
Enhancement of Lycopene Synthesis via Low-Frequency Alternating Magnetic Field in Brassica trispora
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Culture Condition
2.2. Main Reagents and Instruments
2.3. Magnetic Field Treatment
2.4. Analytical Method
2.4.1. Detection of Biomass and Lycopene Yield
2.4.2. Determination of Process Curve
2.4.3. Determination of Cell Permeability of Mycelium
2.4.4. Scanning Electron Microscope (SEM) Observation
2.4.5. Transcriptome Analysis
2.4.6. RT-qPCR
2.4.7. Data Analysis
3. Results
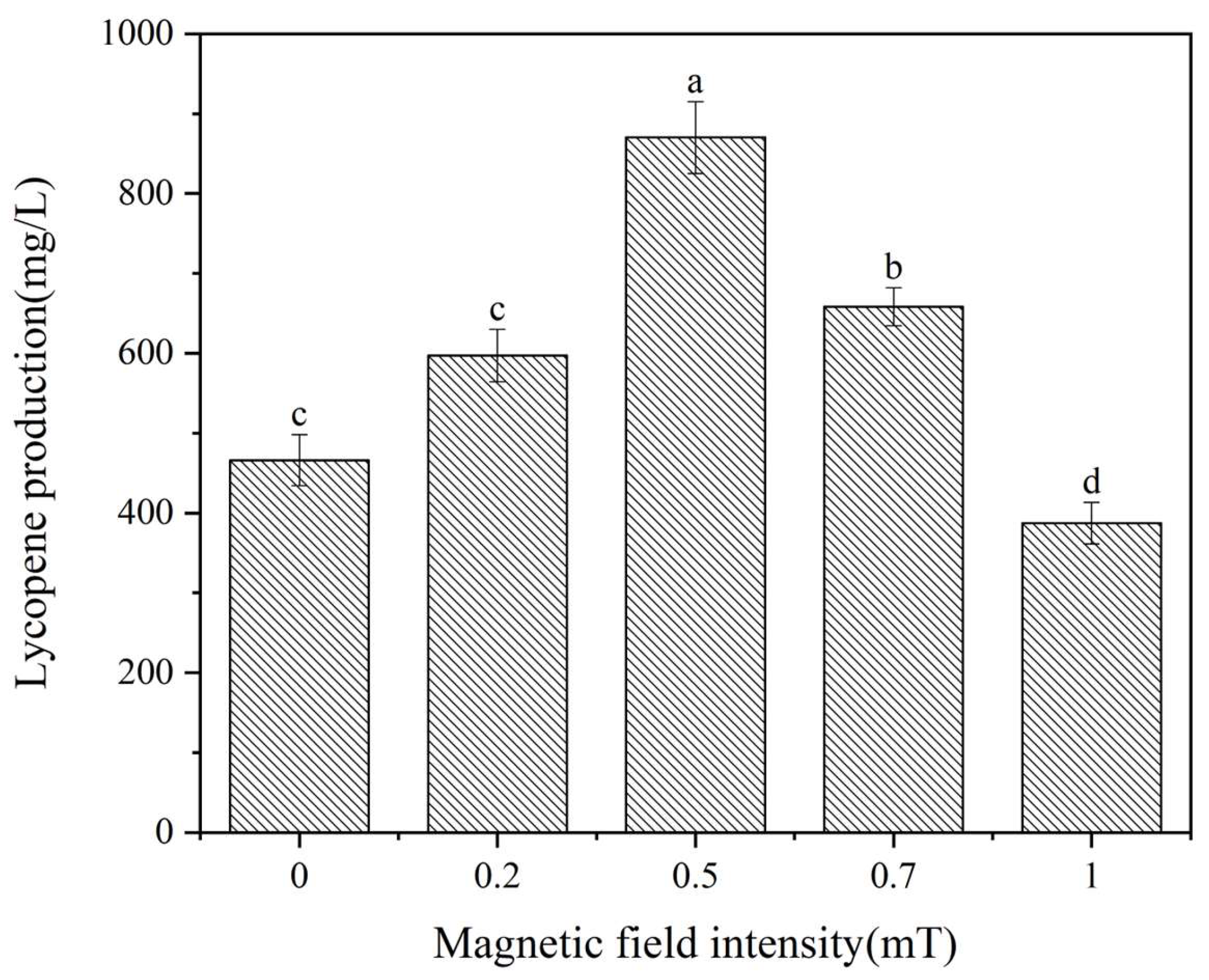
3.1. Magnetic Field Intensity
3.2. Magnetic Field Processing Time
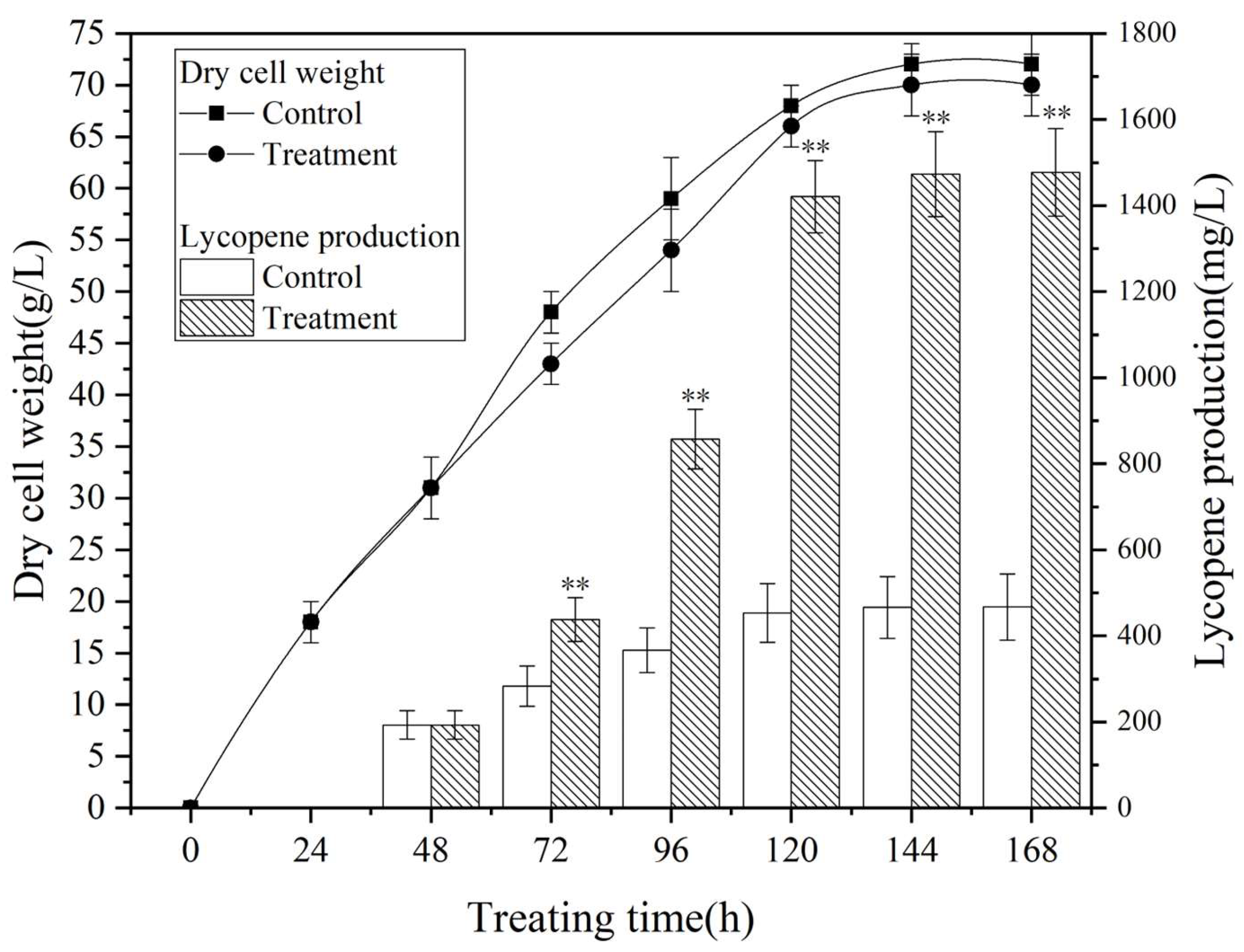
3.3. Process Metabolic Curve
3.4. Transcriptome Analysis of the Effects of Magnetic Field on Bacteriae
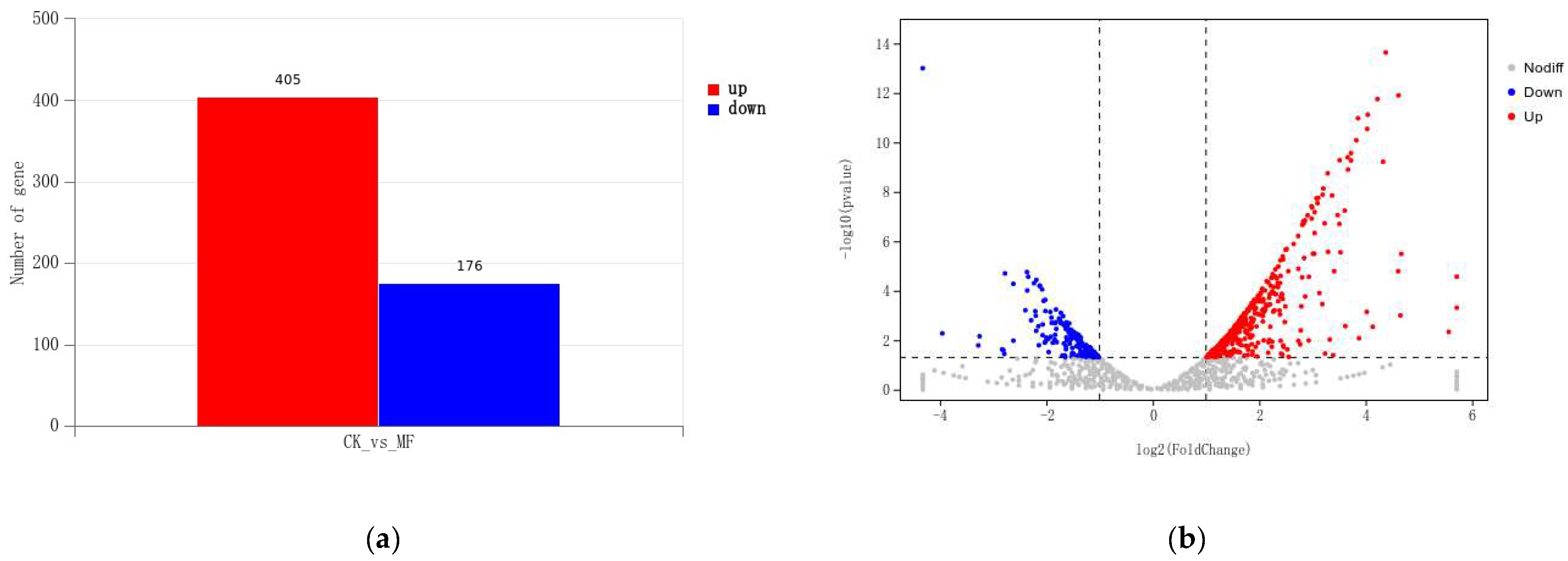
3.4.1. Differentially Expressed Genes
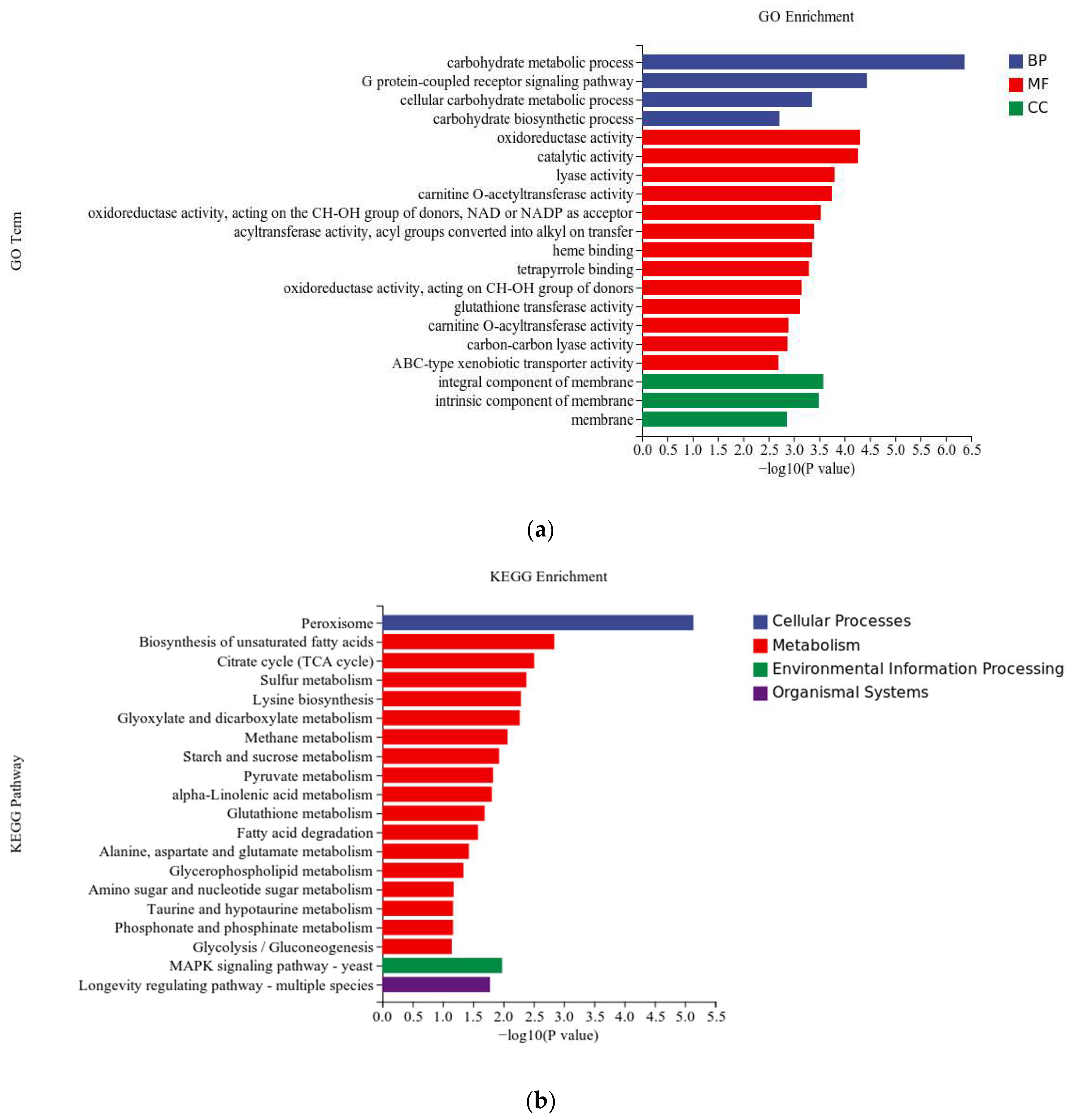
3.4.2. Differentially Expressed Genes
3.5. RT-qPCR Analysis
3.6. Mycelial Morphologye
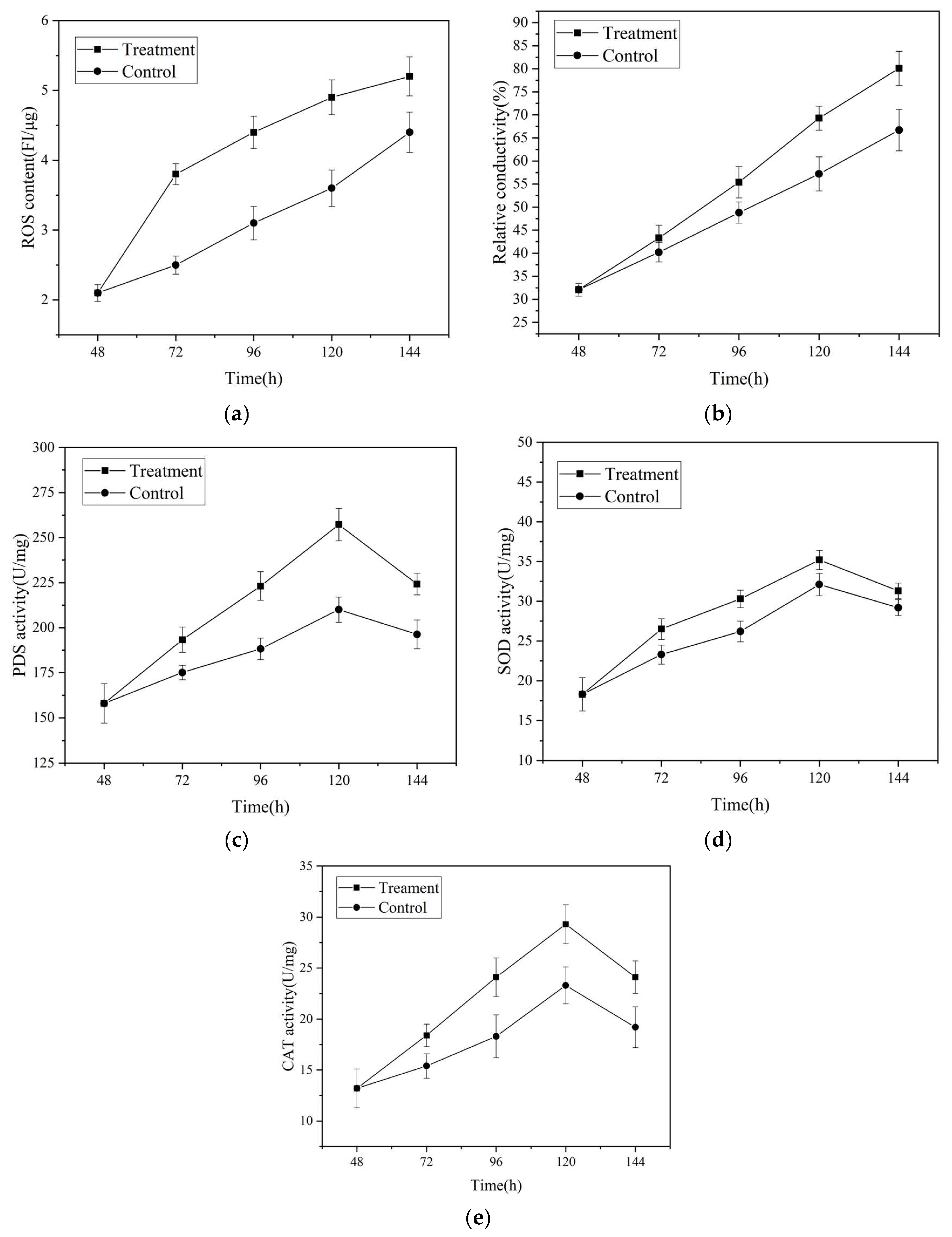
3.7. Effects of Magnetic Field Treatment on Intracellular Reactive Oxygen Species, Cell Membrane Permeabilit, y and Activities of Key enzymes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Yarema, K.; Xu, A. Impact of Static Magnetic Field (SMF) on Microorganisms, Plants and Animals; Springer: Singapore, 2017; pp. 133–172. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, D.; Xu, C.; Gao, M. Effect of low-frequency magnetic field on formation of pigments of Monascus purpureus. Eur. Food Res. Technol. 2014, 240, 577–582. [Google Scholar] [CrossRef]
- Makarov, I.O.K. Effect of Low-Frequency Pulsed Magnetic Field and Low-Level Laser Radiation on Oxidoreductase Activity and Growth of FungiActive Destructors of Polymer Materials. Microbiology 2019, 88, 72–78. [Google Scholar] [CrossRef]
- Aboneima, S.; El-Metwally, M.M. Effect of extremely low frequency magnetic field in growth, CMCase, electric conductivity and DNA of Aspergillus niger. Egypt. J. Phys. 2020, 49, 15–34. [Google Scholar] [CrossRef]
- Long, X.; Ye, J.; Zhao, D.; Zhang, S.J. Magnetogenetics: Remote non-invasive magnetic activation of neuronal activity with a magnetoreceptor. Sci. Bull. 2015, 60, 2107–2119. [Google Scholar] [CrossRef] [PubMed]
- Delbaere, S.M.; Lanssens, L.; Bernaerts, T.; Van Audenhove, J.; Hendrickx, M.E.; Grauwet, T.; Van Loey, A.M. Cell membrane permeabilization by pulsed electric field treatment impacts biochemical conversions and the volatile profile of broccoli stalks. LWT-Food Sci. Technol. 2023, 187, 115307. [Google Scholar] [CrossRef]
- Tagging, A.N.I.M. Re: Tomato and Lycopene and Multiple Health Outcomes: Umbrella Review. J. Urol. 2022, 207, 213. [Google Scholar]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Kaplan, D.N.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M. Lycopene: Food Sources, Biological Activities, and Human Health Benefits; Hindawi Limited: London, UK, 2021. [Google Scholar]
- Nigussie, A.; Tura, A.K.; Sisay, M.; Hagos, B.; Motbaynor, B. Anti-Cancer Effects of Lycopene in Animal Models of Hepatocellular Carcinoma: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2020, 11, 1306. [Google Scholar]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef]
- Crowe-White, K.M.; Phillips, T.A.; Ellis, A.C. Lycopene and cognitive function. J. Nutr. Sci. 2019, 8, e20. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K. Lycopene; chemistry, biosynthesis, metabolism and degradation under various abiotic parameters. J. Food Sci. Technol. 2015, 52, 41–53. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, H.; Zong, J.; Zhang, J.; Zhao, S.; Ma, H. Evaluating the effects of low-frequency alternating magnetic field thawing on oxidation, denaturation, and gelling properties of porcine myofibrillar proteins. Food Chem. 2024, 433, 137337. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-Q.; Yang, Q.; Wang, Y.-R.; Jiang, Y.-X.; Chen, H.-Q. Formation and structural characteristics of pea globulin amyloid-like fibrils pretreated with low-frequency magnetic field. Food Hydrocoll. 2024, 147, 109331. [Google Scholar] [CrossRef]
- Qiang, W.; Ling-Ran, F.; Luo, W.; Han-Guang, L.; Lin, W.; Ya, Z.; Xiao-Bin, Y. Mutation Breeding of Lycopene-Producing Strain Blakeslea Trispora by a Novel Atmospheric and Room Temperature Plasma (ARTP). Appl. Biochem. Biotechnol. 2014, 174, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Feng, L.R.; Luo, W.; Li, H.G.; Zhou, Y.; Yu, X.B. Effect of Inoculation Process on Lycopene Production by Blakeslea trispora in a Stirred-Tank Reactor. Appl. Biochem. Biotechnol. 2015, 175, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Müller, E.; Meffert, M.; Gerthsen, D. On the Progress of Scanning Transmission Electron Microscopy (STEM) Imaging in a Scanning Electron Microscope. Microsc. Microanal. 2018, 24, 99–106. [Google Scholar] [CrossRef]
- Schmidt, A.D.; Heinekamp, T.; Matuschek, M.; Liebmann, B.; Bollschweiler, C.; Brakhage, A.A. Analysis of mating-dependent transcription of Blakeslea trispora carotenoid biosynthesis genes carB and carRA by quantitative real-time PCR. Appl. Microbiol. Biotechnol. 2005, 67, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2013, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, D.; Zhu, L.; Guo, Y.; Zhao, Y.; Betchem, G.; Yolandani, Y.; Ma, H. Enhancing submerged fermentation of Antrodia camphorata by low-frequency alternating magnetic field. Innov. Food Sci. Emerg. Technol. 2023, 86, 103382. [Google Scholar] [CrossRef]
- Masood, S.; Saleem, I.; Smith, D.; Chu, W.K. Growth Pattern of Magnetic Field Treated Bacteria. Curr. Microbiol. 2018, 77, 194–203. [Google Scholar] [CrossRef]
- Falone, S.; Grossi, M.R.; Cinque, B.; D’Angelo, B.; Tettamanti, E.; Cimini, A.; Ilio, C.D.; Amicarelli, F. Fifty hertz extremely low-frequency electromagnetic field causes changes in redox and differentiative status in neuroblastoma cells. Int. J. Biochem. Cell Biol. 2007, 39, 2093–2106. [Google Scholar] [CrossRef]
- Mermall, V.; Post, P.L.; Mooseker, M.S. Unconventional Myosins in Cell Movement, Membrane Traffic, and Signal Transduction. Science 1998, 279, 527–533. [Google Scholar] [CrossRef]
- Klotz, J.L. CELL BIOLOGY SYMPOSIUM: Membrane trafficking and signal transduction. J. Anim. Sci. 2017, 95, 2183–2184. [Google Scholar] [CrossRef] [PubMed]
- Zablotskii, V.; Polyakova, T.; Dejneka, A. Modulation of the Cell Membrane Potential and Intracellular Protein Transport by High Magnetic Fields. Bioelectromagnetics 2020, 42, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Noda, Y.; Eckert, V.; Traber, M.G.; Packer, L. The phorbol 12-myristate 13- acetate (PMA)-induced oxidative burst in rat peritoneal neutrophils is increased by a 0.1 mT (60 Hz) magnetic field. FEBS Lett. 1995, 376, 164–166. [Google Scholar] [CrossRef] [PubMed]
- Dansen, T.B.; Wirtz, K.W.A. The Peroxisome in Oxidative Stress. IUBMB Life 2001, 51, 223–230. [Google Scholar] [CrossRef]
- Bonekamp, N.A.; Vlkl, A.; Fahimi, H.D.; Schrader, M. Reactive oxygen species and peroxisomes: Struggling for balance. Biofactors 2009, 35, 346–355. [Google Scholar] [CrossRef]
- Nanou, K.; Roukas, T.; Papadakis, E. Oxidative stress and morphological changes in Blakeslea trispora induced by enhanced aeration during carotene production in a bubble column reactor. Biochem. Eng. J. 2011, 54, 172–177. [Google Scholar] [CrossRef]
- Wang, H.B.; Luo, J.; Huang, X.Y.; Lu, M.B.; Yu, L.J. Oxidative stress response of Blakeslea trispora induced by HO during β-carotene biosynthesis. J. Ind. Microbiol. Biotechnol. 2014, 41, 555–561. [Google Scholar] [CrossRef]
- Qin, S.; Yin, H.; Yang, C.; Xie, C. A magnetic protein biocompass. Nat. Mater. 2015, 15, 217–226. [Google Scholar] [CrossRef]
- Zhang, Y.; Navarro, E.; Cánovas-Márquez, J.T.; Almagro, L.; Chen, H.; Chen, Y.Q.; Zhang, H.; Torres-Martínez, S.; Chen, W.; Garre, V. A new regulatory mechanism controlling carotenogenesis in the fungus Mucor circinelloides as a target to generate β-carotene over-producing strains by genetic engineering. Microb. Cell Factories 2016, 15, 99. [Google Scholar] [CrossRef]
- Guo, L.N.; Li, X.Y.; Zhang, X.Y.; Ma, H.L. Effect of low-intensity magnetic field on the growth and metabolite of Grifola frondosa in submerged fermentation and its possible mechanisms. Food Res. Int. 2022, 159, 111537. [Google Scholar] [CrossRef] [PubMed]
- Voychuk, S.I.; Gromozova, E.N.; Lytvyn, P.M.; Podgorsky, V.S. Changes of Surface Properties of Yeast Cell Wall Under Exposure of Electromagnetic Field (40.68 MHz) and Action of Nystatin. Environmentalist 2005, 25, 139–144. [Google Scholar] [CrossRef]
- Weaver, J.C. Electroporation: A general phenomenon for manipulating cells and tissues. J. Cell. Biochem. 1993, 51, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Quiles-Rosillo, M.D.; Ruiz-Vázquez, R.M.; Torres-Martínez, S.; Garre, V. Light induction of the carotenoid biosynthesis pathway in Blakeslea trispora. Fungal Genet. Biol. 2005, 42, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Liu, Y.; Zhang, J.; Li, L.; Gao, M. A low-frequency magnetic Field regulates Monascus pigments synthesis via reactive oxygen species in M. purpureus. Process Biochem. 2019, 86, 16–24. [Google Scholar] [CrossRef]
- Ikehara, T.; Yamaguchi, H.; Hosokawa, K.; Miyamoto, H.; Aizawa, K. Effects of ELF magnetic field on membrane protein structure of living HeLa cells studied by Fourier transform infrared spectroscopy. Bioelectromagnetics 2010, 24, 457–464. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Code | Forward Primer Sequence (5′3′) | Reverse Primer Sequence (5′3′) |
|---|---|---|---|
| carRA | gene_8655 | CATCTCGTCGTTGGTTCA | AAGCATAGGCAATAACACAAG |
| carB | gene_5071 | GGCACAGATATAACTTGA | TTATTCTTATTGGCTTCCT |
| sod | gene_5118 | ATCACTACAATCCTACTG | ACCATACTTCTTCCAATA |
| cat | gene_6144 | CTATGCTACCAGAGATATG | CCAGACCTTAGTTACATC |
| iscA | gene_10317 | CTGCTGCCAACTCGTTAA | CTGCTGGTGTCAGTGTAAG |
| elo3 | gene_7519 | TGGTCATCAAGAAGAAGA | GTCAAGTTCAGGATAATAGG |
| tef1 | GGTAAGTCTACCACCACTGGTCACT | CAAGAGGAGGGTAGTCAGTGTAAGC |
| Exposure Period (d) | Exposure Time (h) | Dry Cell Weight (g/L) | Lycopene Yield (mg/g) |
|---|---|---|---|
| Control | 0 | 63.3 ± 3.40 | 7.36 ± 0.38 |
| 1 | 4 | 60.4 ± 4.30 | 6.81 ± 0.31 e,* |
| 12 | 58.3 ± 3.50 | 8.02 ± 0.21 b | |
| 24 | 56.1 ± 3.80 | 7.81 ± 0.17 c | |
| 48 | 57.2 ± 4.20 | 8.60 ± 0.17 a | |
| 72 | 58.3 ± 5.30 | 7.23 ± 0.26 d | |
| 2 | 4 | 59.3 ± 2.70 | 9.80 ± 0.23 e |
| 12 | 58.1 ± 3.40 | 12.45 ± 0.35 d | |
| 24 | 56.3 ± 4.50 | 13.50 ± 0.32 c | |
| 48 | 58.1 ± 4.60 | 25.36 ± 0.42 a | |
| 72 | 60.2 ± 3.20 | 14.82 ± 0.25 b | |
| 3 | 4 | 59.3 ± 2.80 | 10.09 ± 0.19 e |
| 24 | 58.5 ± 5.20 | 11.84 ± 0.16 c | |
| 48 | 60.2 ± 3.20 | 15.15 ± 0.27 a | |
| 72 | 57.1 ± 4.60 | 12.98 ± 0.15 b | |
| 12 | 61.2 ± 2.40 | 10.99 ± 0.22 d |
| Name | Gene ID | Description | Fold Change of RNASeq | 2ΔΔCtof RT-qPCR |
|---|---|---|---|---|
| carRA | gene_8655 | Phytoene synthase | 1.12 | 2.15 |
| carB | gene_5071 | Phytoene desaturase | 2.23 | 1.47 |
| sod | gene_5118 | Superoxide dismutase | 2.42 | 2.15 |
| cat | gene_6144 | catalase | 8.18 | 7.80 |
| iscA | gene_10317 | Ironsulfur cluster assembly protein | 1.27 | 1.08 |
| elo3 | gene_7519 | ELO family | 4.26 | 3.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Hou, J.; Wang, D.; Yang, M.; Liu, J. Enhancement of Lycopene Synthesis via Low-Frequency Alternating Magnetic Field in Brassica trispora. Fermentation 2024, 10, 69. https://doi.org/10.3390/fermentation10010069
Wang H, Hou J, Wang D, Yang M, Liu J. Enhancement of Lycopene Synthesis via Low-Frequency Alternating Magnetic Field in Brassica trispora. Fermentation. 2024; 10(1):69. https://doi.org/10.3390/fermentation10010069
Chicago/Turabian StyleWang, Hong, Jiayang Hou, Dongxu Wang, Maohua Yang, and Jinlong Liu. 2024. "Enhancement of Lycopene Synthesis via Low-Frequency Alternating Magnetic Field in Brassica trispora" Fermentation 10, no. 1: 69. https://doi.org/10.3390/fermentation10010069
APA StyleWang, H., Hou, J., Wang, D., Yang, M., & Liu, J. (2024). Enhancement of Lycopene Synthesis via Low-Frequency Alternating Magnetic Field in Brassica trispora. Fermentation, 10(1), 69. https://doi.org/10.3390/fermentation10010069