
A New Overview of Sex Bias in Fungal Infections


Abstract
1. Introduction
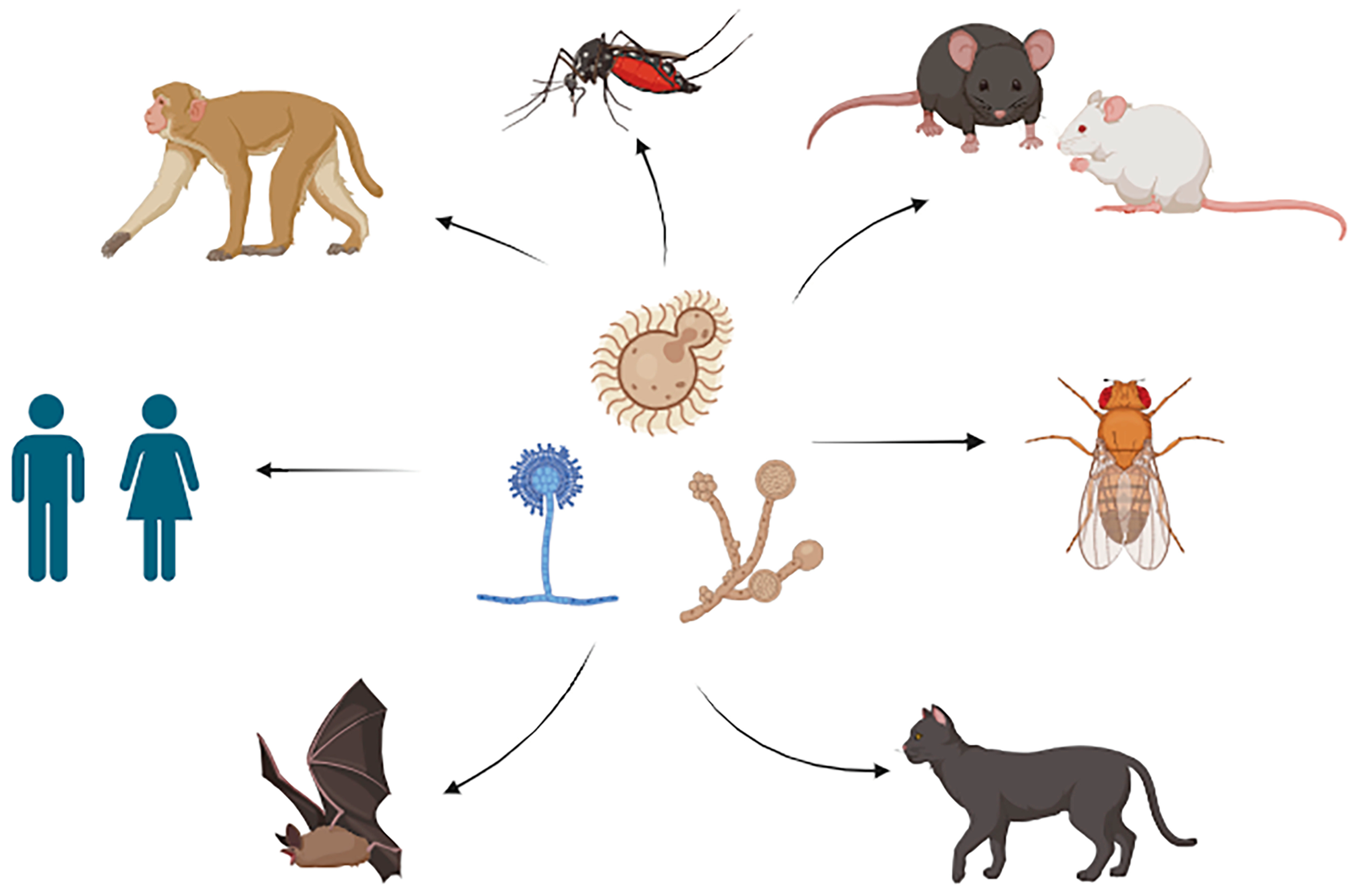
2. Sex Differences in Response to Fungal Infections That Cause Disease in Mammals and Invertebrates
2.1. Mice
2.2. Drosophila
2.3. Cats
2.4. Bats
3. Sex Differences in Responses to Fungal Infections That Cause Disease in Humans
3.1. C. neoformans
3.2. Tinea barbae and T. capitis
4. Sex Differences in Response to Disease-Associated Disruptions in the Mycobiome
4.1. Humans
4.2. Primates
4.3. Mosquitoes
5. Further Directions and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naranjo-Ortiz, M.A.; Gabaldon, T. Fungal evolution: Diversity, taxonomy and phylogeny of the Fungi. Biol. Rev. Camb. Philos. Soc. 2019, 94, 2101–2137. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.J.; Ray, C.G. Sherris Medical Microbiology: An Introduction to Infectious Diseases, 4th ed.; McGraw-Hill Medical: Chicago, IL, USA, 2003. [Google Scholar]
- Walsh, T.J.; Dixon, D.M. Spectrum of Mycoses. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston Galveston (TX): Galveston, TX, USA, 1996. [Google Scholar]
- Fungal Disease Frequency. Global Action for Fungal Infections. Available online: https://gaffi.org/why/fungal-disease-frequency/ (accessed on 6 August 2024).
- Denning, D.W. Global incidence and mortality of severe fungal disease. Lancet Infect. Dis. 2024, 24, e428–e438. [Google Scholar] [CrossRef]
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal infections in humans: The silent crisis. Microb. Cell 2020, 7, 143–145. [Google Scholar] [CrossRef]
- Rhodes, J.; Abdolrasouli, A.; Dunne, K.; Sewell, T.R.; Zhang, Y.; Ballard, E.; Brackin, A.P.; van Rhijn, N.; Chown, H.; Tsitsopoulou, A.; et al. Population genomics confirms acquisition of drug-resistant Aspergillus fumigatus infection by humans from the environment. Nat. Microbiol. 2022, 7, 663–674. [Google Scholar] [CrossRef]
- Duong, T.N.; Le, T.V.; Tran, K.H.; Nguyen, P.T.; Nguyen, B.T.; Nguyen, T.A.; Nguyen, H.P.; Nguyen, B.T.; Fisher, M.C.; Rhodes, J.; et al. Azole-resistant Aspergillus fumigatus is highly prevalent in the environment of Vietnam, with marked variability by land use type. Environ. Microbiol. 2021, 23, 7632–7642. [Google Scholar] [CrossRef]
- Benedict, K.; Park, B.J. Invasive fungal infections after natural disasters. Emerg. Infect. Dis. 2014, 20, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Nnadi, N.E.; Carter, D.A. Climate change and the emergence of fungal pathogens. PLoS Pathog. 2021, 17, e1009503. [Google Scholar] [CrossRef]
- Thompson, G.R., 3rd; Chiller, T.M. Endemic mycoses: Underdiagnosed and underreported. Ann. Intern. Med. 2022, 175, 1759–1760. [Google Scholar] [CrossRef] [PubMed]
- Guess, T.E.; McClelland, E.E. Chapter 29—An Updated Overview of the Gender-Specific Response to Infection. In Principles of Gender-Specific Medicine, 3rd ed.; Legato, M.J., Ed.; Academic Press: San Diego, CA, USA, 2017. [Google Scholar]
- Guess, T.E.; Rosen, J.A.; McClelland, E.E. An overview of sex bias in C. neoformans infections. J. Fungi 2018, 4, 49. [Google Scholar] [CrossRef]
- McClelland, E.E.; Smith, J.M. Gender specific differences in the immune response to infection. Arch. Immunol. Ther. Exp. 2011, 59, 203–213. [Google Scholar] [CrossRef]
- Arnegard, M.E.; Whitten, L.A.; Hunter, C.; Clayton, J.A. Sex as a biological variable: A 5-year progress report and call to action. J. Womens Health 2020, 29, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.L.; Ceesay, M.; Leier, J.A.; Tesch, J.; Wisenden, B.D.; Pandey, S. Factors contributing to sex differences in mice inhaling Aspergillus fumigatus. Int. J. Environ. Res. Public Health 2020, 17, 8851. [Google Scholar] [CrossRef]
- Klein, S.L. Immune cells have sex and so should journal articles. Endocrinology 2012, 153, 2544–2550. [Google Scholar] [CrossRef] [PubMed]
- Dromer, F.; Mathoulin-Pelissier, S.; Launay, O.; Lortholary, O. Determinants of disease presentation and outcome during cryptococcosis: The CryptoA/D study. PLoS Med. 2007, 4, e21. [Google Scholar] [CrossRef]
- McClelland, E.E.; Hobbs, L.M.; Rivera, J.; Casadevall, A.; Potts, W.K.; Smith, J.M.; Ory, J.J. The role of host gender in the pathogenesis of Cryptococcus neoformans infections. PLoS ONE 2013, 8, e63632. [Google Scholar] [CrossRef]
- Arroyo-Mendoza, M.; Peraza, K.; Olson, J.; Adler-Moore, J.P.; Buckley, N.E. Effect of testosterone and estrogen supplementation on the resistance to systemic Candida albicans infection in mice. Heliyon 2020, 6, e04437. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L. The effects of hormones on sex differences in infection: From genes to behavior. Neurosci. Biobehav. Rev. 2000, 24, 627–638. [Google Scholar] [CrossRef]
- Aristizabal, B.H.; Clemons, K.V.; Cock, A.M.; Restrepo, A.; Stevens, D.A. Experimental Paracoccidioides brasiliensis infection in mice: Influence of the hormonal status of the host on tissue responses. Med. Mycol. 2002, 40, 169–178. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, F.; Tompkins, K.C.; McNamara, A.; Jain, A.V.; Moore, B.B.; Toews, G.B.; Huffnagle, G.B.; Olszewski, M.A. Robust Th1 and Th17 immunity supports pulmonary clearance but cannot prevent systemic dissemination of highly virulent Cryptococcus neoformans H99. Am. J. Pathol. 2009, 175, 2489–2500. [Google Scholar] [CrossRef]
- Belmonte, R.L.; Corbally, M.K.; Duneau, D.F.; Regan, J.C. Sexual dimorphisms in innate immunity and responses to infection in Drosophila melanogaster. Front. Immunol. 2019, 10, 3075. [Google Scholar] [CrossRef]
- Shahrestani, P.; Chambers, M.; Vandenberg, J.; Garcia, K.; Malaret, G.; Chowdhury, P.; Estrella, Y.; Zhu, M.; Lazzaro, B.P. Sexual dimorphism in Drosophila melanogaster survival of Beauveria bassiana infection depends on core immune signaling. Sci. Rep. 2018, 8, 12501. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.L.; Wang, J.B.; Brown, M.A.; Euerle, C.; St Leger, R.J. Identification of Drosophila mutants affecting defense to an entomopathogenic fungus. Sci. Rep. 2015, 5, 12350. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.; Li, C.F.; He, Z.; Lu, Y.; Liu, X.S.; Wang, Y.F.; Ip, Y.T.; Strand, M.R.; Yu, X.Q. Toll family members bind multiple Spatzle proteins and activate antimicrobial peptide gene expression in Drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef] [PubMed]
- Folstad, I.; Karter, A.J. Parasites, bright males, and the immunocompetence handicap. Am. Nat. 1992, 139, 603–622. [Google Scholar] [CrossRef]
- Zuk, M.; Stoehr, A.M. Immune defense and host life history. Am. Nat. 2002, 160 (Suppl. 4), S9–S22. [Google Scholar] [CrossRef]
- Proverbio, D.; Perego, R.; Spada, E.; Bagnagatti de Giorgi, G.; Della Pepa, A.; Ferro, E. Survey of dermatophytes in stray cats with and without skin lesions in Northern Italy. Vet. Med. Int. 2014, 2014, 565470. [Google Scholar] [CrossRef]
- Paryuni, A.D.; Indarjulianto, S.; Widyarini, S. Dermatophytosis in companion animals: A review. Vet. World 2020, 13, 1174–1181. [Google Scholar] [CrossRef]
- Moskaluk, A.E.; VandeWoude, S. Current topics in dermatophyte classification and clinical diagnosis. Pathogens 2022, 11, 957. [Google Scholar] [CrossRef]
- Lionakis, M.S.; Kontoyiannis, D.P. Glucocorticoids and invasive fungal infections. Lancet 2003, 362, 1828–1838. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Chrousos, G.P. Stress hormones, Th1/Th2 patterns, pro/anti-inflammatory cytokines and susceptibility to disease. Trends Endocrinol. Metab. 1999, 10, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Kailing, M.J.; Hoyt, J.R.; White, J.P.; Kaarakka, H.M.; Redell, J.A.; Leon, A.E.; Rocke, T.E.; DePue, J.E.; Scullon, W.H.; Parise, K.L.; et al. Sex-biased infections scale to population impacts for an emerging wildlife disease. Proc. Biol. Sci. 2023, 290, 20230040. [Google Scholar] [CrossRef] [PubMed]
- McGuire, L.P.; Mayberry, H.W.; Willis, C.K.R. White-nose syndrome increases torpid metabolic rate and evaporative water loss in hibernating bats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R680–R686. [Google Scholar] [CrossRef]
- Krueger, S.K.; Williams, S.C.; O’Keefe, J.M.; Zirkle, G.A.; Haase, C.G. White-nose syndrome, winter duration, and pre-hibernation climate impact abundance of reproductive female bats. PLoS ONE 2024, 19, e0298515. [Google Scholar] [CrossRef]
- Russo, D.; Bosso, L.; Ancillotto, L. Novel perspectives on bat insectivory highlight the value of this ecosystem service in farmland: Research frontiers and management implications. Agric. Ecosyst. Environ. 2018, 266, 31–38. [Google Scholar] [CrossRef]
- Czenze, Z.J.; Jonasson, K.A.; Willis, C.K.R. Thrifty females, frisky males: Winter energetics of hibernating bats from a cold climate. Physiol. Biochem. Zool. 2017, 90, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, K.A.; Willis, C.K. Changes in body condition of hibernating bats support the thrifty female hypothesis and predict consequences for populations with white-nose syndrome. PLoS ONE 2011, 6, e21061. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, B.J.; Freeman, D.A.; Zucker, I.; Nelson, R.J. Periodic arousal from hibernation is necessary for initiation of immune responses in ground squirrels. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R1054–R1062. [Google Scholar] [CrossRef] [PubMed]
- Bouma, H.R.; Carey, H.V.; Kroese, F.G. Hibernation: The immune system at rest? J. Leukoc. Biol. 2010, 88, 619–624. [Google Scholar] [CrossRef]
- Shaheen, A.A.; Somayaji, R.; Myers, R.; Mody, C.H. Epidemiology and trends of cryptococcosis in the United States from 2000 to 2007: A population-based study. Int. J. STD AIDS 2018, 29, 453–460. [Google Scholar] [CrossRef]
- Egger, M.; Hoenigl, M.; Thompson, G.R., 3rd; Carvalho, A.; Jenks, J.D. Let’s talk about sex characteristics-as a risk factor for invasive fungal diseases. Mycoses 2022, 65, 599–612. [Google Scholar] [CrossRef]
- Bilbey, D.L.; Nicol, T. Effect of various natural steroids on the phagocytic activity of the reticuloendothelial system. Nature 1958, 182, 674. [Google Scholar] [CrossRef]
- Grossman, C. Possible underlying mechanisms of sexual dimorphism in the immune response, fact and hypothesis. J. Steroid Biochem. 1989, 34, 241–251. [Google Scholar] [CrossRef]
- Mohr, J.A.; Tacker, R.J.; Devlin, R.F.; Felton, F.G.; Rhoades, E.R.; Muchmore, H.G. Estrogen-stimulated phagocytic activity in human cryptococcosis. Am. Rev. Respir. Dis. 1969, 99, 979. [Google Scholar]
- Mohr, J.A.; Muchmore, H.G.; Tacker, R. Stimulation of phagocytosis of Cryptococcus neoformans in human cryptococcal meningitis. J. Reticuloendothel. Soc. 1974, 15, 149–154. [Google Scholar]
- Mohr, J.A.; Long, H.; McKown, B.A.; Muchmore, H.G. In vitro susceptibility of Cryptococcus neoformans to steroids. Sabouraudia 1972, 10, 171–172. [Google Scholar] [CrossRef]
- Mohr, J.A.; Tatem, B.A.; Long, H.; Muchmore, H.G.; Felton, F.G. Increased susceptibility of Cryptococcus neoformans to amphotericin B in the presence of steroids. Sabouraudia 1973, 11, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Krasevec, N. The multifaceted role of mating type of the fungus and sex of the host in studies of fungal infections in humans. J. Fungi 2022, 8, 461. [Google Scholar] [CrossRef] [PubMed]
- Giefing-Kroll, C.; Berger, P.; Lepperdinger, G.; Grubeck-Loebenstein, B. How sex and age affect immune responses, susceptibility to infections, and response to vaccination. Aging Cell 2015, 14, 309–321. [Google Scholar] [CrossRef]
- Lasarte, S.; Samaniego, R.; Salinas-Munoz, L.; Guia-Gonzalez, M.A.; Weiss, L.A.; Mercader, E.; Ceballos-Garcia, E.; Navarro-Gonzalez, T.; Moreno-Ochoa, L.; Perez-Millan, F.; et al. Sex hormones coordinate neutrophil immunity in the vagina by controlling chemokine gradients. J. Infect. Dis. 2016, 213, 476–484. [Google Scholar] [CrossRef]
- Blotta, M.H.; Mamoni, R.L.; Oliveira, S.J.; Nouer, S.A.; Papaiordanou, P.M.; Goveia, A.; Camargo, Z.P. Endemic regions of paracoccidioidomycosis in Brazil: A clinical and epidemiologic study of 584 cases in the southeast region. Am. J. Trop. Med. Hyg. 1999, 61, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Loose, D.S.; Stover, E.P.; Restrepo, A.; Stevens, D.A.; Feldman, D. Estradiol binds to a receptor-like cytosol binding protein and initiates a biological response in Paracoccidioides brasiliensis. Proc. Natl. Acad. Sci. USA 1983, 80, 7659–7663. [Google Scholar] [CrossRef] [PubMed]
- Guess, T.E.; Rosen, J.; Castro-Lopez, N.; Wormley, F.L., Jr.; McClelland, E.E. An inherent T cell deficit in healthy males to C. neoformans infection may begin to explain the sex susceptibility in incidence of cryptococcosis. Biol. Sex. Differ. 2019, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, C.M.; Romao, F.; Castelo-Branco, C. Menopause and aging: Changes in the immune system--A review. Maturitas 2010, 67, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Galhardas, C.; Cabete, J. Adult Tinea capitis and Tinea barbae in a tertiary Portuguese hospital: A 11-year audit. Mycoses 2019, 62, 1079–1083. [Google Scholar] [CrossRef]
- Dey, V.K. Misuse of topical corticosteroids: A clinical study of adverse effects. Indian. Dermatol. Online J. 2014, 5, 436–440. [Google Scholar] [CrossRef]
- Belvoncikova, P.; Splichalova, P.; Videnska, P.; Gardlik, R. The human mycobiome: Colonization, composition and the role in health and disease. J. Fungi 2022, 8, 1046. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.M.; de Paula, T.O.; Sarmiento, M.R.A.; de Oliveira, M.G.; Pereira, M.L.M.; Toledo, I.V.; Nascimento, T.C.; Ferreira-Machado, A.B.; Silva, V.L.; Diniz, C.G. Fungal diversity of human gut microbiota among eutrophic, overweight, and obese individuals based on aerobic culture-dependent approach. Curr. Microbiol. 2018, 75, 726–735. [Google Scholar] [CrossRef]
- Mar Rodriguez, M.; Perez, D.; Javier Chaves, F.; Esteve, E.; Marin-Garcia, P.; Xifra, G.; Vendrell, J.; Jove, M.; Pamplona, R.; Ricart, W.; et al. Obesity changes the human gut mycobiome. Sci. Rep. 2015, 5, 14600. [Google Scholar] [CrossRef]
- Gomez-Eguilaz, M.; Ramon-Trapero, J.L.; Perez-Martinez, L.; Blanco, J.R. The microbiota-gut-brain axis and its great projections. Rev. Neurol. 2019, 68, 111–117. [Google Scholar]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef]
- Stopinska, K.; Radziwon-Zaleska, M.; Domitrz, I. The microbiota-gut-brain axis as a key to neuropsychiatric disorders: A mini review. J. Clin. Med. 2021, 10, 4640. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Severance, E.G.; Gressitt, K.L.; Stallings, C.R.; Katsafanas, E.; Schweinfurth, L.A.; Savage, C.L.G.; Adamos, M.B.; Sweeney, K.M.; Origoni, A.E.; Khushalani, S.; et al. Probiotic normalization of Candida albicans in schizophrenia: A randomized, placebo-controlled, longitudinal pilot study. Brain Behav. Immun. 2017, 62, 41–45. [Google Scholar] [CrossRef]
- Severance, E.G.; Alaedini, A.; Yang, S.; Halling, M.; Gressitt, K.L.; Stallings, C.R.; Origoni, A.E.; Vaughan, C.; Khushalani, S.; Leweke, F.M.; et al. Gastrointestinal inflammation and associated immune activation in schizophrenia. Schizophr. Res. 2012, 138, 48–53. [Google Scholar] [CrossRef]
- Li, H.; Miao, M.X.; Jia, C.L.; Cao, Y.B.; Yan, T.H.; Jiang, Y.Y.; Yang, F. Interactions between Candida albicans and the resident microbiota. Front. Microbiol. 2022, 13, 930495. [Google Scholar] [CrossRef]
- Sun, B.; Gu, Z.; Wang, X.; Huffman, M.A.; Garber, P.A.; Sheeran, L.K.; Zhang, D.; Zhu, Y.; Xia, D.P.; Li, J.H. Season, age, and sex affect the fecal mycobiota of free-ranging Tibetan macaques (Macaca thibetana). Am. J. Primatol. 2018, 80, e22880. [Google Scholar] [CrossRef]
- Frankel-Bricker, J. Shifts in the microbiota associated with male mosquitoes (Aedes aegypti) exposed to an obligate gut fungal symbiont (Zancudomyces culisetae). Sci. Rep. 2020, 10, 12886. [Google Scholar] [CrossRef] [PubMed]
- Frankel-Bricker, J.; Buerki, S.; Feris, K.P.; White, M.M. Influences of a prolific gut fungus (Zancudomyces culisetae) on larval and adult mosquito (Aedes aegypti)-associated microbiota. Appl. Environ. Microbiol. 2020, 86, e02334-19. [Google Scholar] [CrossRef]
- Guegan, M.; Martin, E.; Tran Van, V.; Fel, B.; Hay, A.E.; Simon, L.; Butin, N.; Bellvert, F.; Haichar, F.E.Z.; Valiente Moro, C. Mosquito sex and mycobiota contribute to fructose metabolism in the Asian tiger mosquito Aedes albopictus. Microbiome 2022, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Early, A.M.; Shanmugarajah, N.; Buchon, N.; Clark, A.G. Drosophila genotype influences commensal bacterial levels. PLoS ONE 2017, 12, e0170332. [Google Scholar] [CrossRef] [PubMed]
- Kheffache, H.; Seklaoui, N.; Bouchara, J.P.; Boukhemza-Zemmouri, N.; Boukhemza, M. Tinea capitis at the University Hospital of Tizi-Ouzou, Algeria, and first isolation of Trichophyton tonsurans. J. Mycol. Med. 2020, 30, 101040. [Google Scholar] [CrossRef] [PubMed]
- Fink, A.L.; Klein, S.L. The evolution of greater humoral immunity in females than males: Implications for vaccine efficacy. Curr. Opin. Physiol. 2018, 6, 16–20. [Google Scholar] [CrossRef] [PubMed]
- CDC. HIV Surveillance Report, 2011; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2011. [Google Scholar]

{kind=link}
| Organism | Fungi | Sex-Specific Difference | n = Sample Size | Sex Bias Ratio * | Reference(s) |
|---|---|---|---|---|---|
| C57BL/6 Mice | Aspergillus fumigatus | Increased immune responses exhibited in females. | 6–7 mice/ group | 2:1 (F/M) | [16] |
| C57BL/6 Mice | Candida albicans | Males are more susceptible to systemic infection than females. | 10 mice/ group | 1:6.1 (M/F) | [20] |
| BALB/c Mice | Paracoccidioides brasiliensis | Males are more susceptible to infection than female mice. Gonadectomized female mice treated with testosterone showed increased susceptibility to infection. | 3 mice/ group | 2.55:1 (M/F) | [22] |
| BALB/c Mice | Cryptococcus neoformans | Males have a significantly higher fungal burden in the spleen than female mice during chronic infection. | 4–5 mice/ group | 9:1 (M/F) | [19] |
| Drosophila | Beauveria bassiana | Males are more likely to survive infection. | 500–600 flies/group | 1.1:1 (M/F) | [25] |
| Drosophila | Metarhizium anisopliae | Males are more likely to survive infection. | 30 flies/ group | 1.1:1 (M/F) | [26] |
| Drosophila | Candida albicans | Males are more susceptible to systemic challenge via intra-thoracic injection. | 20–30 flies/ group | 4:1 (M/F) | [27] |
| Cats | Microsporum canis | Females are more likely to be infected. | 199 female and 74 male cats | 7:1.4 (F/M) | [30] |
| Bats | Pseudogymnoascus destructans | Females are less likely to be recaptured due to death. | 665 female and 1071 male bats | 0.7:1 (F/M) | [37] |
| Humans | Cryptococcus neoformans | Males have lower CD3+, CD4+, and CD8+ T-cell percentages after infection. | 21 males and 19 females | 2.1:1 (M/F) | [56] |
| Cryptococcus neoformans | Increased phagocytosis observed after treatment with 5 mg diethylstilbestrol. | 6 males and 3 females | 2:1 (M/F) | [48] | |
| Humans | Paracoccidioides brasiliensis | Males are more susceptible to infection. | 492 males and 92 females | 5.3:1 (M/F) | [54] |
| Humans | Tinea barbae | Males are disproportionately affected in number. | 6 males and 1 female | 6:1 (M/F) | [58] |
| Humans | Tinea capitis | Females are disproportionately affected in number. | 9 females and 6 males | 1.5:1 (F/M) | [58] |
| Humans | Tinea capitis | Male children aged 4–6 years comprise 43.3% of infections. | 45 males and 42 females | 1.09:1 (M/F) | [75] |
| Human (Mycobiome) | Candida albicans (Ca) | Levels were elevated in the male mycobiome of those exhibiting symptoms of schizophrenia. | 165 Ca+ males and 219 Ca− males | 1.3:1 | [67] |
| Human (Mycobiome) | Saccharomyces cerevisiae (Sc) | Males with recent-onset schizophrenia had elevated levels of IgG antibodies to Sc. | 51 males and 16 females | 3.2:1 (M/F) | [68] |
| Human (Mycobiome) | Saccharomyces cerevisiae (Sc) | Females with non-recent-onset schizophrenia had elevated levels of IgG antibodies to Sc. | 79 females and 114 males | 1.4:1 (F/M) | [68] |
| Tibetan Macaques | Family: Mycosphaerellaceae; Genus: Devriesia | Females demonstrate significant enrichment of these taxa in the gut microbiota. | 18 females and 13 males | NA | [70] |
| Tibetan Macaques | Phylum: Ascomycota; Family: Tetraplosphaeriaceae | Males demonstrate significant enrichment of these taxa in the gut microbiota. | 18 females and 13 males | NA | [70] |
| Aedes aegypti | Zancudomyces culisetae (Zc) | Males demonstrate higher levels of the Burkholderiaceae family of bacteria in the gut microbiota than females when both sexes are exposed to this fungus during the larval stage. | 12 male mosquitoes and 31 female mosquitoes | 9:1 | [71,72] |
| Aedes albopictus | Aureobasidium genus | This genus is more enriched in the mycobiota post feeding in females at the 4 h mark. | 150 male and female mosquitoes/group | 10:1 (F/M) | [73] |
| Aedes albopictus | Aureobasidium genus | This genus is more enriched in the mycobiota post feeding in males at the 10 h mark. | 150 male and female mosquitoes/group | 3:1 (M/F) | [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, H.H.; McClelland, E.E. A New Overview of Sex Bias in Fungal Infections. J. Fungi 2024, 10, 607. https://doi.org/10.3390/jof10090607
Rao HH, McClelland EE. A New Overview of Sex Bias in Fungal Infections. Journal of Fungi. 2024; 10(9):607. https://doi.org/10.3390/jof10090607
Chicago/Turabian StyleRao, Hari H., and Erin E. McClelland. 2024. "A New Overview of Sex Bias in Fungal Infections" Journal of Fungi 10, no. 9: 607. https://doi.org/10.3390/jof10090607
APA StyleRao, H. H., & McClelland, E. E. (2024). A New Overview of Sex Bias in Fungal Infections. Journal of Fungi, 10(9), 607. https://doi.org/10.3390/jof10090607