
Utilization of the Dicarbonyl Compounds 3-Deoxyglucosone and 3-Deoxymaltosone during Beer Fermentation by Saccharomyces Yeasts


Abstract
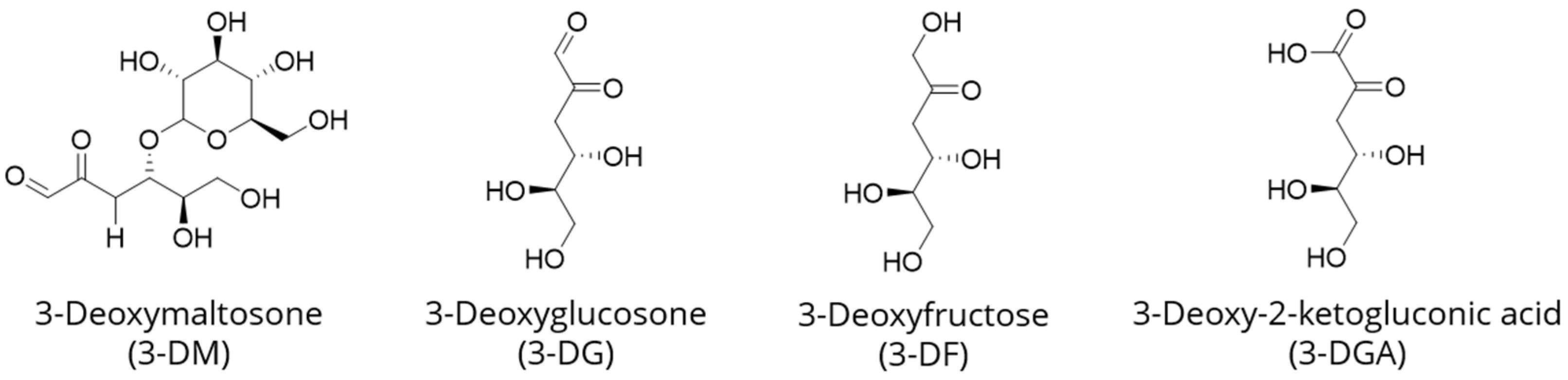
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Yeast Strains and Cultivation Conditions
2.3. Incubation Experiments in Pilsner Wort—Stability of 3-DG in the Presence of Saccharomyces Strains
2.4. Cell Disruption for Analysis of Internalized 3-DG and Yeast Metabolites
2.5. Dead/Live-Cell Experiments
2.6. Growth Curves of Saccharomyces Yeasts with 3-DG in Pilsner Wort
2.7. Fermentation Experiments in Different Beer Worts—Degradation and Metabolization of 3-DG and 3-DM by Saccharomyces Strains
2.8. Analysis of 3-DG and 3-DM
2.9. Gas Chromatography with Mass-Spectrometric Detection (GC-MS)
3. Results and Discussion
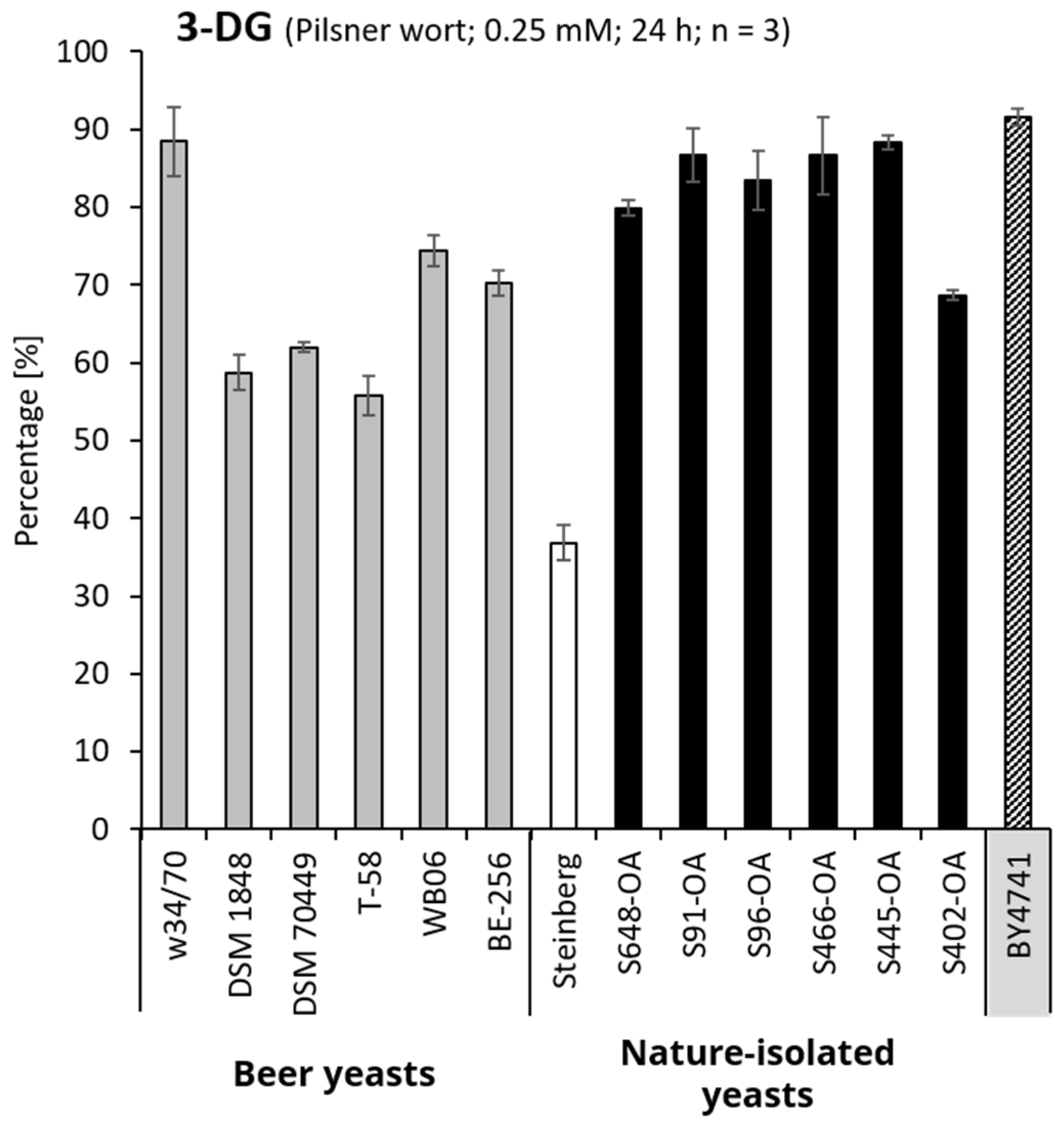
3.1. Utilization of 3-DG by Saccharomyces Strains
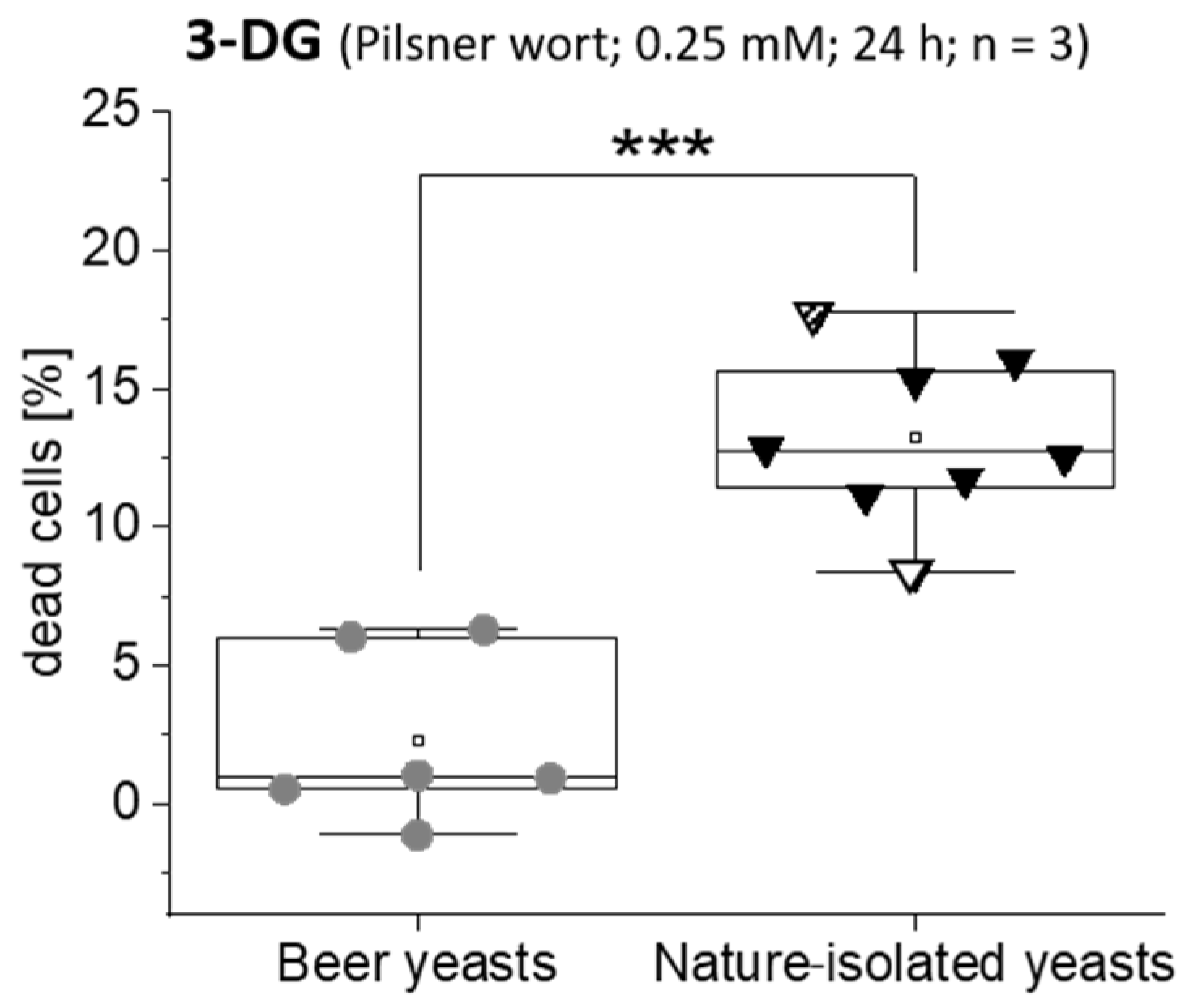
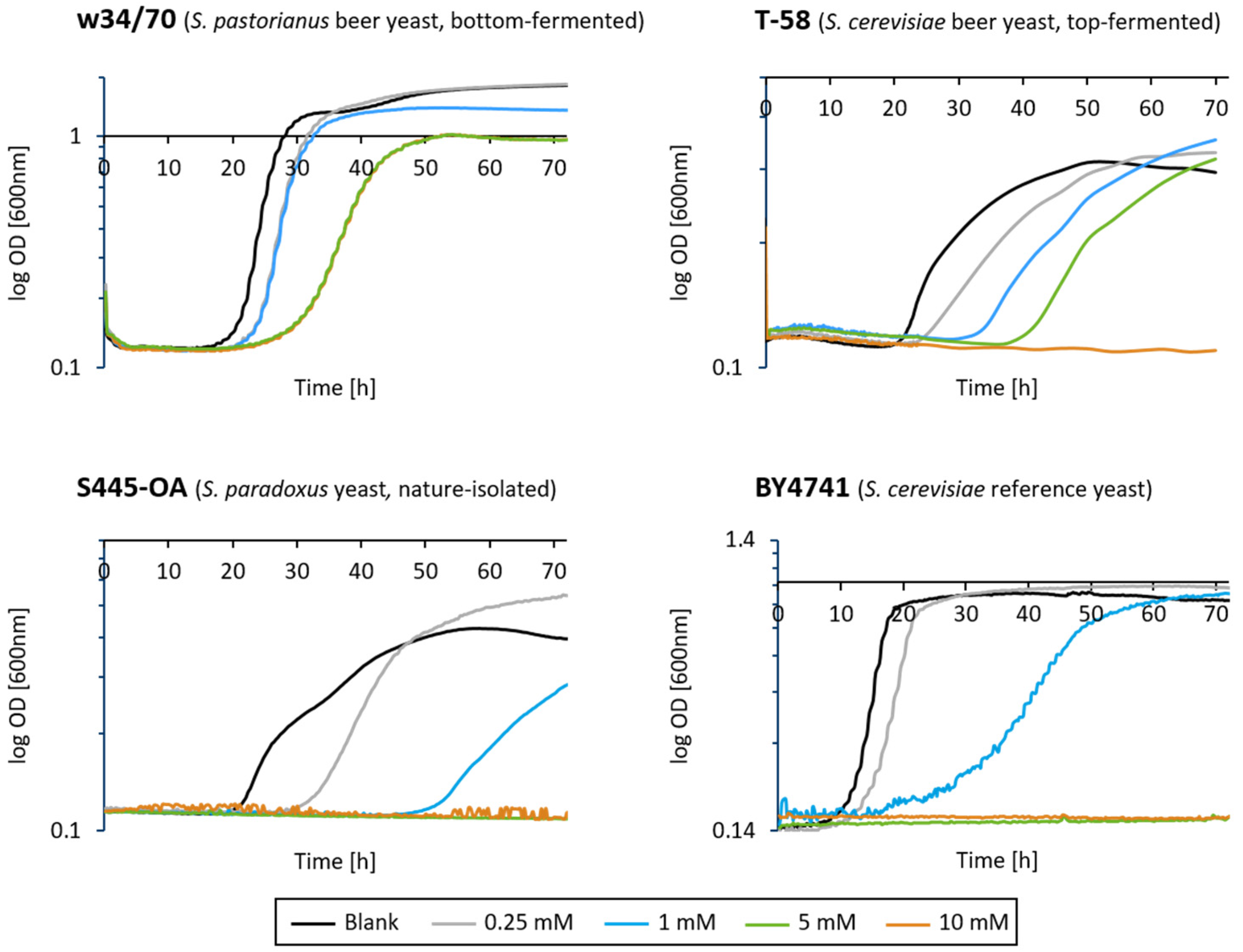
3.2. Influence of 3-DG on the Activity and Growth of Saccharomyces Yeasts
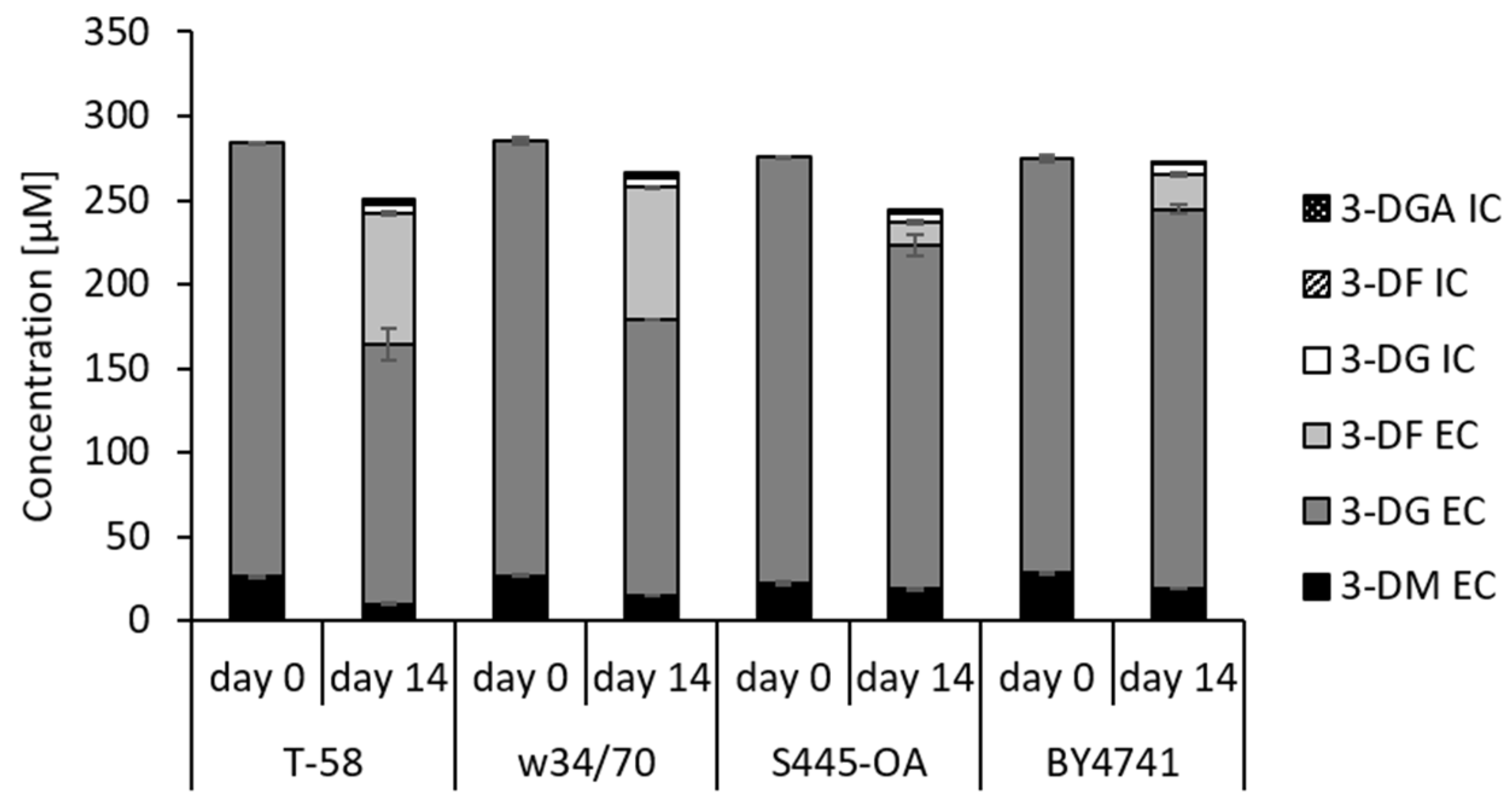
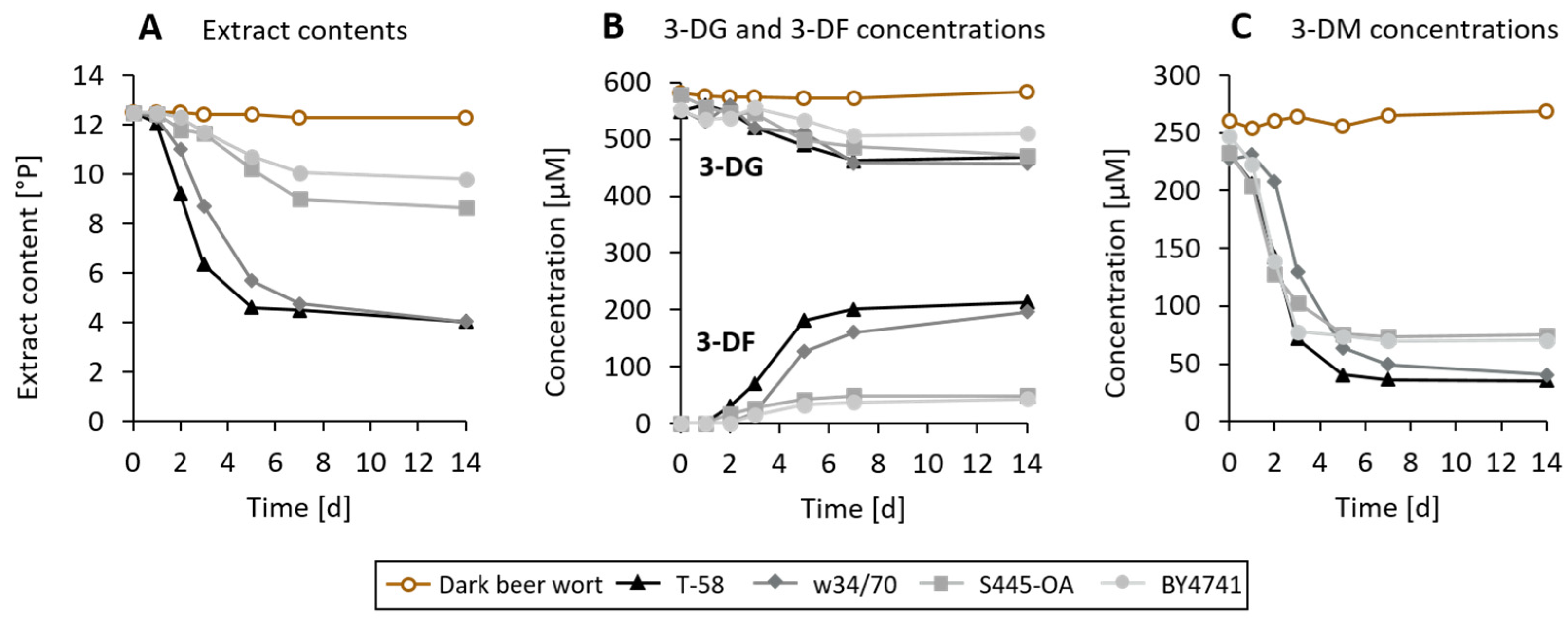
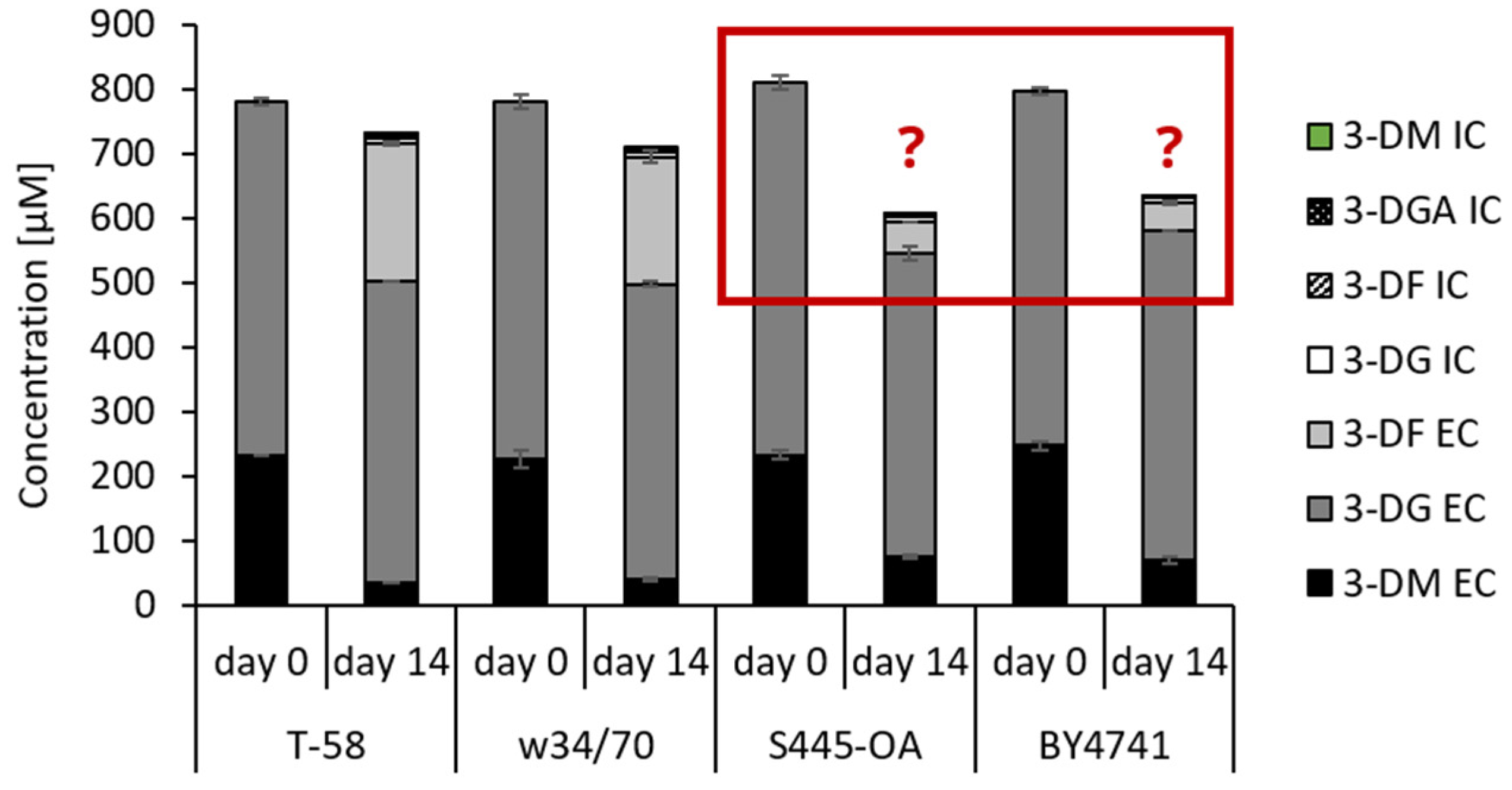
3.3. Degradation and Metabolization of 3-DG and 3-DM during Wort Fermentation by Saccharomyces Yeasts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hellwig, M.; Henle, T. Baking, Ageing, Diabetes: A Short History of the Maillard Reaction. Angew. Chem.-Int. Ed. 2014, 53, 10316–10329. [Google Scholar] [CrossRef]
- Hellwig, M.; Gensberger-Reigl, S.; Henle, T.; Pischetsrieder, M. Food-derived 1,2-dicarbonyl compounds and their role in diseases. Sem. Cancer Biol. 2018, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Turk, Z. Glycotoxines, Carbonyl Stress and Relevance to Diabetes and Its Complications. Physiol. Res 2010, 59, 147–156. [Google Scholar] [CrossRef]
- Degen, J.; Beyer, H.; Heymann, B.; Hellwig, M.; Henle, T. Dietary Influence on Urinary Excretion of 3-Deoxyglucosone and Its Metabolite 3-Deoxyfructose. J. Agric. Food Chem. 2014, 62, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Witte, S.; Henle, T. Free and Protein-Bound Maillard Reaction Products in Beer: Method Development and a Survey of Different Beer Types. J. Agric. Food Chem. 2016, 64, 7234–7243. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Herrera, J.C.; Scherer, E.; Ju-Nam, Y.; Rübsam, H.; Madrid, J.; Zufall, C.; Rangel-Aldao, R. Formation of α-Dicarbonyl Compounds in Beer during Storage of Pilsner. J. Agric. Food Chem. 2008, 56, 4134–4144. [Google Scholar] [CrossRef]
- Pires, E.J.; Teixeira, J.A.; Brányik, T. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef]
- Hellwig, M.; Nobis, A.; Witte, S.; Henle, T. Occurrence of (Z)-3,4-Dideoxyglucoson-3-ene in Different Types of Beer and Malt Beer as a Result of 3-Deoxyhexosone Interconversion. J. Agric. Food Chem. 2016, 64, 2746–2753. [Google Scholar] [CrossRef] [PubMed]
- Nobis, A.; Röhrig, A.; Hellwig, M.; Henle, T.; Becker, T.; Gastl, M. Formation of 3-deoxyglucosone in the malting process. Food Chem. 2019, 290, 187–195. [Google Scholar] [CrossRef]
- Degen, J.; Hellwig, M.; Henle, T. 1,2-Dicarbonyl Compounds in Commonly Consumed Foods. J. Agric. Food Chem. 2012, 60, 7071–7079. [Google Scholar] [CrossRef]
- Rakete, S.; Klaus, A.; Glomb, M.A. Investigations on the Maillard Reaction of Dextrins during Aging of Pilsner Type Beer. J. Agric. Food Chem. 2014, 62, 9876–9884. [Google Scholar] [CrossRef] [PubMed]
- Nobis, A.; Kwasnicki, M.; Lehnhardt, F.; Hellwig, M.; Henle, T.; Becker, T.; Gastl, M. A Comprehensive Evaluation of Flavor Instability of Beer (Part 2): The Influence of De Novo Formation of Aging Aldehydes. Foods 2021, 10, 2668. [Google Scholar] [CrossRef] [PubMed]
- Piddocke, M.P.; Kreisz, S.; Heldt-Hansen, H.P.; Nielsen, K.F.; Olsson, L. Physiological characterization of brewer’s yeast in high-gravity beer fermentations with glucose or maltose syrups as adjuncts. Appl. Microbiol. Biotechnol. 2009, 84, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Henle, T. Maillard Reaction Products in Different Types of Brewing Malt. J. Agric. Food Chem. 2020, 68, 14274–14285. [Google Scholar] [CrossRef]
- Van Bergen, B.; Cyr, N.; Strasser, R.; Blanchette, M.; Sheppard, J.D.; Jardim, A. α,β-Dicarbonyl reduction is mediated by the Saccharomyces Old Yellow Enzyme. FEMS Yeast Res. 2016, 16, fow059. [Google Scholar] [CrossRef] [PubMed]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Börner, M.; Beer, F.; Van Pée, K.H.; Henle, T. Transformation of Free and Dipeptide-Bound Glycated Amino Acids by Two Strains of Saccharomyces cerevisiae. ChemBioChem. 2017, 18, 66–275. [Google Scholar] [CrossRef]
- Hellwig, M.; Börner, M.; Henle, T. Reduction of 5-Hydroxymethylfurfural and 1,2-Dicarbonyl Compounds by Saccharomyces cerevisiae in Model Systems and Beer. J. Agric. Food Chem. 2021, 69, 12807–12817. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Röder, A.; Modig, T.; Laadan, B.; Lidén, G.; Gorwa-Grauslund, M.F. NADH- vs. NADPH-coupled reduction of 5-hydroxymethyl furfural (HMF) and its implications on product distribution in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2008, 78, 939–945. [Google Scholar] [CrossRef]
- Kertsch, A.; Brysch-Herzberg, M.; Hellwig, M.; Henle, T. Metabolization of the glycation compounds 3-deoxyglucosone and 5-hydroxymethylfurfural by Saccharomyces yeasts. Eur. Food Res. Technol. 2023, 249, 103–118. [Google Scholar] [CrossRef]
- Henle, T.; Bachmann, A. A Synthesis of pyrraline reference material. Z. Lebensm. Unters. Forsch. 1996, 202, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Mutz, Y.S.; Rosario, D.K.A.; Conte-Junior, C.A. Insights into chemical and sensorial aspects to understand and manage beer aging using chemometrics. Compr. Revi. Food Sci. Food Saf. 2020, 19, 3774–3801. [Google Scholar] [CrossRef] [PubMed]
- Rückriemen, J.; Hellwig, A.; Schultes, S.; Hellwig, M.; Hahne, F.; Henle, T. Studies on the influence of dietary 3-deoxyglucosone on the urinary excretion of 2-keto-3-deoxygluconic acid. Eur. Food Res. Technol. 2018, 244, 1389–1396. [Google Scholar] [CrossRef]
- Brysch-Herzberg, M.; Seidel, M. Distribution patterns of Saccharomyces species in cultural landscapes of Germany. FEMS Yeast Res. 2017, 17, fox033. [Google Scholar] [CrossRef] [PubMed]
- Thierig, M.; Raupbach, J.; Wolf, D.; Mascher, T.; Subramanian, K.; Henle, T. 3-Phenyllactic Acid and Polyphenols Are Substances Enhancing the Antibacterial Effect of Methylglyoxal in Manuka Honey. Foods 2023, 12, 1098. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Degen, J.; Henle, T. 3-Deoxygalactosone, a “new” 1,2-dicarbonyl compound in milk products. J. Agric. Food Chem. 2010, 58, 10752–10760. [Google Scholar] [CrossRef] [PubMed]
- Lodolo, E.J.; Kock, J.L.F.; Axcell, B.C.; Brooks, M. The yeast Saccharomyces cerevisiae—The main character in beer brewing. FEMS Yeast Res. 2008, 8, 1018–1036. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.M. Yeast Physiology and Biotechnology; John Wiley & Sons Ltd.: Chichester, UK, 1998. [Google Scholar]
- Stewart, G. Saccharomyces species in the Production of Beer. Beverages 2016, 2, 34. [Google Scholar] [CrossRef]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Lehrbuch der Lebensmittelchemie, 6th ed.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Niwa, T. 3-Deoxyglucosone: Metabolism, analysis, biological activity, and clinical implication. J. Chromatogr. B 1999, 731, 23–36. [Google Scholar] [CrossRef]
- Barnett, J.A.; Entian, K.D. A history of research on yeasts 9: Regulation of sugar metabolism. Yeast 2005, 22, 835–894. [Google Scholar] [CrossRef]
- Nobis, A.; Wendl, S.; Becker, T.; Gastl, M. Formation and degradation of 3-deoxyglucosone as a key intermediate for ageing indicators during wort boiling. J. Inst. Brew. 2021, 127, 358–366. [Google Scholar] [CrossRef]
- Legras, J.L.; Galeote, V.; Bigey, F.; Camarasa, C.; Marsit, S.; Nidelet, T.; Sanchez, I.; Couloux, A.; Guy, J.; Franco-Duarte, R. Adaptation of S. cerevisiae to Fermented Food Environments Reveals Remarkable Genome Plasticity and the Footprints of Domestication. Mol. Biol. Evol. 2018, 35, 1712–1727. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Time Frame | Quantifier Ion | Dwell Time | Qualifier Ion | Dwell Time |
|---|---|---|---|---|---|
| min | m/z * | ms | m/z * | ms | |
| 3-DF 1 | 20.60–22.50 | 214 | 40 | 524 | 70 |
| 3-DGA | 348 | 40 | 538 | 40 | |
| [13C]-3-DGA | 352 | 25 | 544 | 40 | |
| 3-DG | 22.85–31.00 | 347 | 60 | 537 | 60 |
| [13C]-3-DG | 351 | 45 | 543 | 45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kertsch, A.-L.; Brysch-Herzberg, M.; Ostermann, K.; Henle, T. Utilization of the Dicarbonyl Compounds 3-Deoxyglucosone and 3-Deoxymaltosone during Beer Fermentation by Saccharomyces Yeasts. Fermentation 2024, 10, 281. https://doi.org/10.3390/fermentation10060281
Kertsch A-L, Brysch-Herzberg M, Ostermann K, Henle T. Utilization of the Dicarbonyl Compounds 3-Deoxyglucosone and 3-Deoxymaltosone during Beer Fermentation by Saccharomyces Yeasts. Fermentation. 2024; 10(6):281. https://doi.org/10.3390/fermentation10060281
Chicago/Turabian StyleKertsch, Anna-Lena, Michael Brysch-Herzberg, Kai Ostermann, and Thomas Henle. 2024. "Utilization of the Dicarbonyl Compounds 3-Deoxyglucosone and 3-Deoxymaltosone during Beer Fermentation by Saccharomyces Yeasts" Fermentation 10, no. 6: 281. https://doi.org/10.3390/fermentation10060281