
Identification and Biotyping of Pythium insidiosum Isolated from Urban and Rural Areas of Thailand by Multiplex PCR, DNA Barcode, and Proteomic Analyses

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Culture Condition
2.2. Genomic DNA Extraction
2.3. Multiplex PCR for Identification and Genotyping of P. insidiosum
2.4. Species Identification by DNA Barcode Analysis
2.5. Phylogenetic Analysis
2.6. Mass Spectrometric Analysis and Dendrogram
2.7. Data Availability
3. Results
3.1. Screening the Colony Morphology of P. insidiosum Isolated from Water Samples
3.2. Identification and Genotyping of P. insidiosum by Multiplex PCR and DNA Barcodes
3.3. Geographic Distribution of the P. insidiosum-Positive Water Samples
3.4. Phylogenetic Relationship among the Water-Isolated and Clinical Strains of P. insidiosum
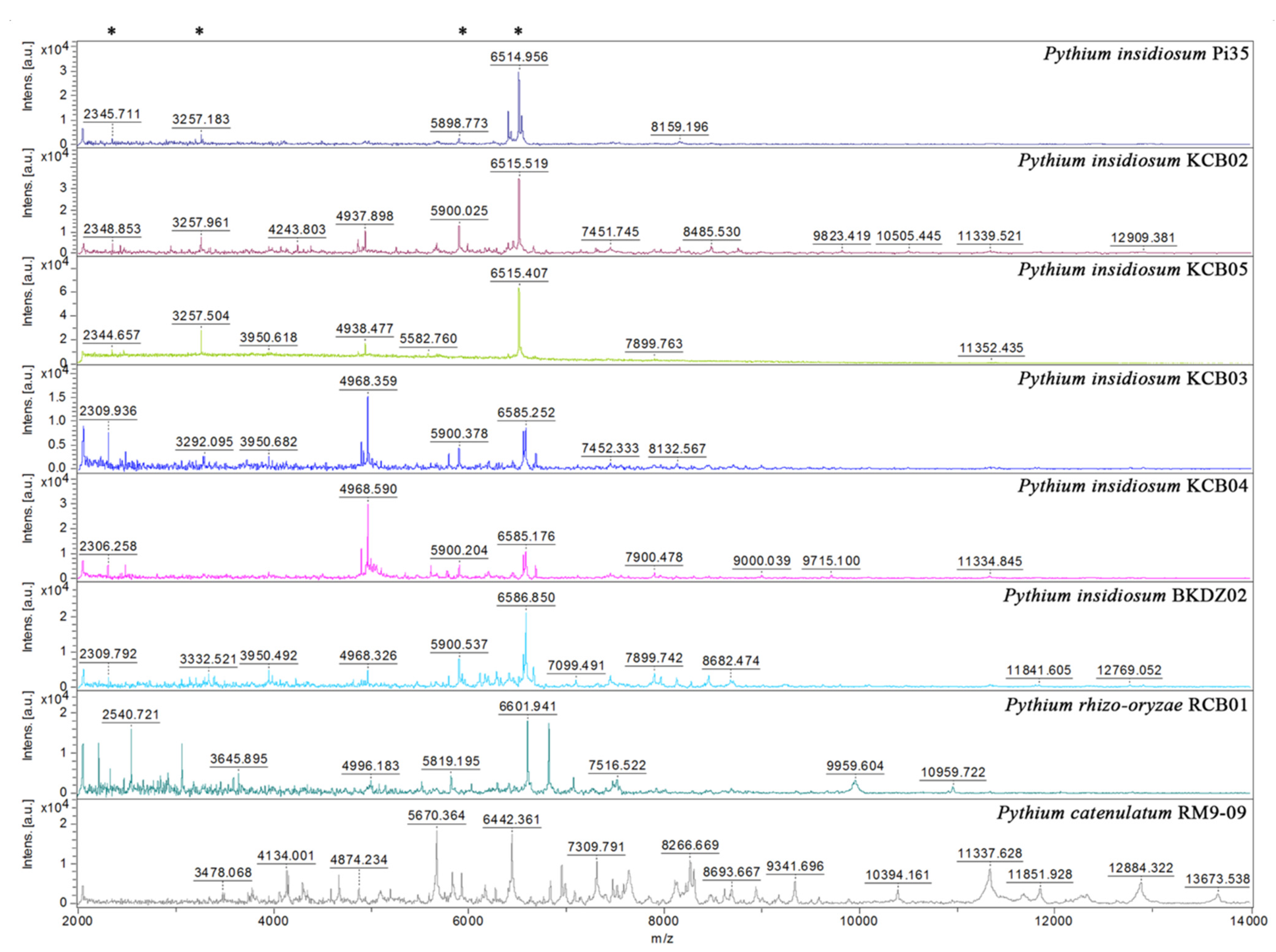
3.5. Mass Spectrometric Analysis and Proteotyping of P. insidiosum
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Ethical Approval
References
- Mendoza, L.; Ajello, L.; McGinnis, M.R. Infection Caused by the Oomycetous Pathogen Pythium Insidiosum. J. Mycol. Med. 1996, 6, 151–164. [Google Scholar]
- Krajaejun, T.; Sathapatayavongs, B.; Pracharktam, R.; Nitiyanant, P.; Leelachaikul, P.; Wanachiwanawin, W.; Chaiprasert, A.; Assanasen, P.; Saipetch, M.; Mootsikapun, P.; et al. Clinical and Epidemiological Analyses of Human Pythiosis in Thailand. Clin. Infect. Dis. 2006, 43, 569–576. [Google Scholar] [CrossRef]
- Gaastra, W.; Lipman, L.J.; de Cock, A.W.; Exel, T.K.; Pegge, R.B.; Scheurwater, J.; Vilela, R.; Mendoza, L. Pythium Insidiosum: An Overview. Vet. Microbiol. 2010, 146. [Google Scholar] [CrossRef] [PubMed]
- Chitasombat, M.N.; Jongkhajornpong, P.; Lekhanont, K.; Krajaejun, T. Recent Update in Diagnosis and Treatment of Human Pythiosis. PeerJ 2020, 8, e8555. [Google Scholar] [CrossRef] [PubMed]
- Rujirawat, T.; Patumcharoenpol, P.; Lohnoo, T.; Yingyong, W.; Lerksuthirat, T.; Tangphatsornruang, S.; Suriyaphol, P.; Grenville-Briggs, L.J.; Garg, G.; Kittichotirat, W.; et al. Draft Genome Sequence of the Pathogenic Oomycete Pythium Insidiosum Strain Pi-S, Isolated from a Patient with Pythiosis. Genome Announc. 2015, 3, e00574-15. [Google Scholar] [CrossRef]
- Rujirawat, T.; Patumcharoenpol, P.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Payattikul, P.; Tangphatsornruang, S.; Suriyaphol, P.; Reamtong, O.; Garg, G.; et al. Probing the Phylogenomics and Putative Pathogenicity Genes of Pythium Insidiosum by Oomycete Genome Analyses. Sci. Rep. 2018, 8, 4135. [Google Scholar] [CrossRef] [PubMed]
- Kittichotirat, W.; Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Krajaejun, T. Draft Genome and Sequence Variant Data of the Oomycete Pythium Insidiosum Strain Pi45 from the Phylogenetically-Distinct Clade-III. Data Brief. 2017, 15, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Krajaejun, T.; Kittichotirat, W.; Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W. Data on Whole Genome Sequencing of the Oomycete Pythium Insidiosum Strain CBS 101555 from a Horse with Pythiosis in Brazil. BMC Res. Notes 2018, 11, 880. [Google Scholar] [CrossRef]
- Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Vanittanakom, N.; Kittichotirat, W.; Krajaejun, T. Draft Genome Sequences of the Oomycete Pythium Insidiosum Strain CBS 573.85 from a Horse with Pythiosis and Strain CR02 from the Environment. Data Brief. 2018, 16, 47–50. [Google Scholar] [CrossRef]
- Krajaejun, T.; Lerksuthirat, T.; Garg, G.; Lowhnoo, T.; Yingyong, W.; Khositnithikul, R.; Tangphatsornruang, S.; Suriyaphol, P.; Ranganathan, S.; Sullivan, T.D. Transcriptome Analysis Reveals Pathogenicity and Evolutionary History of the Pathogenic Oomycete Pythium Insidiosum. Fungal Biol. 2014, 118, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Krajaejun, T.; Khositnithikul, R.; Lerksuthirat, T.; Lowhnoo, T.; Rujirawat, T.; Petchthong, T.; Yingyong, W.; Suriyaphol, P.; Smittipat, N.; Juthayothin, T.; et al. Expressed Sequence Tags Reveal Genetic Diversity and Putative Virulence Factors of the Pathogenic Oomycete Pythium Insidiosum. Fungal Biol. 2011, 115, 683–696. [Google Scholar] [CrossRef]
- Krajaejun, T.; Kittichotirat, W.; Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W. Draft Genome Sequence of the Oomycete Pythium Destruens Strain ATCC 64221 from a Horse with Pythiosis in Australia. BMC Res. Notes 2020, 13, 329. [Google Scholar] [CrossRef] [PubMed]
- Jindayok, T.; Piromsontikorn, S.; Srimuang, S.; Khupulsup, K.; Krajaejun, T. Hemagglutination Test for Rapid Serodiagnosis of Human Pythiosis. Clin. Vaccine Immunol. 2009, 16, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Keeratijarut, A.; Karnsombut, P.; Aroonroch, R.; Srimuang, S.; Sangruchi, T.; Sansopha, L.; Mootsikapun, P.; Larbcharoensub, N.; Krajaejun, T. Evaluation of an In-House Immunoperoxidase Staining Assay for Histodiagnosis of Human Pythiosis. Southeast Asian J. Trop. Med. Public Health 2009, 40, 1298–1305. [Google Scholar] [PubMed]
- Keeratijarut, A.; Lohnoo, T.; Yingyong, W.; Sriwanichrak, K.; Krajaejun, T. A Peptide ELISA to Detect Antibodies against Pythium Insidiosum Based on Predicted Antigenic Determinants of Exo-1,3-β-Glucanase. Southeast Asian J. Trop. Med. Public Health 2013, 44, 672–680. [Google Scholar]
- Keeratijarut, A.; Lohnoo, T.; Yingyong, W.; Nampoon, U.; Lerksuthirat, T.; Onpaew, P.; Chongtrakool, P.; Krajaejun, T. PCR Amplification of a Putative Gene for Exo-1, 3-β-Glucanase to Identify the Pathogenic Oomycete Pythium Insidiosum. Asian Biomed. 2014, 8, 637–644. [Google Scholar] [CrossRef]
- Keeratijarut, A.; Lohnoo, T.; Yingyong, W.; Rujirawat, T.; Srichunrusami, C.; Onpeaw, P.; Chongtrakool, P.; Brandhorst, T.T.; Krajaejun, T. Detection of the Oomycete Pythium Insidiosum by Real-Time PCR Targeting the Gene Coding for Exo-1,3-β-Glucanase. J. Med. Microbiol. 2015, 64, 971–977. [Google Scholar] [CrossRef]
- Krajaejun, T.; Imkhieo, S.; Intaramat, A.; Ratanabanangkoon, K. Development of an Immunochromatographic Test for Rapid Serodiagnosis of Human Pythiosis. Clin. Vaccine Immunol. 2009, 16, 506–509. [Google Scholar] [CrossRef]
- Krajaejun, T.; Lohnoo, T.; Jittorntam, P.; Srimongkol, A.; Kumsang, Y.; Yingyong, W.; Rujirawat, T.; Reamtong, O.; Mangmee, S. Assessment of Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry for Identification and Biotyping of the Pathogenic Oomycete Pythium Insidiosum. Int. J. Infect. Dis. 2018, 77, 61–67. [Google Scholar] [CrossRef]
- Chareonsirisuthigul, T.; Khositnithikul, R.; Intaramat, A.; Inkomlue, R.; Sriwanichrak, K.; Piromsontikorn, S.; Kitiwanwanich, S.; Lowhnoo, T.; Yingyong, W.; Chaiprasert, A.; et al. Performance Comparison of Immunodiffusion, Enzyme-Linked Immunosorbent Assay, Immunochromatography and Hemagglutination for Serodiagnosis of Human Pythiosis. Diagn. Microbiol. Infect. Dis. 2013, 76, 42–45. [Google Scholar] [CrossRef]
- Lohnoo, T.; Jongruja, N.; Rujirawat, T.; Yingyon, W.; Lerksuthirat, T.; Nampoon, U.; Kumsang, Y.; Onpaew, P.; Chongtrakool, P.; Keeratijarut, A.; et al. Efficiency Comparison of Three Methods for Extracting Genomic DNA of the Pathogenic Oomycete Pythium Insidiosum. J. Med. Assoc. Thail. 2014, 97, 342–348. [Google Scholar]
- Intaramat, A.; Sornprachum, T.; Chantrathonkul, B.; Chaisuriya, P.; Lohnoo, T.; Yingyong, W.; Jongruja, N.; Kumsang, Y.; Sandee, A.; Chaiprasert, A.; et al. Protein A/G-Based Immunochromatographic Test for Serodiagnosis of Pythiosis in Human and Animal Subjects from Asia and Americas. Med. Mycol. 2016, 54, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Rujirawat, T.; Sridapan, T.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Sae-Chew, P.; Tonpitak, W.; Krajaejun, T. Single Nucleotide Polymorphism-Based Multiplex PCR for Identification and Genotyping of the Oomycete Pythium Insidiosum from Humans, Animals and the Environment. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2017, 54, 429–436. [Google Scholar] [CrossRef]
- Mani, R.; Vilela, R.; Kettler, N.; Chilvers, M.I.; Mendoza, L. Identification of Pythium Insidiosum Complex by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Med. Microbiol. 2019, 68, 574–584. [Google Scholar] [CrossRef]
- Htun, Z.M.; Rotchanapreeda, T.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Sae-Chew, P.; Payattikul, P.; Yurayart, C.; Limsivilai, O.; et al. Loop-Mediated Isothermal Amplification (LAMP) for Identification of Pythium Insidiosum. Int. J. Infect. Dis. 2020, 101, 149–159. [Google Scholar] [CrossRef]
- Lerksuthirat, T.; Sangcakul, A.; Lohnoo, T.; Yingyong, W.; Rujirawat, T.; Krajaejun, T. Evolution of the Sterol Biosynthetic Pathway of Pythium Insidiosum and Related Oomycetes Contributes to Antifungal Drug Resistance. Antimicrob. Agents Chemother. 2017, 61, e02352-16. [Google Scholar] [CrossRef]
- Yolanda, H.; Krajaejun, T. Review of Methods and Antimicrobial Agents for Susceptibility Testing against Pythium Insidiosum. Heliyon 2020, 6, e03737. [Google Scholar] [CrossRef] [PubMed]
- Sathapatayavongs, B.; Leelachaikul, P.; Prachaktam, R.; Atichartakarn, V.; Sriphojanart, S.; Trairatvorakul, P.; Jirasiritham, S.; Nontasut, S.; Eurvilaichit, C.; Flegel, T. Human Pythiosis Associated with Thalassemia Hemoglobinopathy Syndrome. J. Infect. Dis. 1989, 159, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Hasika, R.; Lalitha, P.; Radhakrishnan, N.; Rameshkumar, G.; Prajna, N.V.; Srinivasan, M. Pythium Keratitis in South India: Incidence, Clinical Profile, Management, and Treatment Recommendation. Indian J. Ophthalmol. 2019, 67, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Appavu, S.P.; Prajna, L.; Rajapandian, S.G.K. Genotyping and Phylogenetic Analysis of Pythium Insidiosum Causing Human Corneal Ulcer. Med. Mycol. 2020, 58, 211–218. [Google Scholar] [CrossRef]
- Agarwal, S.; Iyer, G.; Srinivasan, B.; Benurwar, S.; Agarwal, M.; Narayanan, N.; Lakshmipathy, M.; Radhika, N.; Rajagopal, R.; Krishnakumar, S.; et al. Clinical Profile, Risk Factors and Outcome of Medical, Surgical and Adjunct Interventions in Patients with Pythiuminsidiosum Keratitis. Br. J. Ophthalmol. 2019, 103, 296–300. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, C.E.P.; Ubiali, D.G.; Pescador, C.A.; Zanette, R.A.; Santurio, J.M.; Marques, L.C. Epidemiological Survey of Equine Pythiosis in the Brazilian Pantanal and Nearby Areas: Results of 76 Cases. J. Equine Vet. Sci. 2014, 34, 270–274. [Google Scholar] [CrossRef]
- Mendoza, L.; Hernandez, F.; Ajello, L. Life Cycle of the Human and Animal Oomycete Pathogen Pythium Insidiosum. J. Clin. Microbiol. 1993, 31, 2967–2973. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.I. Investigations into the Biology of Three “phycomycotic” Agents Pathogenic for Horses in Australia. Mycopathologia 1983, 81, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Supabandhu, J.; Fisher, M.C.; Mendoza, L.; Vanittanakom, N. Isolation and Identification of the Human Pathogen Pythium Insidiosum from Environmental Samples Collected in Thai Agricultural Areas. Med. Mycol. 2008, 46, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Presser, J.W.; Goss, E.M. Environmental Sampling Reveals that Pythium Insidiosum Is Ubiquitous and Genetically Diverse in North Central Florida. Med. Mycol. 2015, 53, 674–683. [Google Scholar] [CrossRef]
- Vanittanakom, N.; Szekely, J.; Khanthawong, S.; Sawutdeechaikul, P.; Vanittanakom, P.; Fisher, M.C. Molecular Detection of Pythium Insidiosum from Soil in Thai Agricultural Areas. Int. J. Med. Microbiol. 2014, 304, 321–326. [Google Scholar] [CrossRef]
- Zambrano, C.G.; Fonseca, A.O.S.; Valente, J.S.S.; Braga, C.Q.; Sallis, E.S.V.; Azevedo, M.I.; Weiblen, C.; Santurio, J.M.; Botton, S.A.; Pereira, D.I.B. Isolamento e Caracterização de Espécies de Pythium de Ambientes Aquáticos No Estado Do Rio Grande Do Sul e Avaliação Da Patogenicidade Em Modelo Experimental. Pesqui. Veterinária Bras. 2017, 37, 459–464. [Google Scholar] [CrossRef][Green Version]
- Jara, M.; Holcomb, K.; Wang, X.; Goss, E.M.; Machado, G. The Potential Distribution of Pythium Insidiosum in the Chincoteague National Wildlife Refuge, Virginia. Front. Vet. Sci. 2021, 8, 640339. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Orlando, FL, USA, 1990; pp. 315–322. [Google Scholar]
- Martin, F.N. Phylogenetic Relationships among Some Pythium Species Inferred from Sequence Analysis of the Mitochondrially Encoded Cytochrome Oxidase II Gene. Mycologia 2000, 92, 711–727. [Google Scholar] [CrossRef]
- Robideau, G.P.; de Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA Barcoding of Oomycetes with Cytochrome c Oxidase Subunit I and Internal Transcribed Spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Mahadevakumar, S.; Sridhar, K.R. Diagnosis of Pythium by Classical and Molecular Approaches. In Pythium; Rai, M., Abd-Elsalam, K.A., Ingle, A.P., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 200–224. ISBN 978-0-429-29640-6. [Google Scholar]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny. Fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate Likelihood-Ratio Test for Branches: A Fast, Accurate, and Powerful Alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Chevenet, F.; Brun, C.; Bañuls, A.-L.; Jacq, B.; Christen, R. TreeDyn: Towards Dynamic Graphics and Annotations for Analyses of Trees. BMC Bioinform. 2006, 7, 439. [Google Scholar] [CrossRef] [PubMed]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef]
- Sauer, S.; Freiwald, A.; Maier, T.; Kube, M.; Reinhardt, R.; Kostrzewa, M.; Geider, K. Classification and Identification of Bacteria by Mass Spectrometry and Computational Analysis. PLoS ONE 2008, 3, e2843. [Google Scholar] [CrossRef]
- Schurko, A.M.; Mendoza, L.; Lévesque, C.A.; Désaulniers, N.L.; de Cock, A.W.A.M.; Klassen, G.R. A Molecular Phylogeny of Pythium Insidiosum. Mycol. Res. 2003, 107, 537–544. [Google Scholar] [CrossRef]
- Chaiprasert, A.; Krajaejun, T.; Pannanusorn, S.; Prariyachatigul, C.; Wanachiwanawin, W.; Sathapatayavongs, B.; Juthayothin, T.; Smittipat, N.; Vanittanakom, N.; Chindamporn, A. Pythium Insidiosum Thai Isolates: Molecular Phylogenetic Analysis. Asian Biomed. 2009, 3, 623–633. [Google Scholar]
- Chindamporn, A.; Kammarnjessadakul, P.; Kesdangsakonwut, S.; Banlunara, W. A Case of Canine Cutaneous Pythiosis in Thailand. Access Microbiol. 2020, 2. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Province (Province ID; Number of P. insidiosum Isolates) | Sampling Locations | Type of Water Collection Site | Number of Sites | Collected Samples | Number of Culture-Positive Samples (Species; % of Collected Samples) | |||
|---|---|---|---|---|---|---|---|---|
| Code | GPS Coordinate | P. insidiosum | Other Pythium Species | Other Organisms | ||||
| Bangkok (BK; n = 7) | BK01 | 13.770966, 100.516366 | Pond (zoo) | 10 | 50 | 2 (4%) | 1 (Pythium sp.; 2%) | - |
| BK02 | 13.854501, 100.859078 | Pond (zoo) | 3 | 15 | - | - | - | |
| BK03 | 13.807507, 100.555620 | Pond (public park) | 8 | 40 | - | - | - | |
| BK04 | 13.929843, 100.568340 | Pond (public park) | 4 | 20 | 1 (5%) | - | 1 (unclassified; 5%) | |
| BK05 | 13.787263, 100.674502 | Pond (public park) | 3 | 15 | - | - | - | |
| BK06 | 13.731485, 100.541550 | Pond (public park) | 4 | 20 | - | - | - | |
| BK07 | 13.652241, 100.491406 | Pond (public park) | 4 | 20 | - | - | - | |
| BK08 | 13.770022, 100.494723 | Pond (public park) | 3 | 15 | - | - | - | |
| BK09 | 13.686055, 100.662055 | Pond (public park) | 7 | 35 | 4 (11%) | 6 (P. catenulatum; 17%), 1 (Pythium sp.; 3%) | - | |
| BK10 | 13.744389, 100.352753 | Pond (public park) | 2 | 10 | - | - | - | |
| Chonburi (CB; n = 0) | CB01 | 13.257211, 101.151760 | Pond, Irrigation channel | 3 | 15 | - | 3 (P. catenulatum; 20%) | 1 (unclassified; 7%) |
| Chachoengsao (CS; n = 6) | CS01 | 13.668917, 101.191042 | Pond | 4 | 20 | 1 (5%) | 1 (P. rhizo-oryzae; 5%), 1 (Pythium sp.; 5%) | - |
| CS02 | 13.652780, 101.161328 | Pond | 2 | 10 | - | 1 (P. rhizo-oryzae; 10%) | - | |
| CS03 | 13.606459, 101.232437 | Rice field | 2 | 10 | 5 (50%) | - | - | |
| Nakhon Pathom (NP; n = 0) | NP01 | 14.020776, 99.973345 | Pond | 12 | 60 | - | 6 (P. catenulatum; 10%) | 1 (Sclerotium hydrophilum; 2%), 1 (Mucor amphibiorum; 2%) |
| Kanchanaburi (KB; n = 11) | KB01 | 14.138575, 99.324756 | Pond | 5 | 25 | 1 (4%) | - | - |
| KB02 | 13.978799, 99.659559 | Pond | 1 | 5 | - | 1 (P. catenulatum; 20%) | 2 (unclassified; 40%) | |
| KB03 | 14.027293, 99.791957 | Pond, Rice field | 5 | 25 | 10 (40%) | 1 (Pythium sp.; 4%) | - | |
| Ratchaburi (RB; n = 1) | RB01 | 13.806338, 99.689354 | Pond | 6 | 30 | 1 (3%) | 1 (P. rhizo-oryzae; 3%) | - |
| RB02 | 13.571397, 99.774272 | Pond | 3 | 15 | - | 4 (P. rhizo-oryzae; 27%), 1 (P. inflatum; 7%) | 1 (Pezizomycetes sp.; 7%) | |
| RB03 | 13.514250, 99.715126 | Pond | 2 | 10 | - | 1 (P. rhizo-oryzae; 10%) | - | |
| RB04 | 13.507072, 99.840394 | Pond | 2 | 10 | - | - | - | |
| Trang (TG; n = 2) | TG01 | 07.534068, 99.618507 | Pond, Rice field | 5 | 25 | 2 (8%) | 1 (Pythium sp.; 4%) | - |
| Total (n = 27) | - | - | - | 100 | 500 | 27 (5.4%) | 30 (6.0%) | 7 (1.4%) |
| Province (P. insidiosum Isolates) | Sampling Locations | Water Source (Collection Site) | Strain ID | DNA Barcodes | Genotype (Clade) | MALDI-TOF MS | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Code | GPS Coordinate | rDNA | cox1 | cox2 | DNA Barcode | Multiplex PCR | Score | Proteotype | ||||||
| Accession | % Identity | Accession | % Identity | Accession | % Identity | |||||||||
| Bangkok (n = 7) | BK01 | 13.770966, 100.516366 | Pond#1 (zoo) | BKDZ01 | LC556017 | 99.9 | LC547937 | 94.3 | LC549516 | 100.0 | III | III | - | - |
| Pond#2 (zoo) | BKDZ02 | LC556018 | 98.0 | LC547938 | 94.1 | LC549517 | 100.0 | III | III | 2.200 | B | |||
| BK04 | 13.929843, 100.568340 | Pond#3 (park) | RT02 | LC556073 | 99.9 | LC547929 | 99.7 | LC549523 | 100.0 | II | II | - | - | |
| BK09 | 13.686055, 100.662055 | Pond#4 (park) | RM9-02 | LC556063 | 99.5 | LC547930 | 100.0 | LC549519 | 100.0 | II | II | - | - | |
| RM9-03 | LC556064 | 99.8 | LC547931 | 100.0 | LC549520 | 100.0 | II | II | - | - | ||||
| RM9-04 | LC556065 | 99.8 | LC547932 | 100.0 | LC549521 | 100.0 | II | II | - | - | ||||
| RM9-05 | LC556066 | 99.4 | LC547933 | 100.0 | LC549522 | 100.0 | II | II | - | - | ||||
| Chachoengsao (n = 6) | CS01 | 13.668917, 101.191042 | Pond#5 | CCS01 | LC556020 | 99.9 | LC547923 | 100.0 | LC549500 | 100.0 | II | II | - | - |
| CS03 | 13.606459, 101.232437 | Rice Field#1 | CCS03 | LC556022 | 99.8 | LC547924 | 100.0 | LC549501 | 100.0 | II | II | - | - | |
| CCS04 | LC556023 | 100.0 | LC547925 | 100.0 | LC549502 | 100.0 | II | II | - | - | ||||
| CCS05 | LC556024 | 100.0 | LC547926 | 100.0 | LC549503 | 100.0 | II | II | - | - | ||||
| Rice Field#2 | CCS07 | LC556025 | 99.9 | LC547927 | 100.0 | LC549504 | 100.0 | II | II | - | - | |||
| CCS08 | LC556026 | 99.9 | LC547928 | 100.0 | LC549505 | 100.0 | II | II | - | - | ||||
| Kanchanaburi (n = 11) | KB01 | 14.138575, 99.324756 | Pond#6 | KCB12 | LC556043 | 99.4 | - | - | - | - | - | II | - | - |
| KB03 | 14.027293, 99.791957 | Rice Field#3 | KCB01 | LC556032 | 99.8 | LC547939 | 94.4 | LC549506 | 100.0 | III | III | 2.490 | B | |
| KCB02 | LC556033 | 99.9 | LC547919 | 99.3 | LC549507 | 100.0 | II | II | 2.274 | A | ||||
| KCB03 | LC556034 | 99.7 | LC547940 | 94.3 | LC549508 | 100.0 | III | III | 2.151 | B | ||||
| KCB04 | LC556035 | 99.4 | LC547941 | 94.4 | LC549509 | 100.0 | III | III | 2.187 | B | ||||
| KCB05 | LC556036 | 99.9 | LC547920 | 99.3 | LC549510 | 100.0 | II | II | 2.149 | A | ||||
| KCB06 | LC556037 | 99.7 | LC547942 | 94.4 | LC549511 | 100.0 | III | III | - | - | ||||
| KCB07 | LC556038 | 100.0 | LC547921 | 99.3 | LC549512 | 100.0 | II | II | 2.196 | A | ||||
| KCB08 | LC556039 | 99.9 | LC547943 | 94.4 | LC549513 | 100.0 | III | III | 2.400 | B | ||||
| KCB09 | LC556040 | 99.8 | LC547944 | 94.4 | LC549514 | 100.0 | III | III | 2.187 | B | ||||
| KCB10 | LC556041 | 99.4 | LC547922 | 99.3 | LC549515 | 100.0 | II | II | 2.200 | A | ||||
| Ratchaburi (n = 1) | RB01 | 13.806338, 99.689354 | Pond#7 | RCB04 | LC556056 | 99.6 | LC547936 | 99.3 | LC549518 | 100.0 | II | II | - | - |
| Trang (n = 2) | TG01 | 07.534068, 99.618507 | Rice Field#4 | TRG02 | LC556075 | 99.9 | LC547934 | 100.0 | LC549524 | 100.0 | II | II | - | - |
| Pond#8 | TRG03 | LC556076 | 99.9 | LC547935 | 100.0 | LC549525 | 100.0 | II | II | - | - | |||
| Genotype | Strain ID | Reference ID | Source | Country | Proteotype | GenBank Accession | ||
|---|---|---|---|---|---|---|---|---|
| rDNA | cox1 | cox2 | ||||||
| Clade-I (n = 7) | Pi08 | CBS 580.85 | Horse | Costa Rica | A | AB898107 | LC553008 | LC553029 |
| Pi10 | ATCC 200269 | Human | USA | A | AB898108 | LC553003 | LC553024 | |
| Pi59 | EQ02 | Horse | Brazil | not done | LC550290 | LC553010 | LC553031 | |
| Pi60 | EQ04 | Horse | Brazil | not done | LC550291 | LC553011 | LC553032 | |
| Pi62 | EQ06 | Horse | Brazil | not done | LC550293 | LC553012 | LC553034 | |
| Pi07 | CBS 573.85 | Horse | Costa Rica | not done | AB971180 | LC553007 | LC553028 | |
| Pi74 | KU40017.3 | Dog | Thailand | not done | MT459311 | LC553013 | LC553035 | |
| Clade-II (n = 9) | Pi23 | MCC10 | Human | Thailand | not done | AB898115 | LC553019 | LC553038 |
| Pi25 | P19 | Human | Thailand | not done | AB898116 | LC553020 | LC553039 | |
| Pi29 | SIMI 1126-46 | Human | Thailand | not done | LC199882 | LC553016 | LC553040 | |
| Pi32 | P34 | Human | Thailand | not done | AB898121 | LC553017 | LC553041 | |
| Pi35 | Pi-S | Human | Thailand | A | AB898124 | BAS30577 | BAS30578 | |
| Pi36 | ATCC 64221 | Horse | Australia | A | LC199883 | LC553005 | LC553026 | |
| Pi37 | ATCC 28251 | Horse | Papua New Guinea | not done | LC199884 | LC553004 | LC553025 | |
| Pi40 | CBS 777.84 | Mosquito | India | A | LC199886 | LC553009 | LC553030 | |
| Pi53 | P39 | Horse | Thailand | not done | LC199889 | LC553018 | LC553042 | |
| Clade-III (n = 6) | Pi44 | MCC 17 | Human | Thailand | not done | AB971185 | LC553015 | LC553037 |
| Pi45 | MCC 13 | Human | Thailand | B | AB971186 | LC553014 | LC553036 | |
| Pi46 | SIMI 3306-44 | Human | Thailand | not done | AB971187 | LC553022 | LC553045 | |
| Pi47 | SIMI 2921-45 | Human | Thailand | B | AB971188 | LC553021 | LC553044 | |
| Pi49 | SIMI 7695-48 | Human | Thailand | B | AB898127 | LC553023 | LC553046 | |
| Pi50 | ATCC 90586 | Human | USA | B | AB971190 | LC553006 | LC553027 | |
| No | Sample Collection Locations (Province) | Water Collection Sites | Isolated Organisms | Strain ID | % Identity | DNA Barcode-Based Genotype (Clade) | Multiplex PCR Results (Clade) | GenBank Accessions | ||
|---|---|---|---|---|---|---|---|---|---|---|
| rDNA | cox1 | cox2 | ||||||||
| 1 | Bangkok | Zoo | Pythium insidiosum | BKDZ01 | 99.9 | III | Positive (III) | LC556017 | LC547937 | LC549516 |
| 2 | Bangkok | Zoo | Pythium insidiosum | BKDZ02 | 98.0 | III | Positive (III) | LC556018 | LC547938 | LC549517 |
| 3 | Bangkok | Zoo | Pythium species | BKDZ03 | 99.1 | - | Negative | LC556019 | - | - |
| 4 | Bangkok | Public park | Pythium species | RM9-01 | 95.9 | - | Negative | LC556062 | - | - |
| 5 | Bangkok | Public park | Pythium insidiosum | RM9-02 | 99.5 | II | Positive (II) | LC556063 | LC547930 | LC549519 |
| 6 | Bangkok | Public park | Pythium insidiosum | RM9-03 | 99.8 | II | Positive (II) | LC556064 | LC547931 | LC549520 |
| 7 | Bangkok | Public park | Pythium insidiosum | RM9-04 | 99.8 | II | Positive (II) | LC556065 | LC547932 | LC549521 |
| 8 | Bangkok | Public park | Pythium insidiosum | RM9-05 | 99.4 | II | Positive (II) | LC556066 | LC547933 | LC549522 |
| 9 | Bangkok | Public park | Pythium catenulatum | RM9-06 | 99.0 | - | Negative | LC556067 | LC553640 | LC553642 |
| 10 | Bangkok | Public park | Pythium catenulatum | RM9-07 | 99.9 | - | Negative | LC556068 | - | - |
| 11 | Bangkok | Public park | Pythium catenulatum | RM9-08 | 99.6 | - | Negative | LC556069 | - | - |
| 12 | Bangkok | Public park | Pythium catenulatum | RM9-09 | 99.7 | - | Negative | LC556070 | - | - |
| 13 | Bangkok | Public park | Pythium catenulatum | RM9-10 | 99.5 | - | Negative | LC556071 | - | - |
| 14 | Bangkok | Public park | Pythium catenulatum | RM9-11 | 98.6 | - | Negative | LC556072 | - | - |
| 15 | Bangkok | Public park | Pythium insidiosum | RT02 | 99.9 | II | Positive (II) | LC556073 | LC547929 | LC549523 |
| 16 | Bangkok | Public park | Unclassified | RT01 | - | - | Negative | - | - | - |
| 17 | Chachangsao | Pond | Pythium insidiosum | CCS01 | 99.9 | II | Positive (II) | LC556020 | LC547923 | LC549500 |
| 18 | Chachangsao | Pond | Pythium species | CCS02 | 99.5 | - | Negative | LC556021 | - | - |
| 19 | Chachangsao | Pond | Pythium insidiosum | CCS03 | 99.8 | II | Positive (II) | LC556022 | LC547924 | LC549501 |
| 20 | Chachangsao | Pond | Pythium insidiosum | CCS04 | 100.0 | II | Positive (II) | LC556023 | LC547925 | LC549502 |
| 21 | Chachangsao | Pond | Pythium insidiosum | CCS05 | 100.0 | II | Positive (II) | LC556024 | LC547926 | LC549503 |
| 22 | Chachangsao | Pond | Pythium insidiosum | CCS07 | 99.9 | II | Positive (II) | LC556025 | LC547927 | LC549504 |
| 23 | Chachangsao | Pond | Pythium insidiosum | CCS08 | 99.9 | II | Positive (II) | LC556026 | LC547928 | LC549505 |
| 24 | Chachangsao | Pond | Pythium rhizo-oryzae | CCS13 | 99.8 | - | Negative | LC556031 | - | - |
| 25 | Chonburi | Pond | Unclassified | CCS06 | - | - | Negative | - | - | - |
| 26 | Chonburi | Pond | Pythium rhizo-oryzae | CCS09 | 99.6 | - | Negative | LC556027 | - | - |
| 27 | Chonburi | Irrigation channel | Pythium catenulatum | CCS10 | 99.8 | - | Negative | LC556028 | - | - |
| 28 | Chonburi | Pond | Pythium catenulatum | CCS11 | 99.8 | - | Negative | LC556029 | - | - |
| 29 | Chonburi | Pond | Pythium catenulatum | CCS12 | 99.9 | - | Negative | LC556030 | - | - |
| 30 | Kanchanaburi | Rice field | Pythium insidiosum | KCB01 | 99.8 | III | Positive (III) | LC556032 | LC547939 | LC549506 |
| 31 | Kanchanaburi | Rice field | Pythium insidiosum | KCB02 | 99.9 | II | Positive (II) | LC556033 | LC547919 | LC549507 |
| 32 | Kanchanaburi | Rice field | Pythium insidiosum | KCB03 | 99.7 | III | Positive (III) | LC556034 | LC547940 | LC549508 |
| 33 | Kanchanaburi | Rice field | Pythium insidiosum | KCB04 | 99.4 | III | Positive (III) | LC556035 | LC547941 | LC549509 |
| 34 | Kanchanaburi | Rice field | Pythium insidiosum | KCB05 | 99.9 | II | Positive (II) | LC556036 | LC547920 | LC549510 |
| 35 | Kanchanaburi | Rice field | Pythium insidiosum | KCB06 | 99.7 | III | Positive (III) | LC556037 | LC547942 | LC549511 |
| 36 | Kanchanaburi | Rice field | Pythium insidiosum | KCB07 | 100.0 | II | Positive (II) | LC556038 | LC547921 | LC549512 |
| 37 | Kanchanaburi | Rice field | Pythium insidiosum | KCB08 | 99.9 | III | Positive (III) | LC556039 | LC547943 | LC549513 |
| 38 | Kanchanaburi | Rice field | Pythium insidiosum | KCB09 | 99.8 | III | Positive (III) | LC556040 | LC547944 | LC549514 |
| 39 | Kanchanaburi | Rice field | Pythium insidiosum | KCB10 | 99.4 | II | Positive (II) | LC556041 | LC547922 | LC549515 |
| 40 | Kanchanaburi | Rice field | Pythium species | KCB11 | 99.6 | - | Negative | LC556042 | - | - |
| 41 | Kanchanaburi | Rice field | Pythium insidiosum | KCB12 | 99.4 | - | Positive (II) | LC556043 | not done | not done |
| 42 | Kanchanaburi | Rice field | Unclassified | KCB13 | - | - | Negative | - | - | - |
| 43 | Kanchanaburi | Rice field | Unclassified | KCB14 | - | - | Negative | - | - | - |
| 44 | Kanchanaburi | Rice field | Pythium catenulatum | KCB15 | 99.0 | - | Negative | LC556044 | - | - |
| 45 | Nakhon Pathom | Pond | Pythium catenulatum | KPS01 | 99.8 | - | Negative | LC556045 | - | - |
| 46 | Nakhon Pathom | Pond | Pythium catenulatum | KPS02 | 99.6 | - | Negative | LC556046 | - | - |
| 47 | Nakhon Pathom | Pond | Mucor amphibiorum | KPS03 | 95.4 | - | Negative | LC556047 | - | - |
| 48 | Nakhon Pathom | Pond | Pythium catenulatum | KPS04 | 99.6 | - | Negative | LC556048 | - | - |
| 49 | Nakhon Pathom | Pond | Pythium catenulatum | KPS05 | 99.8 | - | Negative | LC556049 | - | - |
| 50 | Nakhon Pathom | Pond | Sclerotium hydrophilum | KPS06 | 90.5 | - | Negative | LC556050 | - | - |
| 51 | Nakhon Pathom | Pond | Pythium catenulatum | KPS07 | 99.8 | - | Negative | LC556051 | - | - |
| 52 | Nakhon Pathom | Pond | Pythium catenulatum | KPS08 | 99.8 | - | Negative | LC556052 | - | - |
| 53 | Ratchaburi | Pond | Pythium rhizo-oryzae | RCB01 | 99.9 | - | Negative | LC556053 | LC553639 | LC553641 |
| 54 | Ratchaburi | Pond | Pythium rhizo-oryzae | RCB02 | 99.6 | - | Negative | LC556054 | - | - |
| 55 | Ratchaburi | Pond | Pythium rhizo-oryzae | RCB03 | 99.9 | - | Negative | LC556055 | - | - |
| 56 | Ratchaburi | Pond | Pythium insidiosum | RCB04 | 99.6 | II | Positive (II) | LC556056 | LC547936 | LC549518 |
| 57 | Ratchaburi | Pond | Pythium inflatum | RCB05 | 100.0 | - | Negative | LC556057 | - | - |
| 58 | Ratchaburi | Pond | Pythium rhizo-oryzae | RCB06 | 99.4 | - | Negative | LC556058 | - | - |
| 59 | Ratchaburi | Pond | Pezizomycetes species | RCB07 | 99.8 | - | Negative | LC556059 | - | - |
| 60 | Ratchaburi | Pond | Pythium rhizo-oryzae | RCB08 | 99.8 | - | Negative | LC556060 | - | - |
| 61 | Ratchaburi | Pond | Pythium rhizo-oryzae | RCB09 | 99.6 | - | Negative | LC556061 | - | - |
| 62 | Trang | Rice field | Pythium species | TRG01 | 95.9 | - | Negative | LC556074 | - | - |
| 63 | Trang | Rice field | Pythium insidiosum | TRG02 | 99.9 | II | Positive (II) | LC556075 | LC547934 | LC549524 |
| 64 | Trang | Ponds | Pythium insidiosum | TRG03 | 99.9 | II | Positive (II) | LC556076 | LC547935 | LC549525 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mar Htun, Z.; Laikul, A.; Pathomsakulwong, W.; Yurayart, C.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Payattikul, P.; Sae-Chew, P.; Rujirawat, T.; et al. Identification and Biotyping of Pythium insidiosum Isolated from Urban and Rural Areas of Thailand by Multiplex PCR, DNA Barcode, and Proteomic Analyses. J. Fungi 2021, 7, 242. https://doi.org/10.3390/jof7040242
Mar Htun Z, Laikul A, Pathomsakulwong W, Yurayart C, Lohnoo T, Yingyong W, Kumsang Y, Payattikul P, Sae-Chew P, Rujirawat T, et al. Identification and Biotyping of Pythium insidiosum Isolated from Urban and Rural Areas of Thailand by Multiplex PCR, DNA Barcode, and Proteomic Analyses. Journal of Fungi. 2021; 7(4):242. https://doi.org/10.3390/jof7040242
Chicago/Turabian StyleMar Htun, Zin, Aree Laikul, Watcharapol Pathomsakulwong, Chompoonek Yurayart, Tassanee Lohnoo, Wanta Yingyong, Yothin Kumsang, Penpan Payattikul, Pattarana Sae-Chew, Thidarat Rujirawat, and et al. 2021. "Identification and Biotyping of Pythium insidiosum Isolated from Urban and Rural Areas of Thailand by Multiplex PCR, DNA Barcode, and Proteomic Analyses" Journal of Fungi 7, no. 4: 242. https://doi.org/10.3390/jof7040242
APA StyleMar Htun, Z., Laikul, A., Pathomsakulwong, W., Yurayart, C., Lohnoo, T., Yingyong, W., Kumsang, Y., Payattikul, P., Sae-Chew, P., Rujirawat, T., Jittorntam, P., Jaturapaktrarak, C., Chongtrakool, P., & Krajaejun, T. (2021). Identification and Biotyping of Pythium insidiosum Isolated from Urban and Rural Areas of Thailand by Multiplex PCR, DNA Barcode, and Proteomic Analyses. Journal of Fungi, 7(4), 242. https://doi.org/10.3390/jof7040242