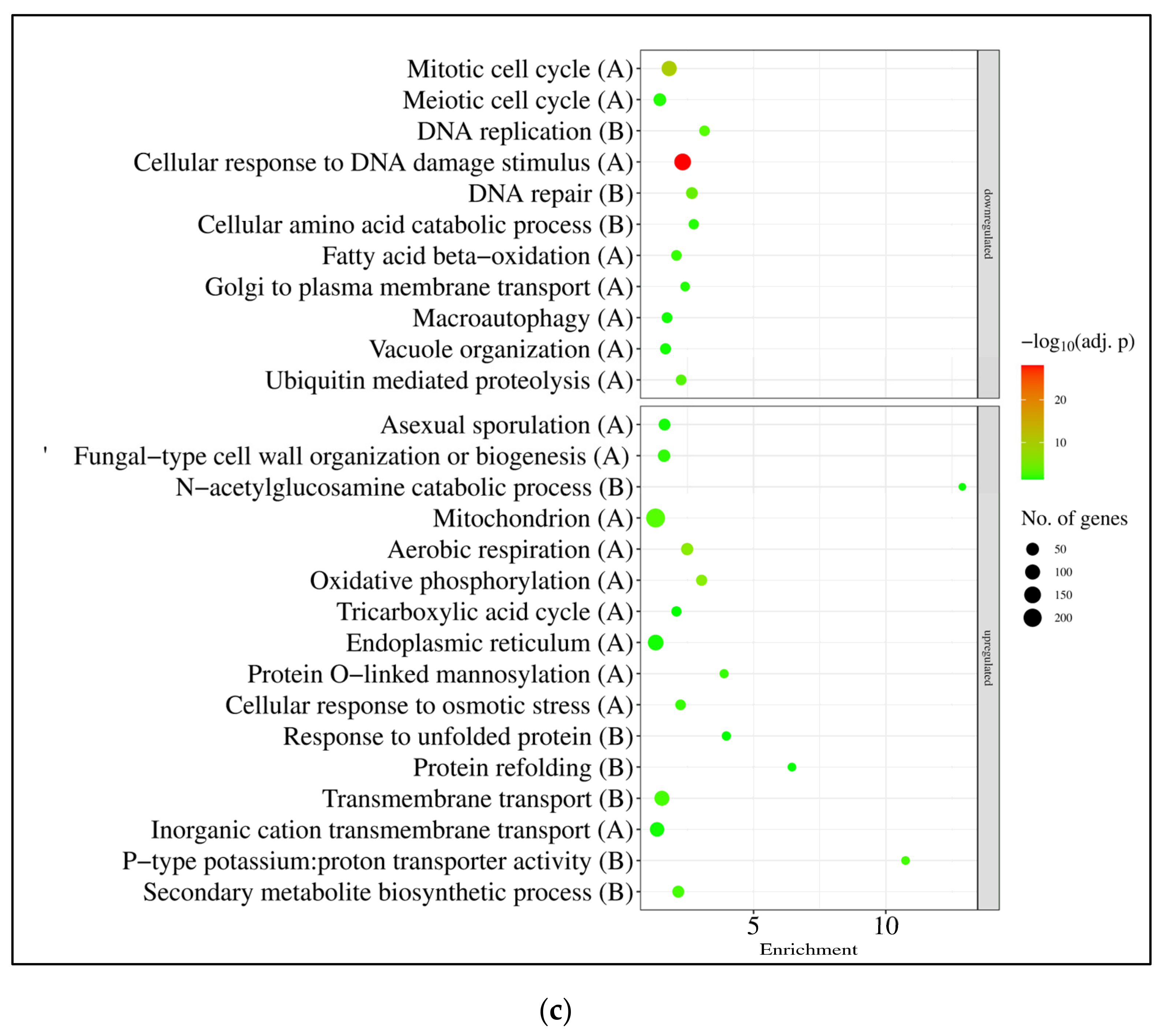
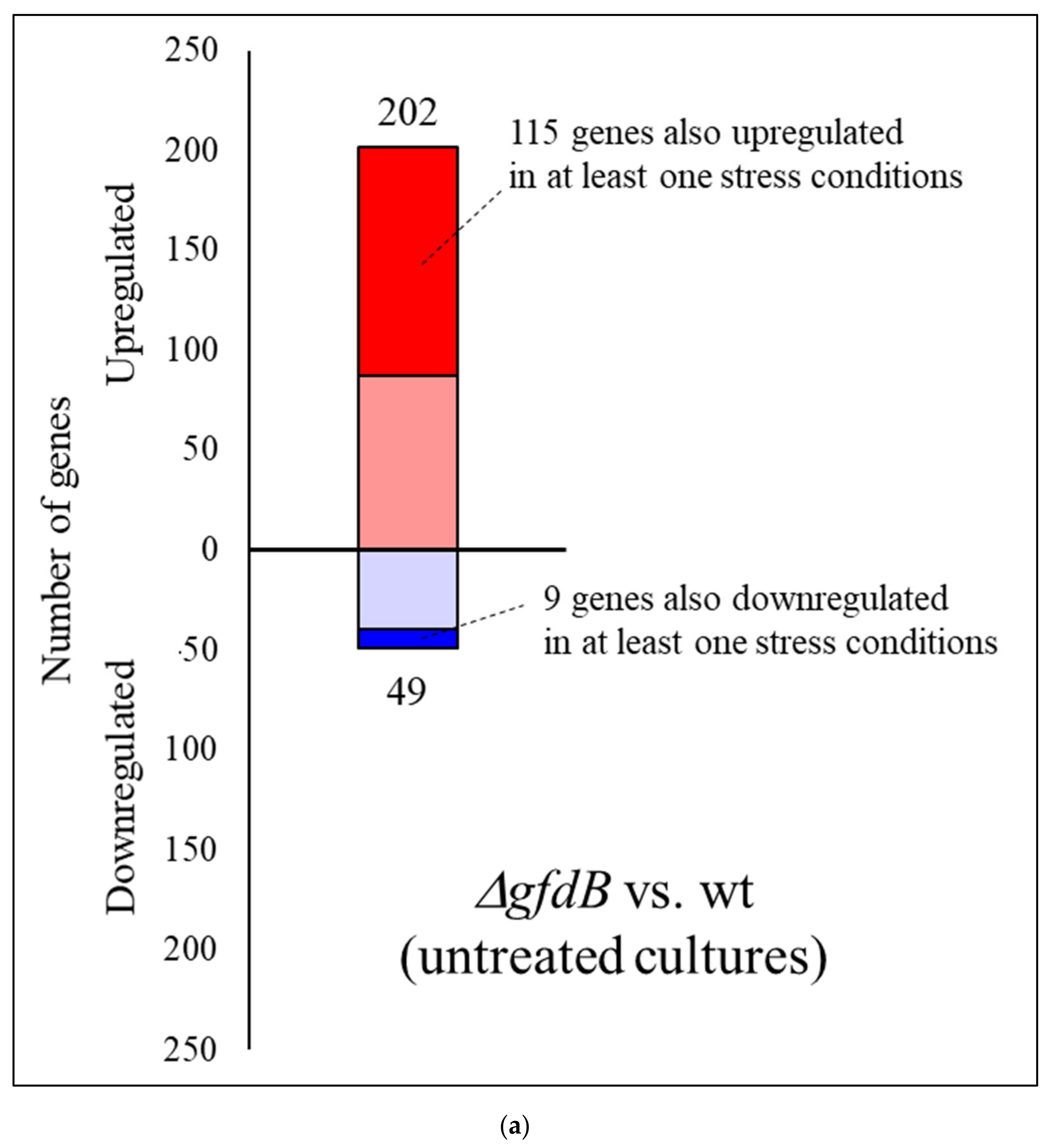
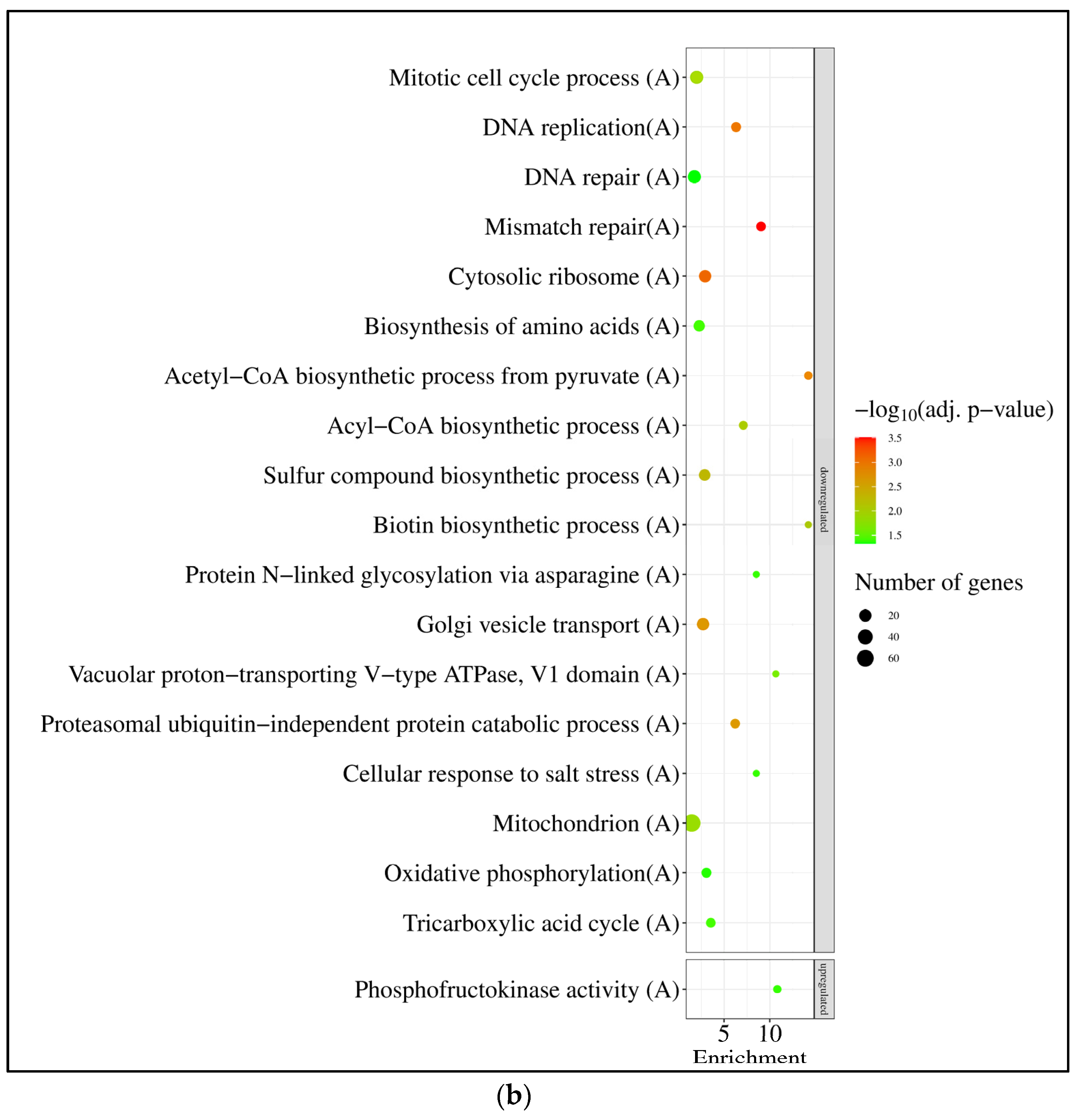
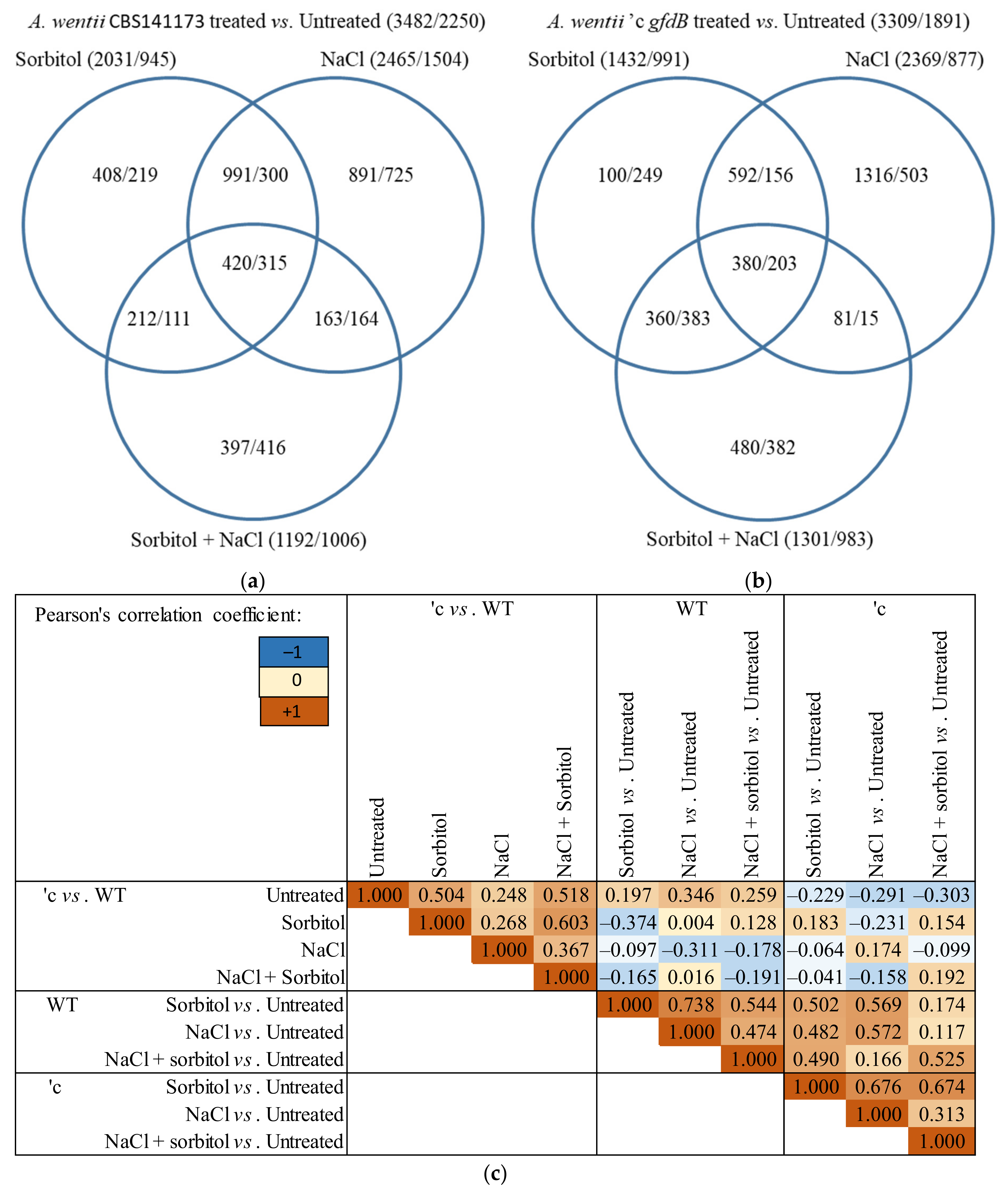
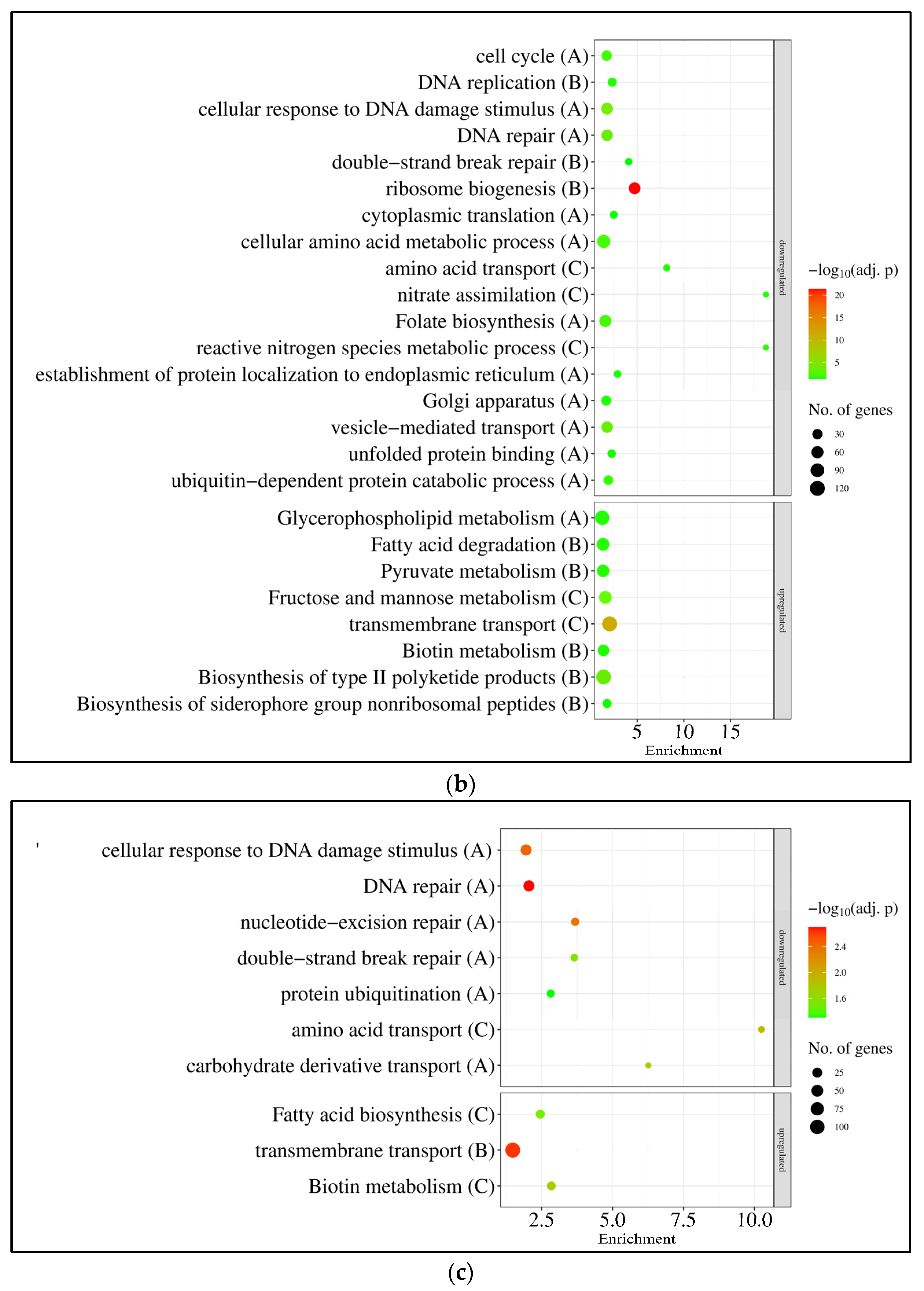
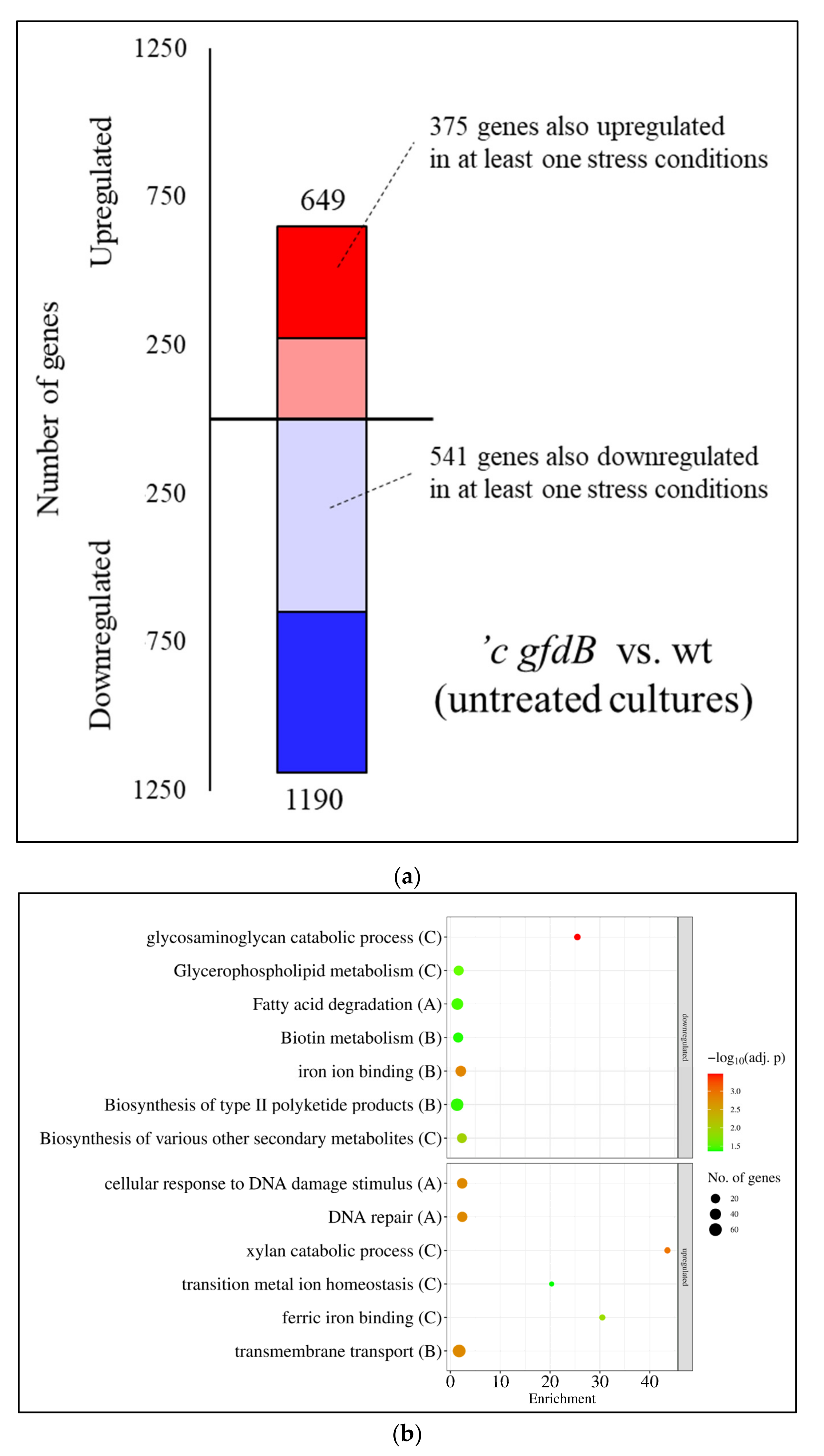
3.3. Expression of A. nidulans gfdB Substantially Disturbed the Physiology of A. wentii
4. Discussion
4.1. A. nidulans THS30 Showed Canonic Hyperosmotic Stress Response under Sorbitol or NaCl Stress
4.2. The Combination of Sorbitol and NaCl Stress Treatments Was Antagonistic
4.3. The Responses of A. wentii CBS141173 to Sorbitol and NaCl Were Not Canonic Hyperosmotic Stress Responses
4.4. GfdB–Easier to Lose Than to Get Back
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kis-Papo, T.; Weig, A.R.; Riley, R.; Peršoh, D.; Salamov, A.; Sun, H.; Lipzen, A.; Wasser, S.P.; Rambold, G.; Grigoriev, I.V.; et al. Genomic adaptations of the halophilic Dead Sea filamentous fungus Eurotium rubrum. Nat. Commun. 2014, 9, 3745. [Google Scholar] [CrossRef]
- Paulussen, C.; Hallsworth, J.E.; Álvarez-Pérez, S.; Nierman, W.C.; Hamill, P.G.; Blain, D.; Rediers, H.; Lievens, B. Ecology of aspergillosis: Insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb. Biotechnol. 2017, 10, 296–322. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.M.; Hilário, S.; Van de Peer, Y.; Esteves, A.C.; Alves, A. Genomic and Metabolomic Analyses of the Marine Fungus Emericellopsis cladophorae: Insights into Saltwater Adaptability Mechanisms and Its Biosynthetic Potential. J. Fungi 2021, 8, 31. [Google Scholar] [CrossRef]
- Yaakoub, H.; Sanchez, N.S.; Ongay-Larios, L.; Courdavault, V.; Calenda, A.; Bouchara, J.P.; Coria, R.; Papon, N. The high osmolarity glycerol (HOG) pathway in fungi†. Crit. Rev. Microbiol. 2022, 48, 657–695. [Google Scholar] [CrossRef] [PubMed]
- Poyatos, F.; Morales, F.; Nicholson, A.W.; Giordano, A. Physiology of biodeterioration on canvas paintings. J. Cell Physiol. 2018, 233, 2741–2751. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.Z.; Sutar, P.P.; Mujumdar, A.S.; Tao, Y.; Pan, Z.; Liu, Y.H.; Xiao, H.W. Thermal Decontamination Technologies for Microorganisms and Mycotoxins in Low-Moisture Foods. Annu. Rev. Food Sci. Technol. 2021, 25, 287–305. [Google Scholar] [CrossRef]
- Al Hallak, M.; Verdier, T.; Bertron, A.; Roques, C.; Bailly, J.D. Fungal Contamination of Building Materials and the Aerosolization of Particles and Toxins in Indoor Air and Their Associated Risks to Health: A Review. Toxins 2023, 25, 175. [Google Scholar] [CrossRef]
- Henderson, A.G.; Ehre, C.; Button, B.; Abdullah, L.H.; Cai, L.H.; Leigh, M.W.; DeMaria, G.C.; Matsui, H.; Donaldson, S.H.; Davis, C.W.; et al. Cystic fibrosis airway secretions exhibit mucin hyperconcentration and increased osmotic pressure. J. Clin. Investig. 2014, 124, 3047–3060. [Google Scholar] [CrossRef]
- Ross, B.S.; Lofgren, L.A.; Ashare, A.; Stajich, J.E.; Cramer, R.A. Aspergillus fumigatus In-Host HOG Pathway Mutation for Cystic Fibrosis Lung Microenvironment Persistence. mBio 2021, 31, e0215321. [Google Scholar] [CrossRef]
- Betlej, G.; Bator, E.; Oklejewicz, B.; Potocki, L.; Górka, A.; Slowik-Borowiec, M.; Czarny, W.; Domka, W.; Kwiatkowska, A. Long-Term Adaption to High Osmotic Stress as a Tool for Improving Enological Characteristics in Industrial Wine Yeast. Genes 2020, 20, 576. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Bourbon-Melo, N.; Sá-Correia, I. The cell wall and the response and tolerance to stresses of biotechnological relevance in yeasts. Front. Microbiol. 2022, 13, 953479. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, X.; Liu, L.; Wu, J.; Gao, C. Engineering Microorganisms to Produce Bio-Based Monomers: Progress and Challenges. Fermentation 2023, 9, 137. [Google Scholar] [CrossRef]
- Duran, R.; Cary, J.W.; Calvo, A.M. Role of the osmotic stress regulatory pathway in morphogenesis and secondary metabolism in filamentous fungi. Toxins 2010, 2, 367–381. [Google Scholar] [CrossRef]
- Wucherpfennig, T.; Hestler, T.; Krull, R. Morphology engineering-osmolarity and its effect on Aspergillus niger morphology and productivity. Microb. Cell Fact. 2011, 10, 58. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Yoshimi, A.; Sakamoto, K.; Gomi, K.; Abe, K.H.; Kitagaki, H. Response and Adaptation to Cell Wall Stress and Osmotic Stress in Aspergillus Species. In Stress Biology of Yeasts and Fungi; Springer: Berlin/Heidelberg, Germany, 2015; pp. 199–218. [Google Scholar] [CrossRef]
- Blomberg, A.; Adler, L. Physiology of osmotolerance in fungi. Adv. Microb. Physiol. 1992, 33, 145–212. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Budge, S.; Kaloriti, D.; Tillmann, A.; Jacobsen, M.D.; Yin, Z.; Ene, I.V.; Bohovych, I.; Sandai, D.; Kastora, S.; et al. Stress adaptation in a pathogenic fungus. J. Exp. Biol. 2014, 217, 144–155. [Google Scholar] [CrossRef] [PubMed]
- de Lima Alves, F.; Stevenson, A.; Baxter, E.; Gillion, J.L.; Hejazi, F.; Hayes, S.; Morrison, I.E.; Prior, B.A.; McGenity, T.J.; Rangel, D.E.; et al. Concomitant osmotic and chaotropicity-induced stresses in Aspergillus wentii: Compatible solutes determine the biotic window. Curr. Genet. 2015, 61, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Sakamoto, K.; Abe, K.; Gomi, K. Signaling pathways for stress responses and adaptation in Aspergillus species: Stress biology in the post-genomic era. Biosci. Biotechnol. Biochem. 2016, 80, 1667–1680. [Google Scholar] [CrossRef]
- de Vries, R.P.; Flitter, S.J.; van de Vondervoort, P.J.; Chaveroche, M.K.; Fontaine, T.; Fillinger, S.; Ruijter, G.J.; d’Enfert, C.; Visser, J. Glycerol dehydrogenase, encoded by gldB is essential for osmotolerance in Aspergillus nidulans. J. Mol. Microbiol. 2003, 49, 131–141. [Google Scholar] [CrossRef]
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Szarvas, V.; Orosz, E.; Antal, K.; Park, H.; Han, K.H.; Yu, J.H.; Pócsi, I. Core oxidative stress response in Aspergillus nidulans. BMC Genomics. 2015, 16, 478. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Király, A.; Hámori, C.; Gyémánt, G.; Kövér, K.E.; Pócsi, I.; Leiter, É. Characterization of gfdB.; putatively encoding a glycerol 3-phosphate dehydrogenase in Aspergillus nidulans. Fungal Biol. 2020, 124, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, S.; Ruijter, G.; Tamás, M.J.; Visser, J.; Thevelein, J.M.; d’Enfert, C. Molecular and physiological characterization of the NAD-dependent glycerol 3-phosphate dehydrogenase in the filamentous fungus Aspergillus nidulans. Mol. Microbiol. 2001, 39, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, X.; Gu, H.; Ma, Z.; Lu, L. Predicted Glycerol 3-Phosphate Dehydrogenase Homologs and the Glycerol Kinase GlcA Coordinately Adapt to Various Carbon Sources and Osmotic Stress in Aspergillus fumigatus. G3 2018, 8, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Király, A.; Szabó, I.G.; Emri, T.; Leiter, É.; Pócsi, I. Supplementation of Aspergillus glaucus with gfdB gene encoding a glycerol 3-phosphate dehydrogenase in Aspergillus nidulans. J. Basic. Microbiol. 2020, 60, 691–698. [Google Scholar] [CrossRef]
- Bodnár, V.; Király, A.; Orosz, E.; Miskei, M.; Emri, T.; Karányi, Z.; Leiter, É.; de Vries, R.P.; Pócsi, I. Species-specific effects of the introduction of Aspergillus nidulans gfdB in osmophilic aspergilli. Appl. Microbiol. Biotechnol. 2023, 107, 2423–2436. [Google Scholar] [CrossRef] [PubMed]
- Tamás, M.J.; Hohmann, S. The osmotic stress response of Saccharomyces cerevisiae. In Yeast Stress Responses; Hohmann, S., Mager, W.H., Eds.; Topics in Current Genetics; Springer: Berlin/Heidelberg, Germany, 2003; Volume 1, pp. 121–200. [Google Scholar] [CrossRef]
- Wadskog, I.; Adler, L. Ion homeostasis in Saccharomyces cerevisiae under NaCl stress. In Yeast Stress Responses; Hohmann, S., Mager, W.H., Eds.; Topics in Current Genetics; Springer: Berlin/Heidelberg, Germany, 2003; Volume 1, pp. 201–239. [Google Scholar] [CrossRef]
- Barratt, R.W.; Johnson, G.B.; Ogata, W.N. Wild-Type and Mutant Stocks of Aspergillus nidulans. Genetics 1965, 52, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Man, Y.Y.; Lee, M.K.; Maeng, J.P.; Chang Kim, S.; Yu, J.H. Velvet-mediated repression of β-glucan synthesis in Aspergillus nidulans spores. Sci. Rep. 2015, 5, 10199. [Google Scholar] [CrossRef]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 536–537. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Prade, R.A. Osmotic stress-coupled maintenance of polar growth in Aspergillus nidulans. Mol. Microbiol. 2002, 43, 1065–1078. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus nidulans HOG pathway is activated only by two-component signalling pathway in response to osmotic stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Asano, Y.; Marui, J.; Furukawa, K.; Kanamaru, K.; Kato, M.; Abe, K.; Kobayashi, T.; Yamashino, T.; Mizuno, T. The SskA and SrrA response regulators are implicated in oxidative stress responses of hyphae and asexual spores in the phosphorelay signaling network of Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2007, 71, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Asano, Y.; Marui, J.; Yoshimi, A.; Mizuno, T.; Abe, K. Transcriptional profiling for Aspergillus nidulans HogA MAPK signaling pathway in response to fludioxonil and osmotic stress. Fungal Genet. Biol. 2009, 46, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Mizuno, T.; Abe, K. Characterization of NikA histidine kinase and two response regulators with special reference to osmotic adaptation and asexual development in Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2009, 73, 1566–1571. [Google Scholar] [CrossRef]
- Lara-Rojas, F.; Sánchez, O.; Kawasaki, L. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses; development and spore functions. J. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef]
- Furukawa, K.; Yoshimi, A.; Furukawa, T.; Hoshi, Y.; Hagiwara, D.; Sato, N.; Fujioka, T.; Mizutani, O.; Mizuno, T.; Kobayashi, T.; et al. Novel reporter gene expression systems for monitoring activation of the Aspergillus nidulans HOG pathway. Biosci. Biotechnol. Biochem. 2007, 71, 1724–1730. [Google Scholar] [CrossRef] [PubMed]
- Fréalle, E.; Noël, C.; Viscogliosi, E.; Camus, D.; Dei-Cas, E.; Delhaes, L. Manganese superoxide dismutase in pathogenic fungi: An issue with pathophysiological and phylogenetic involvements. FEMS Immunol. Med. Microbiol. 2005, 45, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Navarro, R.E.; Stringer, M.A.; Hansberg, W.; Timberlake, W.E.; Aguirre, J. catA, a new Aspergillus nidulans gene encoding a developmentally regulated catalase. Curr. Genet. 1996, 29, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Shimizu, M.; Hoshino, T.; Takaya, N. The glutathione system of Aspergillus nidulans involves a fungus-specific glutathione S-transferase. J. Biol. Chem. 2009, 284, 8042–8053. [Google Scholar] [CrossRef] [PubMed]
- Redkar, R.J.; Locy, R.D.; Singh, N.K. Biosynthetic pathways of glycerol accumulation under salt stress in Aspergillus nidulans. Exp. Mycol. 1995, 19, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Borgia, P.T.; Miao, Y.; Dodge, C.L. The orlA gene from Aspergillus nidulans encodes a trehalose-6-phosphate phosphatase necessary for normal growth and chitin synthesis at elevated temperatures. Mol. Microbiol. 1996, 20, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, S.; Chaveroche, M.K.; van Dijck, P.; de Vries, R.; Ruijter, G.; Thevelein, J.; d’Enfert, C. Trehalose is required for the acquisition of tolerance to a variety of stresses in the filamentous fungus Aspergillus nidulans. Microbiology 2001, 147, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Zähringer, H.; Thevelein, J.M.; Nwaka, S. Induction of neutral trehalase Nth1 by heat and osmotic stress is controlled by STRE elements and Msn2/Msn4 transcription factors: Variations of PKA effect during stress and growth. Mol. Microbiol. 2000, 35, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Pereira Silva, L.; Alves de Castro, P.; Dos Reis, T.F.; Paziani, M.H.; Von Zeska Kress, M.R.; Riaño-Pachón, D.M.; Hagiwara, D.; Ries, L.N.; Brown, N.A.; Goldman, G.H. Genome-wide transcriptome analysis of Aspergillus fumigatus exposed to osmotic stress reveals regulators of osmotic and cell wall stresses that are SakAHOG1 and MpkC dependent. Cell Microbiol. 2017, 19, e12681. [Google Scholar] [CrossRef]
- Vargas-Pérez, I.; Sánchez, O.; Kawasaki, L.; Georgellis, D. Response regulators SrrA and SskA are central components of a phosphorelay system involved in stress signal transduction and asexual sporulation in Aspergillus nidulans. J. Eukaryot. Cell. 2007, 6, 1570–1583. [Google Scholar] [CrossRef]
- Gasch, A.P. Comparative genomics of the environmental stress response in ascomycete fungi. Yeast 2007, 24, 961–976. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T.; Sasse, C.; Schedler, A.; Hasenberg, M.; Gunzer, M.; Krappmann, S. Shaping the fungal adaptome-stress responses of Aspergillus fumigatus. Int. J. Med. Microbiol. 2011, 301, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R. Salt tolerance in plants and microorganisms: Toxicity targets and defense responses. Int. Rev. Cytol. 1996, 165, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Murguía, J.R.; Bellés, J.M.; Serrano, R. The yeast HAL2 nucleotidase is an in vivo target of salt toxicity. J. Biol. Chem. 1996, 271, 29029–29033. [Google Scholar] [CrossRef] [PubMed]
- Ke, R.; Ingram, P.J.; Haynes, K. An integrative model of ion regulation in yeast. PLoS Comput. Biol. 2013, 9, e1002879. [Google Scholar] [CrossRef] [PubMed]
- Ben Hmad, I.; Gargouri, A. Halophilic filamentous fungi and their enzymes: Potential biotechnological applications. J. Biotechnol. 2024, 381, 11–18. [Google Scholar] [CrossRef]
- Musa, H.; Kasim, F.H.; Gunny, A.A.N.; Gopinath, S.C.B. Salt-adapted moulds and yeasts: Potentials in industrial and environmental biotechnology. Proc. Biochem. 2018, 69, 33–44. [Google Scholar] [CrossRef]
- Ibrar, M.; Ullah, M.W.; Manan, S.; Farooq, U.; Rafiq, M.; Hasan, F. Fungi from the extremes of life: An untapped treasure for bioactive compounds. Appl. Microbiol. Biotechnol. 2020, 104, 2777–2801. [Google Scholar] [CrossRef] [PubMed]
- Śliżewska, W.; Struszczyk-Świta, K.; Marchut-Mikołajczyk, O. Metabolic potential of halophilic filamentous fungi-current perspective. Int. J. Mol. Sci. 2022, 23, 4189. [Google Scholar] [CrossRef]
- Leong, S.L.; Lantz, H.; Pettersson, O.V.; Frisvad, J.C.; Thrane, U.; Heipieper, H.J.; Dijksterhuis, J.; Grabherr, M.; Pettersson, M.; Tellgren-Roth, C.; et al. Genome and physiology of the ascomycete filamentous fungus Xeromyces bisporus, the most xerophilic organism isolated to date. Environ. Microbiol. 2014, 17, 496–513. [Google Scholar] [CrossRef]
- Fernando, L.D.; Pérez-Llano, Y.; Dickwella Widanage, M.C.; Jacob, A.; Martínez-Ávila, L.; Lipton, A.S.; Gunde-Cimerman, N.; Latgé, J.P.; Batista-García, R.A.; Wang, T. Structural adaptation of fungal cell wall in hypersaline environment. Nat. Commun. 2023, 14, 7082. [Google Scholar] [CrossRef]
- Chander, H.; Batish, V.K.; Sannabhadti, S.S.; Srinivasan, R.A. Factors affecting lipase production in Aspergillus wentii. J. Food Sci. 1980, 45, 598–600. [Google Scholar] [CrossRef]
- Li, X.; Li, X.M.; Xu, G.M.; Li, C.S.; Wang, B.G. Antioxidant metabolites from marine alga-derived fungus Aspergillus wentii EN-48. Phytochem. Lett. 2014, 7, 120–123. [Google Scholar] [CrossRef]
- Shoaib, A.; Bhran, A.; Rasmey, A.H.; Mikky, Y. Optimization of cultural conditions for lipid accumulation by Aspergillus wentii Ras101 and its transesterification to biodiesel: Application of response surface methodology. 3 Biotech. 2018, 8, 417. [Google Scholar] [CrossRef] [PubMed]
- Hasan, H.A. Studies on toxigenic fungi in roasted foodstuff (salted seed) and halotolerant activity of emodin-producing Aspergillus wentii. Folia Microbiol. 1998, 43, 383–391. [Google Scholar] [CrossRef]
- Halsey, C.; Lumley, H.; Luckit, J. Necrotising external otitis caused by Aspergillus wentii: A case report. Mycoses 2011, 54, e211–e213. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Reference |
|---|---|---|
| A. nidulans THS30 | pyrG89, AfupyrG+; pyroA+; veA+ | [32] |
| A. nidulans ΔgfdB | pyrG89; ΔgfdB::AfupyrG+; pyroA+; veA+ | [24] |
| A. wentii CBS141173 | Wild-type | [22] |
| A. wentii ′c gfdB | CBS141173 strain harboring one copy of A. nidulans gfdB with its own promoter and terminator sequences and HygR as a selection marker gene | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodnár, V.; Antal, K.; de Vries, R.P.; Pócsi, I.; Emri, T. Aspergillus nidulans gfdB, Encoding the Hyperosmotic Stress Protein Glycerol-3-phosphate Dehydrogenase, Disrupts Osmoadaptation in Aspergillus wentii. J. Fungi 2024, 10, 291. https://doi.org/10.3390/jof10040291
Bodnár V, Antal K, de Vries RP, Pócsi I, Emri T. Aspergillus nidulans gfdB, Encoding the Hyperosmotic Stress Protein Glycerol-3-phosphate Dehydrogenase, Disrupts Osmoadaptation in Aspergillus wentii. Journal of Fungi. 2024; 10(4):291. https://doi.org/10.3390/jof10040291
Chicago/Turabian StyleBodnár, Veronika, Károly Antal, Ronald P. de Vries, István Pócsi, and Tamás Emri. 2024. "Aspergillus nidulans gfdB, Encoding the Hyperosmotic Stress Protein Glycerol-3-phosphate Dehydrogenase, Disrupts Osmoadaptation in Aspergillus wentii" Journal of Fungi 10, no. 4: 291. https://doi.org/10.3390/jof10040291