Abstract
A substantial body of research indicates that the gut microbiota exerts a profound influence on host health. The purpose of this work was to characterize selected, most promising, well-known next-generation probiotics (NGPs) and review the potential applications of the bacteria in food technology. The isolation of gut bacteria with significant health benefits has led to the emergence of NGPs. In contrast to traditional probiotics, these originate directly from the gut microbiota, thereby ensuring their optimal adaptation to the intestinal ecosystem. NGPs exert their effects on the host organism through a variety of mechanisms, including the synthesis of bioactive compounds, modulation of the gut microbiota, and metabolism of substances provided by the host. Several bacterial species have been identified as potential candidates for NGPs, including Akkermansia muciniphila, Faecalibacterium prausnitzii, Bacteroides thetaiotaomicron, Christensenella minuta, and many others. These bacteria have demonstrated the capacity to exert beneficial effects, including the reduction of obesity, type 2 diabetes, metabolic disorders, and even cancers. The greatest limitation to their commercialization is their lack of oxygen tolerance, which presents challenges not only for research but also for their potential application in food. The most optimal approach for their application in food appears to be microencapsulation. Further research is required to establish the safety of NGP supplementation and to protect them from environmental conditions.
1. Introduction
Food plays a crucial role in the prevention of non-communicable diseases, including lifestyle-related diseases [1]. In recent years, there has been a noticeable increase in interest in healthy lifestyles, known as the well-being trend, which is defined as a healthy lifestyle that focuses on improving physical, mental, and social well-being [2]. It is typically motivated by the desire to improve quality of life, increase energy levels, improve mental health, or reduce the risk of developing chronic diseases [3,4].
The growing awareness of health and nutrition is also reflected in the significant development of the functional food market. Functional foods are those that, in addition to their basic nutritional functions, provide additional health benefits through the addition of bioactive substances or components [5]. Examples include the fortification of foods with vitamins, minerals, prebiotics, or probiotics, with probiotic foods accounting for approximately 60–70% of the total functional food market [5,6,7].
The functional food market is experiencing dynamic growth due to the increasing demand for healthier food products. It is estimated that the global functional and natural health food market was valued at USD 23.5 billion and is projected to increase to USD 38.5 billion by 2033 [8]. This growth is being driven by innovation in new food products, their increased availability, and rising consumer nutritional awareness. Dairy products have the largest market share due to their high bioactive content and consumer trust. The functional food market is expanding, particularly in developed countries, driven by widespread access to health information and increased total awareness [9,10].
Probiotic foods currently available on the market contain live cultures of bacteria belonging mostly to the Bifidobacterium and previous Lactobacillus genera, which have been shown to have beneficial effects on the body’s systemic functions. In 2020, scientists reclassified the genus Lactobacillus into 25 genera, including the emended genus Lactobacillus and 23 novel genera [11]. The vast majority of probiotic bacteria currently available belong to these genera because they are easy to cultivate under laboratory conditions, allowing their probiotic properties to be studied in detail [12,13]. Consumers are increasingly choosing fermented products, such as dairy-based fermented foods, but also fermented vegetables and fruits. Although supplementation with probiotic bacteria in pharmaceutical forms is also popular, probiotic foods often attract more interest because of their naturalness [14,15,16].
Fermentation using probiotic bacteria, as opposed to wild fermentation, plays a crucial role in the controlled production of functional foods. During the fermentation process, these bacteria are capable of synthesizing a wide range of bioactive compounds, such as organic acids, short-chain fatty acids (SCFAs), B and K vitamins, bioactive peptides, exopolysaccharides, as well as bacteriocins, which have synergistic antimicrobial activity against pathogenic bacteria, thereby extending the shelf life of these products. In addition, the metabolome of probiotic bacteria also includes gamma-aminobutyric acid (GABA), a neurotransmitter that can act through the gut–brain axis and, therefore, influence human well-being [15,17].
An exceptionally new direction in research into the effects of biotics on the human body is the study of postbiotics, which are defined as a preparation of inanimate microorganisms and/or their components that confer a health benefit on the host [18]. Due to their lack of fermentative ability and, consequently, no interference with the sensory properties of products, together with their confirmed health benefits for the human body, postbiotics represent a promising direction for application in food technology and the development of functional foods [19,20].
The high level of consumer interest in probiotic bacteria and/or postbiotics is linked to numerous reports from both the media and scientists on their wide-ranging effects on human health, including immune enhancement, mental health improvement, and gut health. By helping to maintain the homeostasis of the gut microbiota and inhibiting the growth of pathogenic bacteria, probiotics can not only alleviate symptoms of gastrointestinal disorders but also reduce the risk of cancer and intestinal inflammation and improve metabolic efficiency [19,21,22]. In addition to improving systemic bodily functions, probiotics also have a significant impact on mental health through the gut–brain axis and on skin health, which can be exploited in the beauty industry and in products aimed at individuals seeking to improve their appearance [23,24].
The purpose of this work was to characterize the selected gut bacteria in the context of the next-generation probiotics (NGPs) concept and to discuss the potential of their applications in food technology. The novelty of this review is related to the dynamic development of the NGP knowledge. Although several reviews have been published recently [25,26,27], none of them focused on the challenges associated with the application of NGPs in food technology. Therefore, in this work, we collected information on the advantages and disadvantages of adding NGPs to food and also identified the possibilities and limitations related to their use in food technology.
2. Gut Microbiota
The term “gut microbiota” refers to the collective of all commensal, symbiotic, and pathogenic microorganisms that inhabit the intestines of the host organism. This encompasses not only bacteria, which are the most extensively studied, but also viruses, archaea, protozoa, and fungi. The Human Microbiome Project, initiated in 2007, aimed to create a genomic dataset of microorganisms residing in various human body niches. This research project marked the beginning of a new era in the study of the significant influence of the gut microbiota on host functioning and revolutionized our understanding of the microbiota and its crucial role in shaping health [28].
The gut microbiota is estimated to comprise approximately 1014 microbial cells, which is comparable to the number of human cells. The microbiota is highly diverse, comprising over 1000 different bacterial species. The collective genome of these microorganisms (the microbiome) is approximately 150 times larger than the human genome, indicating an immense metabolic potential. The sum of the human and microbial genes forms a complex metabolic system that is unattainable without bacterial genomes [24]. This system is termed a “superorganism”, resulting from the symbiotic relationship among thousands of species. As a consequence of the evolutionary development of interactions between the microbiota and the host organism, these entities influence each other through genetic, metabolic, and immunological signals. Since the majority of microorganisms that constitute the human microbiota are strictly anaerobic, microbiologists are only able to culture approximately 20–60% of these bacteria using traditional techniques. Consequently, metagenomics is employed to conduct precise studies of the microbiota [29,30].
The gut microbiota exerts a direct influence on human health due to its capacity to synthesize a diverse range of bioactive compounds that affect multiple human body systems, including the immune, nervous, and digestive systems. Moreover, the gut microbiota interacts closely with the gut-associated lymphoid tissue (GALT), which is essential for the impact on the host. On the one hand, the gut microbiota supports the maturation of immune cells and the production of cytokines and protects against pathogens, thereby influencing the essential functioning of the immune system. Conversely, the gut microbiota is linked to the central nervous system (CNS) via the gut–brain axis, which affects mental health, mood regulation, and cognitive functions [27]. Recently, other microbiota axes have also been discovered, such as the microbiota–bone axis, the microbiota–liver axis, and the microbiota–lung axis [31]. Dysbiosis, or the disruption of gut microbiota homeostasis, is associated with the development of numerous diseases, including obesity, type II diabetes, inflammatory bowel diseases, neurodegenerative diseases, osteoporosis, hepatic diseases, respiratory diseases such as asthma, chronic obstructive pulmonary disease, and certain cancers [23,32,33].
2.1. The Role of Food in Modulating the Gut Microbiota
The dietary choices of humans have a profound impact on the diversity and composition of their gut microbiota. An imbalanced diet has a detrimental effect on the changes occurring within the gut microbiota, while the provision of appropriate substances, including probiotics, has a wide-ranging impact on health modulation [26]. A diet rich in dietary fiber, prebiotics, probiotics, and fermented products may beneficially modulate the composition of the gut microbiota by increasing the abundance of health-promoting bacteria and limiting potential pathogens. Research indicates that a Western-type diet, characterized by high sugar and saturated fat intake, disrupts the homeostasis of the gut microbiota. In contrast, a Mediterranean diet contributes to increased microbial diversity within the microbiota and significant improvements in health [32,34,35].
Among the nutrients provided in the diet, carbohydrates appear to be of particular significance, serving as a nutritional substrate for gut microorganisms. Soluble fiber serves as a source of nourishment for microorganisms in the colon, resulting in the synthesis of a range of compounds, including SCFAs. A diet deficient in prebiotic substances, which are part of soluble carbohydrates, has been shown to result in a reduction in the growth of Bacteroides thetaiotaomicron bacteria, which degrade mucin glycans. This, in turn, affects the thinning of the intestinal barrier [36,37].
In addition to carbohydrates, other components present in the diet also exhibit significant effects. In vitro studies have demonstrated that the fermentation of plant proteins increases the abundance of Lactobacillus and Bifidobacterium bacteria and stimulates the synthesis of SCFA while contributing to a decrease in the abundance of harmful bacteria such as Bacteroides fragilis and Clostridium perfringens [36]. It has been postulated that a high consumption of fat, particularly saturated fatty acids (SFAs), may result in gut dysbiosis. A high intake of SFA-rich diets has been demonstrated to stimulate the growth of sulfate-reducing bacteria, which have been shown to negatively impact the intestinal mucus layer, leading to intestinal inflammation, intestinal diseases, and colitis [36,38]. Omega-3 fatty acids may exert a beneficial influence on gut microbiota alterations by increasing the synthesis of anti-inflammatory compounds and SCFA acetate. The ratio of omega-3 to omega-6 fatty acids in the diet should be 1:1 [39].
A lack of homeostatic regulation contributes to an increased permeability of the gut and the development of metabolic endotoxemia. A diet high in sugar has been shown to cause similar gut microbiota modulation to that induced by a diet high in saturated fatty acids [36,40]. Moreover, scientists indicate that artificial sweeteners such as aspartame, saccharin, and acesulfame K affect the modulation of the gut microbiota by reducing the quantity of Clostridiales, Lactobacillales, and Bifidobacterium bacteria while increasing the abundance of Enterobacteriaceae. These changes result in elevated fasting glucose levels and impaired insulin response, which, in turn, leads to decreased glucose tolerance [36,41]. In vitro studies have demonstrated that emulsifiers, including lecithins, mono- and diglycerides of fatty acids, and carboxymethylcellulose, can increase bacterial translocation across the intestinal epithelium, thereby promoting systemic inflammation by altering the location and composition of the microbiota. A reduction in the levels of SCFAs in feces and free amino acids has been documented. These alterations result in gut dysbiosis, which, in turn, promotes the development of colitis and metabolic disorders [36].
The principal agents involved in the modulation of gut microbiota encompass prebiotics, probiotics, polyphenols, and fermented products [14,35]. Prebiotics are dietary substrates that selectively promote the proliferation or activity of beneficial bacteria indigenous to the colon. Prebiotic substances, such as inulin and fructooligosaccharides, are essential for selectively stimulating the growth of beneficial bacteria like Bifidobacterium and Lactobacillus. Fermented products, including yogurt, kefir, sauerkraut, and miso, are natural sources of potentially probiotic bacteria. These may influence gut microbiota by introducing beneficial bacteria and enhancing its diversity [14,35].
Probiotics are defined as live microorganisms that, when administered in adequate amounts, confer health benefits to the host. They have been designated as generally recognized as safe (GRAS) at the strain level by the United States Food and Drug Administration (FDA) or as qualified presumption of safety (QPS) at the species level by the European Food Safety Authority (EFSA) [42,43]. Probiotic supplementation may result in short-term alterations to the composition of the gut microbiota. Supplementation may increase the abundance of beneficial bacteria, which may be helpful in the treatment of gastrointestinal disorders or other diseases. Nevertheless, the long-term visibility of these alterations is contingent upon the continuous administration of probiotics and the availability of suitable nutritional substrates [44,45].
Probiotics are capable of modulating gut microbiota through a range of mechanisms. These include competition for nutrients and adhesion sites, production of antimicrobial substances (bacteriocins), and modulation of the host’s immune system. Dietary fiber can be fermented by the microbiota into SCFAs, which have a wide range of health benefits. These include the regulation of glucose and lipid metabolism, as well as the modulation of the immune response [46,47].
2.2. The New Generation of Probiotics and Postbiotics
The analysis of the impact of bacteria on human health has been ongoing for at least a decade, with a primary focus on the microbiota present in food. The advancement of technology has facilitated the advancement of scientific progress through the utilization of more precise methodological tools. Modern studies focusing on the analysis of the impact of specific microbiota components on the host organism allow the identification of health-promoting bacteria that significantly influence bodily functions. Following the advancement of scientific knowledge, NGPs have been defined as live microorganisms identified based on comparative analyses of the microbiota. When administered in appropriate amounts, these NGPs confer health benefits to the host [43,48]. The distinction between probiotic and NGP definitions is primarily methodological, with NGPs identified within the human natural microbiota, for which significant health impacts have been demonstrated. NGPs have been recently isolated using novel, sophisticated techniques for isolating, identifying, and even modifying these commensal bacteria. They have been identified primarily through comparative analyses of microbiota composition in both healthy and unhealthy individuals, and they belong to various genera [43,49].
The principal concerns regarding NGPs relate to their efficacy, safety, and technological robustness. The majority of gut bacteria that have been identified as potential NGPs are nutritionally fastidious and highly sensitive to oxygen. This makes them challenging to produce and maintain in large quantities on an industrial scale, as well as to process and formulate into products [50]. The administration of traditional probiotics is not intended to combat specific diseases. In light of these considerations, the urgent need for the identification and characterization of new, disease-specific NGPs has become apparent. Concerning NGPs, there is a growing recognition of their potential utility as postbiotics, which are non-viable bacterial products or metabolic byproducts of NGP microorganisms that exhibit biological activity in the host [43,48,51].
It is of paramount importance to maintain the health of the intestinal microbiota to preserve homeostasis. This is achieved through the maintenance of microbial abundance and diversity. The right number of diverse microbiota ensures the existence of multiple metabolic pathways, thereby enhancing the host organism’s capacity to adapt to various environmental challenges and dietary changes [28,37]. In light of the available scientific data, it can be claimed that among the important, health-promoting bacteria present in a healthy gut microbiota, there are a number of potential candidates for NGPs, like Akkermansia muciniphila, Faecalibacterium prausnitzii, Bacteroides thetaiotaomicron, Christensenella minuta, and a few others.
2.2.1. Akkermansia muciniphila
Akkermansia muciniphila is a non-spore-forming, Gram-negative bacterium. Although originally classified as an obligate anaerobe, studies have shown that it tolerates low oxygen levels similar to Bacteroides fragilis and Bifidobacterium adolescentis, which can survive for 24 h when exposed to ambient air. Discovered in 2004, it was the first bacterium from the phylum Verrucomicrobia to be identified in the human gut [52]. A. muciniphila is abundant in the intestinal mucus layer, where mucin serves as its primary source of carbon and nitrogen [53]. It is one of the most important gut symbionts, colonizing the gut within the first year of life and persisting throughout life, constituting approximately 1–4% of the total gut microbiota, making it one of the twenty most abundant species. Under laboratory conditions, it can be cultured on synthetic media where glucosamine-6-phosphate, found in mucin, is essential. Research has shown that A. muciniphila cultured on synthetic media is safe for use in human nutritional supplementation [52,54,55].
Following the discovery of A. muciniphila and the recognition of its significant abundance in the core gut microbiota, numerous studies have been conducted to elucidate its impact on human health. These studies have involved both laboratory animals and, in later stages, human participants. Research has demonstrated the critical role of A. muciniphila in obesity-related parameters in animal models, including reduced body weight, fat mass, hip circumference, and caloric intake [56]. The administration of A. muciniphila to mice resulted in enhanced insulin sensitivity, accompanied by elevated levels of anti-inflammatory cytokines, which collectively contributed to a reduction in inflammation. Studies have demonstrated the efficacy of A. muciniphila supplementation in the treatment of obesity, type 2 and type 1 diabetes, fatty liver disease, inflammatory bowel disease (particularly ulcerative colitis), and various cancers, including colon cancer, by modulating the immune system in mice [57,58]. Furthermore, mouse studies have demonstrated the protective effect of A. muciniphila against egg albumin allergy, attributed to reduced jejunal damage and modulation of the immune system [59]. These findings indicate that these bacteria play an important role in modulating metabolic processes. However, further human studies are necessary to fully elucidate the effects and mechanisms of these bacteria on human health. Furthermore, research has investigated the influence of A. muciniphila on the development of neurodegenerative, neurodevelopmental, and autoimmune diseases [60]. However, the results have been inconsistent, and a causal relationship has not been established. A. muciniphila has been described as having beneficial effects on health, particularly by reducing mucosal permeability and the expression of pro-inflammatory cytokines. Moreover, a study conducted by Parrish et al. (2023) indicates that the impact of this bacterium is context-dependent and may be harmful in cases of food allergies when the microbiota is deprived of dietary fiber [61].
A promising discovery was the demonstration of an enhanced health-promoting effect of A. muciniphila following prior pasteurization (30 min at 70 °C) in relation to diet-induced metabolic disorders in mice. Pasteurized A. muciniphila also significantly improved glucose tolerance and hepatic insulin sensitivity and completely blocked diet-induced metabolic endotoxemia. It has been demonstrated that the health-promoting effect is attributed to proteins located on the outer membrane of the bacterium, with one of the most abundant being the Amuc_1100 protein [57,62]. The increased efficacy and safety of pasteurized A. muciniphila cells demonstrated in studies involving mice and humans led to the approval of the use of such a preparation in human dietary supplementation. EFSA has determined that daily supplementation of 3.4 × 1010 pasteurized cells is considered safe, provided that the number of viable cells is less than 10 colony-forming units per gram (CFU/g). This opinion has enabled the use of pasteurized A. muciniphila cells in food technology, allowing the creation of functional foods with their addition [60].
2.2.2. Faecalibacterium prausnitzii
Faecalibacterium prausnitzii is a strictly anaerobic, Gram-positive bacterium belonging to the family Ruminococcaceae. In laboratory conditions, it is challenging to culture, even under anaerobic conditions. It is one of the most prevalent species in the gut microbiota, accounting for 1–6% of the total bacterial population. F. prausnitzii is one of the most important producers of butyric and salicylic acid in the gut, earning it the colloquial name “gut guardian” [63].
These metabolites exhibit crucial immunomodulatory effects, thereby protecting the colon from inflammation and the development of colorectal cancer. Butyrate serves as the primary energy source for colonocytes. Studies have demonstrated that the concentration of salicylic acid present in the colon can reduce the level of the pro-inflammatory cytokine IL-8 in vitro. The reduction of IL-8 by salicylic acid and butyrate is due to the inhibition of NF-κB activation. Furthermore, the inhibition of cytokines IL-6, IL-12, IFN-γ, and TNF-α has been demonstrated to occur through the induction of IL-10 production. In a mouse study, the use of F. prausnitzii bacteria cells or cell-free supernatant was found to significantly reduce the severity of 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced colitis. A significant decrease in the abundance of F. prausnitzii has been observed in patients with obesity, type 2 diabetes, and inflammatory bowel diseases, including ulcerative colitis, Crohn’s disease (CD), colorectal cancer, and psoriasis. These findings indicate that F. prausnitzii deficiency is associated with reduced anti-inflammatory activity and the development of inflammation. Studies involving pregnant women have demonstrated that the quantity of F. prausnitzii increases during the first trimester, while in the third trimester, the quantity decreases. Some authors suggested that the abundance of F. prausnitzii, which is attributed to the production of butyrate and other anti-inflammatory substances, could contribute to successful pregnancy outcomes; however, further studies are needed to prove this concept. Additionally, F. prausnitzii has been shown to regulate gut barrier function by potentially increasing the levels of tight junction proteins occludin and E-cadherin. This reduces colon permeability and protects the host from potential harm [64,65,66,67].
The unavailability of an active pharmaceutical preparation containing F. prausnitzii bacteria currently precludes the possibility of modulating its abundance through the administration of pharmaceuticals. Instead, dietary interventions based on prebiotic substances and lifestyle modifications represent the most viable avenues for achieving this goal. However, the high nutritional requirements and strict anaerobic nature of F. prausnitzii present a challenge for its commercialization [63,64]. Nevertheless, it has been demonstrated that antioxidants commonly found in the gut, such as cysteine and riboflavin, facilitate the survival of these bacteria upon exposure to air, allowing the highly oxygen-sensitive bacteria to remain viable for up to 24 h with air contact. This finding paves the way for further research and the future commercialization of the bacterium [68].
2.2.3. Bacteroides thetaiotaomicron
Bacteroides thetaiotaomicron is a Gram-negative, anaerobic, non-spore-forming rod-shaped bacterium. It is among the most prevalent bacteria in the gut microbiota, comprising approximately 6% of the total population, thereby underscoring its significant role in the core microbiota common to all humans. A genome analysis conducted in 2003 revealed the extensive metabolic capabilities of this bacterium and its potential to encode antibiotic resistance, including resistance to erythromycin and tetracycline. Such resistances can be transferred to other bacteria within the gut microbiota [69]. Due to its numerous health benefits, it has been identified as a potential NGP candidate [70].
B. thetaiotaomicron exhibits the ability to metabolize a highly diverse range of polysaccharides that would otherwise remain unprocessed, including amylose, amylopectin, and pullulan. Its repertoire of glycolytic enzymes is among the largest known in prokaryotes, and it is believed to be capable of hydrolyzing nearly all glycosidic bonds present in polysaccharides [71,72,73]. During its metabolic processes, it synthesizes substances such as acetic acid, propionic acid, and succinic acid, which serve as energy sources for host cells and for the bacterium F. prausnitzii, thereby participating in the production of intestinal mucus [74]. In addition to SCFAs and hydrogen sulfide (H2S), the presence of B. thetaiotaomicron is associated with the secretion of outer membrane vesicles (OMVs) [75,76,77].
A series of studies conducted on mice have demonstrated that B. thetaiotaomicron is capable of inhibiting the growth of C. difficile in a state of gut dysbiosis caused by the excessive proliferation of this pathogen [78,79]. This is achieved by restoring gut microbiota homeostasis and improving the bile acid profile. The production of enzymes such as galactanase by B. thetaiotaomicron enhances the prebiotic activity of galactooligosaccharides and promotes the growth of probiotic bacteria such as Lactobacillus and Bifidobacterium [80,81]. This phenomenon is referred to as mutual cross-feeding in vivo [82,83]. Furthermore, it has been demonstrated that B. thetaiotaomicron induces the expression of the proline-rich protein Sprr2A, which is essential for maintaining desmosomes in the gastrointestinal tract [84]. Furthermore, this bacterium has been demonstrated to enhance goblet cell differentiation, the expression of mucus-related genes, and the ratio of sialylated to sulfated mucins in rats. This study indicated a complementary metabolic relationship between B. thetaiotaomicron and F. prausnitzii, resulting in the modulation of the intestinal barrier [74]. In vivo studies have demonstrated that B. thetaiotaomicron exerts regulatory influence on the network of enteroendocrine cells in various regions of the gastrointestinal tract [85]. Furthermore, B. thetaiotaomicron exhibits immunomodulatory effects by antagonizing NF-κB and reducing pro-inflammatory cytokines, especially TNF-α, as well as stimulating the expression of IL-6 and IL-10 [76,86]. This is also attributed to the presence of OMVs. Due to their pleiotropic effects on the host, these bacteria exhibit anti-inflammatory properties, enhance resistance to pathogens, and process important dietary nutrients [76,77,87]. A series of human studies involving 16–18-year-olds with Crohn’s disease have confirmed the safety of supplementation. However, further research is needed to fully establish its safety [86].
Dysbiotic disturbances associated with both excess and deficiency of B. thetaiotaomicron have been linked to a range of serious health consequences. As these bacteria are both commensal and opportunistic pathogens, they can cause significant health issues, such as the accumulation of sialic acid, which promotes the growth of pathogenic bacteria such as C. difficile or E. coli [88]. The excessive proliferation of B. thetaiotaomicron and the subsequent damage to the intestinal barrier can result in the development of colorectal cancer, as well as their translocation into the bloodstream, leading to bacteremia [75]. Given the significant health benefits of B. thetaiotaomicron and the potential risks associated with its use, its application in supplementation and food technology should be carefully monitored and used in justified cases, such as personalized nutrition [89].
2.2.4. Christensenella minuta
Christensenella minuta is a strictly anaerobic, non-spore-forming, Gram-negative bacillus. It is the first representative of the family Christensenellaceae within the phylum Firmicutes, isolated in 2012. Despite its relatively recent discovery, this species has already undergone extensive study, which has highlighted its significant health implications, even though it is a subdominant member of the gut microbiota. Notably, despite its anaerobic nature, C. minuta is capable of tolerating prolonged exposure to atmospheric oxygen. Its primary metabolic products from glucose are acetic and butyric acids [90]. To date, research has elucidated a multifaceted mechanism of action for C. minuta, which includes modulating the gut microbiota, producing SCFAs, influencing lipid metabolism, regulating the integrity of the intestinal epithelium, and modulating bile acid metabolism [91,92,93].
The aforementioned mechanisms of C. minuta have been demonstrated to significantly reduce the risk of obesity. This is achieved through the regulation of the Firmicutes/Bacteroidetes ratio, the influence of SCFAs on leptin synthesis, and the modulation of lipid and glucose metabolism precursors [93,94]. Additionally, C. minuta increases thermogenesis in adipose tissue, enhances the expression of tight junction proteins in the intestines, and affects the metabolism of bile acids [92,95]. In vitro and in vivo studies have demonstrated that C. minuta exhibits anti-inflammatory and immunomodulatory effects. These effects are achieved through the inhibition of the NF-κB signaling pathway and the suppression of pro-inflammatory cytokines IL-8 and IL-1β [96]. Additionally, C. minuta stimulates the production of the anti-inflammatory cytokine IL-10. Furthermore, C. minuta has been demonstrated to facilitate mucosal healing and reduce neutrophil activation due to inflammation in two distinct animal models of acute colitis and in human intestinal cell lines. These studies indicate that C. minuta plays a pivotal role in inflammatory bowel diseases (IBD), where its efficacy is comparable to that of mesalazine, a common treatment for IBD. Metagenomic analyses have revealed that patients with obesity, IBD, and CD have significantly lower levels of C. minuta in their gut microbiota. Furthermore, research has indicated that C. minuta impacts the development of type 2 diabetes by inhibiting the expression of SGLT1 and GLUT2 in intestinal glucose transport and by promoting the secretion of GLP-1, which stimulates insulin sensitivity and regulates glucose homeostasis [90,91,97,98,99]. Mouse studies have demonstrated that inoculation with C. minuta is associated with increased physical activity and reduced feed intake. These findings suggest a potential role for this bacterium in regulating metabolic processes and influencing the gut–brain axis [100].
At present, there is no evidence to suggest that C. minuta contributes to the development of diseases. Nevertheless, given the paucity of human studies, further research is required to ascertain its safety.
3. A Perspective on the Potential Use of These Types of Bacteria in Food Technology
The principal concerns regarding NGPs relate to their efficacy, safety, and technological robustness. The majority of gut bacteria that have been identified as potential NGPs are nutritionally fastidious and highly sensitive to oxygen. This makes them challenging to produce and maintain in large quantities on an industrial scale, as well as to process and formulate into products. On the other hand, the administration of traditional probiotics is not intended to combat specific diseases. In light of these considerations, the urgent need for the identification and characterization of new, disease-specific NGPs has become apparent [43,48]. The potential for the development of products with such specific characteristic NGPs in traditional food or capsule-based supplement forms could yield substantial economic benefits, driven by the current trend towards a healthy lifestyle [6,8,19]. The selection of suitable delivery vehicles for probiotics is an important aspect since it may positively or negatively affect cell viability and effective target release in the gut. Although probiotics can be delivered as pharmaceutical formulations or food-based products, it is considered that consuming probiotics in food may seem a more natural way of receiving a regular dose. As a result, foods are becoming more widely accepted by consumers as effective probiotic delivery vehicles [101].
Figure 1 presents the overview of next-generation biotics (NGBs) applications in food technology. The term NGBs that we propose include next-generation probiotics, defined as life bacterial cells, as well as their metabolites (postbiotics).
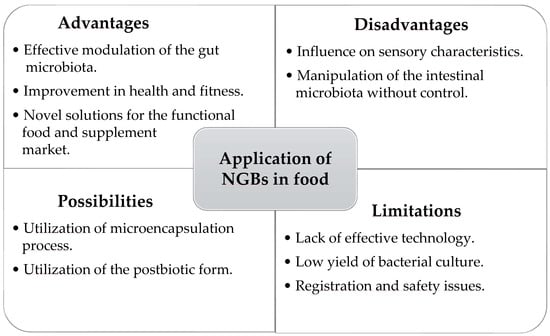
Figure 1.
Next-generation biotics (NGBs) applications in food technology—advantages, disadvantages, perspectives, and limitations.
A number of studies have been conducted with the objective of analyzing the survival of anaerobic bacteria in yogurt following the addition of oxygen-consuming bacteria, such as S. thermophilus. However, the results were not as anticipated due to the high acidification of the product by the adjunct bacteria, which did not protect the anaerobic bacteria present in the product after opening and consumption [102]. Consequently, the utilization of NGPs should not be focused on their technological incorporation during production but rather on enriching food through supplementation in a form that protects them from exposure to air.
One potential solution could be the use of microencapsulation. It is essential that the microencapsulation process is conducted in an appropriate manner, utilizing innovative materials and minimizing the impact of oxygen on the bacteria [103,104,105,106,107]. The materials employed in the microencapsulation process must be safe and well-digested by the human body. Moreover, the encapsulated bacteria must be able to survive in the presence of these materials, which may be challenging for bacteria like B. thetaiotaomicron due to their enzymatic apparatus [70,103]. Additionally, the outer layer of the microcapsule should be hydrophobic and resistant to pH changes, thus enabling their full utilization in food production, such as in fermented dairy products, beverages, and others. This approach would render their application virtually limitless in food products that are not subjected to high temperatures. However, it is likely that these products would require refrigeration temperatures for storage [16]. It is crucial to recognize that the achievement of a health benefit necessitates not only supplementation but also the continuous maintenance of gut microbiota homeostasis. Consequently, these products should be based on health-promoting raw materials that are desirable to probiotic gut bacteria species. The production of microencapsulated NGPs would facilitate their use in the manufacture of functional foods, nutraceuticals, and pharmaceuticals with a broad range of health benefits [25,27].
Another potential avenue for the utilization of NGPs is the development of postbiotics derived from them. Research indicates that bacterial metabolites and cell wall components exhibit a wide range of health-promoting effects. The use of postbiotics would be significantly more straightforward due to the absence of the need to protect bacterial cells from oxygen exposure. This approach would entail cultivating NGPs under optimal conditions to obtain health-promoting supernatants, followed by the concentration or isolation of the desired bioactive substances, and then incorporating these substances into food. The potential applications of postbiotics in food products also appear to have a wide range of possibilities, including traditional fermented products, sweets, and meat products [19,20].
In the near future, the bacterium A. muciniphila may be employed. This is related to the proven enhanced probiotic effects of their pasteurized form and its recognition as safe by the EFSA. It is likely that its use would also be based on the supplementation of food to impart probiotic properties. The pasteurized form of the bacteria is not limited to use exclusively in refrigerated food products [60,108].
4. Limitations and Prospects for the Use of Next-Generation Probiotics in Food Supplementation and Technology
The application of NGPs in food represents a significant advancement in the field of functional foods, marking a new chapter in the perception of food. This would entail that food would not only have a preventative impact on human health but could also act as a natural next-generation medicine [27]. However, this necessitates the advancement of knowledge and technology due to significant limitations.
The most significant technological challenge in the development of this sector of food is the lack of tolerance to the presence of oxygen for the growth and survival of NGPs [109]. This presents a substantial problem that almost entirely prevents the use of these bacteria for fermentation, such as using B. thetaiotaomicron to partially metabolize carbohydrates to obtain intermediate products with health-promoting effects. Moreover, their use in fermentation would likely be highly unacceptable to consumers from a sensory perspective due to the synthesis of a range of specific aromatic compounds, including phenols, indoles, sulfur compounds, amines, and BCFA esters [110,111].
The development of technology enabling the application of NGPs in their live or postbiotic form in food production requires substantial financial investment and the collaboration of scientists from various fields, as well as food manufacturers. The optimization of laboratory cultivation methods is of paramount importance if the mass cultivation and commercialization of these bacteria are to be facilitated. Furthermore, the development of technology to minimize oxygen exposure is of great importance, with microencapsulation appearing to be the most promising process. Once suitable commercialization and protection methods for the bacteria have been identified, food technologists will be required to analyze the impact of NGPs on food and its acceptability [5,25,106].
Nevertheless, prior to the implementation of NGPs in food products, further research is required to ascertain the safety of specific bacterial strains. Without their recognition as safe, it is impossible to prepare appropriate legal regulations permitting their use in food production. Therefore, a comprehensive strain characterization is required, including the whole-genome sequence analysis, as well as virulence and antibiotic resistance genes or transferrable genetic elements. Safety assessment in animal models of specific diseases and human clinical trials should also be assessed [101]. Since today, the oral administration of live and pasteurized A. muciniphila has been reported to be safe and well-tolerated [112,113]. Also, the safety of F. prausnitzii, B. thetaiotaomicron, and C. minuta has been reported in animal and human studies [49,86,114].
Another important issue is the registration of NGPs as food additives. It should be underlined that these bacteria have no history of safe use in the food industry, unlike traditional probiotics. Therefore, they cannot have generally recognized as safe (GRAS) status in the USA or qualified presumption of safety (QPS) status of the European Food Safety Authority (EFSA). NGPs, as well as postbiotics from NGPs, must be treated according to the novel food EU regulation when applied in food [49,101]. Although the regulations in Europe and other countries like the US, Canada, and Japan lack harmonization, the path to the application of NGPs in both food and pharmaceutical supplementation is still long, but the prospects are very promising.
5. Summary
The advancement of scientific knowledge pertaining to the analysis of the impact of gut microbiota on human health is transforming the way in which food is perceived to influence the body. The isolation of bacteria with significant effects on host function, followed by their precise analysis and commercialization, represents a significant challenge that requires the involvement of scientists from a range of disciplines. The commercialization of next-generation probiotics and their application in food production could revolutionize the market of functional foods, provided that they are safe and acceptable to consumers. Although the path to the use of NGPs in food production is still long, the prospects for their use in this context are promising. A multitude of studies are still required to ascertain the safety of specific NGP strains and to facilitate their commercialization.
Author Contributions
Conceptualization, D.Z. and P.L.; methodology, P.L. and D.Z.; software, P.L. and D.Z.; validation, D.Z.; formal analysis, D.Z.; writing—original draft preparation, P.L.; writing—review and editing: D.Z.; visualization, P.L.; supervision, D.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gille, D.; Schmid, A.; Walther, B.; Vergères, G. Fermented Food and Non-Communicable Chronic Diseases: A Review. Nutrients 2018, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Grunert, K.G.; do Canto, N.R.; Liu, R.; Salnikova, E. Well-being as a global food trend: Health, sustainability and authenticity. Dan. Food Innov. Pap. 2019. [Google Scholar]
- Zwierczyk, U.; Sowada, C.; Duplaga, M. Eating Choices—The Roles of Motivation and Health Literacy: A Cross-Sectional Study. Nutrients 2022, 14, 4026. [Google Scholar] [CrossRef] [PubMed]
- Baz, S.S.B.; Malibarey, W.M.; Alsalmi, H.A.; Alzaydi, M.D.; Alqahtani, A.M.; Alghamdi, R.Y.; Sr, S.S.B.B.; Ii, W.M.M.; Sr, H.A.; Alzaydi, M.D.; et al. The Impact of a Healthy Lifestyle on Psychological Well-Being Among Saudi Adolescent Girls Attending Secondary Schools in Taif City, Saudi Arabia. Cureus 2023, 15, e50189. [Google Scholar] [CrossRef]
- Granato, D.; Barba, F.J.; Kovačević, D.B.; Lorenzo, J.M.; Cruz, A.G.; Putnik, P. Functional Foods: Product Develop-ment, Technological Trends, Efficacy Testing, and Safety. Annu. Rev. Food Sci. Technol. 2020, 11, 93–118. [Google Scholar] [CrossRef]
- Küçükgöz, K.; Trząskowska, M. Nondairy Probiotic Products: Functional Foods That Require More Attention. Nutrients 2022, 14, 753. [Google Scholar] [CrossRef]
- Goetzke, B.; Nitzko, S.; Spiller, A. Consumption of Organic and Functional Food. A Matter of Well-Being and Health? Appetite 2014, 77, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Functional and Natural Health Food: Market Value Forecast Worldwide 2023–2033. Available online: https://www.statista.com/statistics/502267/global-health-and-wellness-food-market-value/ (accessed on 8 June 2024).
- Shiby, V.K.; Mishra, H.N. Fermented Milks and Milk Products as Functional Foods—A Review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Branco, G.F.; Cruz, A.G.; de Faria, J.A.F.; Shah, N.P. Probiotic Dairy Products as Functional Foods. Compr. Rev. Food Sci. Food Saf. 2010, 9, 455–470. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Ghelardi, E.; Mazzantini, D.; Celandroni, F.; Calvigioni, M.; Panattoni, A.; Lupetti, A.; Bois De Fer, B.; Perez, M. Analysis of the Microbial Content of Probiotic Products Commercialized Worldwide and Survivability in Conditions Mimicking the Human Gut Environment. Front. Microbiol. 2023, 14, 1127321. [Google Scholar] [CrossRef] [PubMed]
- Aponte, M.; Murru, N.; Shoukat, M. Therapeutic, Prophylactic, and Functional Use of Probiotics: A Current Perspective. Front. Microbiol. 2020, 11, 562048. [Google Scholar] [CrossRef]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Negrete-Romero, B.; Valencia-Olivares, C.; Baños-Dossetti, G.A.; Pérez-Armendáriz, B.; Cardoso-Ugarte, G.A. Nutritional Contributions and Health Associations of Traditional Fermented Foods. Fermentation 2021, 7, 289. [Google Scholar] [CrossRef]
- Kruk, M.; Lalowski, P.; Hoffmann, M.; Trząskowska, M.; Jaworska, D. Probiotic Bacteria Survival and Shelf Life of High Fibre Plant Snack—Model Study. Plant Foods Hum. Nutr. 2024. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Butani, K.; Kumar, A.; Singh, S.; Prajapati, B.G. Effects of Fermented Food Consumption on Non-Communicable Diseases. Foods 2023, 12, 687. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Vera-Santander, V.E.; Hernández-Figueroa, R.H.; Jiménez-Munguía, M.T.; Mani-López, E.; López-Malo, A. Health Benefits of Consuming Foods with Bacterial Probiotics, Postbiotics, and Their Metabolites: A Review. Molecules 2023, 28, 1230. [Google Scholar] [CrossRef]
- Aggarwal, S.; Sabharwal, V.; Kaushik, P.; Joshi, A.; Aayushi, A.; Suri, M. Postbiotics: From Emerging Concept to Application. Front. Sustain. Food Syst. 2022, 6, 887642. [Google Scholar] [CrossRef]
- Latif, A.; Shehzad, A.; Niazi, S.; Zahid, A.; Ashraf, W.; Iqbal, M.W.; Rehman, A.; Riaz, T.; Aadil, R.M.; Khan, I.M.; et al. Probiotics: Mechanism of Action, Health Benefits and Their Application in Food Industries. Front. Microbiol. 2023, 14, 1216674. [Google Scholar] [CrossRef]
- Das, T.K.; Pradhan, S.; Chakrabarti, S.; Mondal, K.C.; Ghosh, K. Current Status of Probiotic and Related Health Benefits. Appl. Food Res. 2022, 2, 100185. [Google Scholar] [CrossRef]
- Borkent, J.; Ioannou, M.; Laman, J.D.; Haarman, B.C.M.; Sommer, I.E.C. Role of the Gut Microbiome in Three Major Psychiatric Disorders. Psychol. Med. 2022, 52, 1222–1242. [Google Scholar] [CrossRef] [PubMed]
- Loh, J.S.; Mak, W.Q.; Tan, L.K.S.; Ng, C.X.; Chan, H.H.; Yeow, S.H.; Foo, J.B.; Ong, Y.S.; How, C.W.; Khaw, K.Y. Microbiota–Gut–Brain Axis and Its Therapeutic Applications in Neurodegenerative Diseases. Sig. Transduct. Target Ther. 2024, 9, 1–53. [Google Scholar] [CrossRef]
- Al-Fakhrany, O.M.; Elekhnawy, E. Next-Generation Probiotics: The Upcoming Biotherapeutics. Mol. Biol. Rep. 2024, 51, 505. [Google Scholar] [CrossRef]
- Hasnain, M.A.; Kang, D.-K.; Moon, G.-S. Research Trends of next Generation Probiotics. Food Sci. Biotechnol. 2024, 33, 2111–2121. [Google Scholar] [CrossRef]
- Abouelela, M.E.; Helmy, Y.A. Next-Generation Probiotics as Novel Therapeutics for Improving Human Health: Current Trends and Future Perspectives. Microorganisms 2024, 12, 430. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- D’Argenio, V.; Salvatore, F. The Role of the Gut Microbiome in the Healthy Adult Status. Clin. Chim. Acta 2015, 451, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Wrigley-Carr, H.E.; van Dorst, J.M.; Ooi, C.Y. Intestinal Dysbiosis and Inflammation in Cystic Fibrosis Impacts Gut and Multi-Organ Axes. Med. Microecol. 2022, 13, 100057. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the Gut Microbiota in Disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Sig. Transduct. Target Ther. 2022, 7, 1–28. [Google Scholar] [CrossRef]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut Microbiota Metabolism and Interaction with Food Components. Int. J. Mol. Sci. 2020, 21, 3688. [Google Scholar] [CrossRef] [PubMed]
- Conlon, M.A.; Bird, A.R. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Ijssennagger, N.; van der Meer, R.; van Mil, S.W.C. Sulfide as a Mucus Barrier-Breaker in Inflammatory Bowel Dis-ease? Trends Mol. Med. 2016, 22, 190–199. [Google Scholar] [CrossRef]
- Kaliannan, K.; Wang, B.; Li, X.-Y.; Kim, K.-J.; Kang, J.X. A Host-Microbiome Interaction Mediates the Opposing Effects of Omega-6 and Omega-3 Fatty Acids on Metabolic Endotoxemia. Sci. Rep. 2015, 5, 11276. [Google Scholar] [CrossRef]
- Dahl, W.J.; Rivero Mendoza, D.; Lambert, J.M. Chapter Eight—Diet, Nutrients and the Microbiome. In Progress in Molecular Biology and Translational Science; The Microbiome in Health and Disease; Sun, J., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 171, pp. 237–263. [Google Scholar]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Wein-berger, A.; et al. Artificial Sweeteners Induce Glucose Intolerance by Altering the Gut Microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Reid, G. Probiotics: Definition, Scope and Mechanisms of Action. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 17–25. [Google Scholar] [CrossRef]
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.E.; Zimmermann, A.K.; Ouwehand, A.C. Contemporary Meta-Analysis of Short-Term Probiotic Consumption on Gastrointestinal Transit. World J. Gastroenterol. 2016, 22, 5122–5131. [Google Scholar] [CrossRef]
- Ng, Q.X.; Lim, Y.L.; Yaow, C.Y.L.; Ng, W.K.; Thumboo, J.; Liew, T.M. Effect of Probiotic Supplementation on Gut Microbiota in Patients with Major Depressive Disorders: A Systematic Review. Nutrients 2023, 15, 1351. [Google Scholar] [CrossRef] [PubMed]
- Klingbeil, E.; de La Serre, C.B. Microbiota Modulation by Eating Patterns and Diet Composition: Impact on Food In-take. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R1254–R1260. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Scott, S.M.; Whelan, K. Probiotics and Constipation: Mechanisms of Action, Evidence for Effectiveness and Utilisation by Patients and Healthcare Professionals. Proc. Nutr. Soc. 2020, 79, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Lin, T.-L.; Tsai, Y.-L.; Wu, T.-R.; Lai, W.-F.; Lu, C.-C.; Lai, H.-C. Next Generation Probiotics in Disease Amelioration. J. Food Drug Anal. 2019, 27, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Saarela, M.H. Safety Aspects of next Generation Probiotics. Curr. Opin. Food Sci. 2019, 30, 8–13. [Google Scholar] [CrossRef]
- Kubasova, T.; Seidlerova, Z.; Rychlik, I. Ecological Adaptations of Gut Microbiota Members and Their Consequences for Use as a New Generation of Probiotics. Int. J. Mol. Sci. 2021, 22, 5471. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Derrien, M.; Collado, M.C.; Ben-Amor, K.; Salminen, S.; de Vos, W.M. The Mucin Degrader Akkermansia Muciniphila Is an Abundant Resident of the Human Intestinal Tract. Appl. Environ. Microbiol. 2008, 74, 1646–1648. [Google Scholar] [CrossRef]
- Hagi, T.; Belzer, C. The Interaction of Akkermansia Muciniphila with Host-Derived Substances, Bacteria and Diets. Appl. Microbiol. Biotechnol. 2021, 105, 4833–4841. [Google Scholar] [CrossRef] [PubMed]
- van der Ark, K.C.H.; Aalvink, S.; Suarez-Diez, M.; Schaap, P.J.; de Vos, W.M.; Belzer, C. Model-Driven Design of a Minimal Medium for Akkermansia Muciniphila Confirms Mucus Adaptation. Microb. Biotechnol. 2018, 11, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia Muciniphila Is a Promising Probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef]
- Abuqwider, J.N.; Mauriello, G.; Altamimi, M. Akkermansia Muciniphila, a New Generation of Beneficial Microbiota in Modulating Obesity: A Systematic Review. Microorganisms 2021, 9, 1098. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia Muciniphila: Paradigm for next-Generation Beneficial Microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia Muciniphila and Its Role in Regulating Host Functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef]
- Miranda, V.C.; Souza, R.O.; Quintanilha, M.F.; Gallotti, B.; Assis, H.C.; Faria, A.M.C.; Nicoli, J.R.; Cara, D.C.; Mar-tins, F.S. A Next-Generation Bacteria (Akkermansia Muciniphila BAA-835) Presents Probiotic Potential Against Ovalbumin-Induced Food Allergy in Mice. Probiotics Antimicro. Prot. 2024, 16, 737–751. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Pasteurised Akkermansia Muciniphila as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06780. [Google Scholar] [CrossRef]
- Parrish, A.; Boudaud, M.; Grant, E.T.; Willieme, S.; Neumann, M.; Wolter, M.; Craig, S.Z.; De Sciscio, A.; Cosma, A.; Hunewald, O.; et al. Akkermansia Muciniphila Exacerbates Food Allergy in Fibre-Deprived Mice. Nat. Microbiol. 2023, 8, 1863–1879. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia Muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Machado, D.; Barbosa, J.C.; Domingos, M.; Almeida, D.; Andrade, J.C.; Freitas, A.C.; Gomes, A.M. Revealing Antimi-crobial Resistance Profile of the Novel Probiotic Candidate Faecalibacterium Prausnitzii DSM 17677. Int. J. Food Microbiol. 2022, 363, 109501. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhao, S.; Li, Y. Faecalibacterium Prausnitzii: A Next-Generation Probiotic in Gut Disease Improvement. Can. J. Infect. Dis. Med. Microbiol. 2021, 6666114. [Google Scholar] [CrossRef]
- Maioli, T.U.; Borras-Nogues, E.; Torres, L.; Barbosa, S.C.; Martins, V.D.; Langella, P.; Azevedo, V.A.; Chatel, J.-M. Possible Benefits of Faecalibacterium Prausnitzii for Obesity-Associated Gut Disorders. Front. Pharmacol. 2021, 12, 740636. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional Characterization of Novel Faecalibacterium Prausnitzii Strains Isolated from Healthy Volunteers: A Step Forward in the Use of F. Prausnitzii as a Next-Generation Probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar] [CrossRef]
- Ferreira-Halder, C.V.; de Faria, A.V.S.; Andrade, S.S. Action and Function of Faecalibacterium Prausnitzii in Health and Disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Khan, M.T.; van Dijl, J.M.; Harmsen, H.J.M. Antioxidants Keep the Potentially Probiotic but Highly Oxygen-Sensitive Human Gut Bacterium Faecalibacterium Prausnitzii Alive at Ambient Air. PLoS ONE 2014, 9, e96097. [Google Scholar] [CrossRef]
- Ryan, D.; Jenniches, L.; Reichardt, S.; Barquist, L.; Westermann, A.J. A High-Resolution Transcriptome Map Identifies Small RNA Regulation of Metabolism in the Gut Microbe Bacteroides Thetaiotaomicron. Nat. Commun. 2020, 11, 3557. [Google Scholar] [CrossRef]
- Ye, M.; Yu, J.; Shi, X.; Zhu, J.; Gao, X.; Liu, W. Polysaccharides Catabolism by the Human Gut Bacterium -Bacteroides Thetaiotaomicron: Advances and Perspectives. Crit. Rev. Food Sci. Nutr. 2021, 61, 3569–3588. [Google Scholar] [CrossRef]
- Xu, J.; Bjursell, M.K.; Himrod, J.; Deng, S.; Carmichael, L.K.; Chiang, H.C.; Hooper, L.V.; Gordon, J.I. A Genomic View of the Human-Bacteroides Thetaiotaomicron Symbiosis. Science 2003, 299, 2074–2076. [Google Scholar] [CrossRef]
- Xu, J.; Chiang, H.C.; Bjursell, M.K.; Gordon, J.I. Message from a Human Gut Symbiont: Sensitivity Is a Prerequisite for Sharing. Trends Microbiol. 2004, 12, 21–28. [Google Scholar] [CrossRef]
- Comstock, L.E.; Coyne, M.J. Bacteroides Thetaiotaomicron: A Dynamic, Niche-Adapted Human Symbiont. BioEssays 2003, 25, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Wrzosek, L.; Miquel, S.; Noordine, M.-L.; Bouet, S.; Chevalier-Curt, M.J.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides Thetaiotaomicron and Faecalibacterium Prausnitziiinfluence the Production of Mucus Glycans and the Development of Goblet Cells in the Colonic Epithelium of a Gnotobiotic Model Rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, J.; Zhang, H.; Lee, Y.-K.; Zhai, Q.; Chen, W. Roles of Intestinal Bacteroides in Human Health and Diseases. Crit. Rev. Food Sci. Nutr. 2021, 61, 3518–3536. [Google Scholar] [CrossRef] [PubMed]
- Durant, L.; Stentz, R.; Noble, A.; Brooks, J.; Gicheva, N.; Reddi, D.; O’Connor, M.J.; Hoyles, L.; McCartney, A.L.; Man, R.; et al. Bacteroides Thetaiotaomicron-Derived Outer Membrane Vesicles Promote Regulatory Dendritic Cell Responses in Health but Not in Inflammatory Bowel Disease. Microbiome 2020, 8, 88. [Google Scholar] [CrossRef]
- Fonseca, S.; Carvalho, A.L.; Miquel-Clopés, A.; Jones, E.J.; Juodeikis, R.; Stentz, R.; Carding, S.R. Extracellular Vesicles Produced by the Human Gut Commensal Bacterium Bacteroides Thetaiotaomicron Elicit Anti-Inflammatory Responses from Innate Immune Cells. Front. Microbiol. 2022, 13, 1050271. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kang, Y.; Huang, Y.; Xiao, Y.; Song, L.; Lu, S.; Ren, Z. A Strain of Bacteroides Thetaiotaomicron Attenuates Colonization of Clostridioides Difficile and Affects Intestinal Microbiota and Bile Acids Profile in a Mouse Model. Biomed. Pharmacother. 2021, 137, 111290. [Google Scholar] [CrossRef] [PubMed]
- Elahi, M.; Nakayama-Imaohji, H.; Hashimoto, M.; Tada, A.; Yamasaki, H.; Nagao, T.; Kuwahara, T. The Human Gut Microbe Bacteroides Thetaiotaomicron Suppresses Toxin Release from Clostridium Difficile by Inhibiting Autolysis. Antibiotics 2021, 10, 187. [Google Scholar] [CrossRef]
- Böger, M.; Hekelaar, J.; van Leeuwen, S.S.; Dijkhuizen, L.; Lammerts van Bueren, A. Structural and Functional Char-acterization of a Family GH53 β-1,4-Galactanase from Bacteroides Thetaiotaomicron That Facilitates Degradation of Prebiotic Galactooligosaccharides. J. Struct. Biol. 2019, 205, 1–10. [Google Scholar] [CrossRef]
- Lammerts van Bueren, A.; Mulder, M.; van Leeuwen, S.; Dijkhuizen, L. Prebiotic Galactooligosaccharides Activate Mucin and Pectic Galactan Utilization Pathways in the Human Gut Symbiont Bacteroides Thetaiotaomicron. Sci. Rep. 2017, 7, 40478. [Google Scholar] [CrossRef]
- Fernandez-Julia, P.J.; Munoz-Munoz, J.; van Sinderen, D. A Comprehensive Review on the Impact of β-Glucan Metabolism by Bacteroides and Bifidobacterium Species as Members of the Gut Microbiota. Int. J. Biol. Macromol. 2021, 181, 877–889. [Google Scholar] [CrossRef]
- Li, M.; Li, S.; Guo, X.; Guo, C.; Wang, Y.; Du, Z.; Zhang, Z.; Xie, C.; Ding, K. Discrete Genetic Loci in Human Gut Bacteroides Thetaiotaomicron Confer Pectin Metabolism. Carbohydr. Polym. 2021, 272, 118534. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhang, C.; Sifuentes-Dominguez, L.; Zarek, C.M.; Propheter, D.C.; Kuang, Z.; Wang, Y.; Pendse, M.; Ruhn, K.A.; Hassell, B.; et al. Small Proline-Rich Protein 2A Is a Gut Bactericidal Protein Deployed during Helminth Infection. Science 2021, 374, eabe6723. [Google Scholar] [CrossRef]
- Modasia, A.; Parker, A.; Jones, E.; Stentz, R.; Brion, A.; Goldson, A.; Defernez, M.; Wileman, T.; Ashley Blackshaw, L.; Carding, S.R. Regulation of Enteroendocrine Cell Networks by the Major Human Gut Symbiont Bacteroides Thetaiotaomicron. Front. Microbiol. 2020, 11, 575595. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.; Sanderson, I.R.; Muhammed, R.; Allen, S.; Tzivinikos, C.; Henderson, P.; Gervais, L.; Jeffery, I.B.; Mullins, D.P.; O’Herlihy, E.A.; et al. A Double-Blind, Placebo-Controlled Trial to Assess Safety and Tolerability of (Thetanix) Bacteroides Thetaiotaomicron in Adolescent Crohn’s Disease. Clin. Transl. Gastroenterol. 2021, 12, e00287. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Hayashi, N.; Lu, T.K. Engineering the Human Gut Commensal Bacteroides Thetaiotaomicron with Synthetic Biology. Curr. Opin. Chem. Biol. 2022, 70, 102178. [Google Scholar] [CrossRef]
- Curtis, M.M.; Hu, Z.; Klimko, C.; Narayanan, S.; Deberardinis, R.; Sperandio, V. The Gut Commensal Bacteroides Thetaiotaomicron Exacerbates Enteric Infection through Modification of the Metabolic Landscape. Cell Host Microbe 2014, 16, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Oba, S.; Sunagawa, T.; Tanihiro, R.; Awashima, K.; Sugiyama, H.; Odani, T.; Nakamura, Y.; Kondo, A.; Sasaki, D.; Sasaki, K. Prebiotic Effects of Yeast Mannan, Which Selectively Promotes Bacteroides Thetaiotaomicron and Bacteroides Ovatus in a Human Colonic Microbiota Model. Sci. Rep. 2020, 10, 17351. [Google Scholar] [CrossRef]
- Ignatyeva, O.; Tolyneva, D.; Kovalyov, A.; Matkava, L.; Terekhov, M.; Kashtanova, D.; Zagainova, A.; Ivanov, M.; Yudin, V.; Makarov, V.; et al. Christensenella Minuta, a New Candidate next-Generation Probiotic: Current Evidence and Future Trajectories. Front. Microbiol. 2024, 14, 1241259. [Google Scholar] [CrossRef]
- Ang, W.-S.; Law, J.W.-F.; Letchumanan, V.; Hong, K.W.; Wong, S.H.; Ab Mutalib, N.S.; Chan, K.-G.; Lee, L.-H.; Tan, L.T.-H. A Keystone Gut Bacterium Christensenella Minuta—A Potential Biotherapeutic Agent for Obesity and Associated Metabolic Diseases. Foods 2023, 12, 2485. [Google Scholar] [CrossRef]
- Pető, Á.; Kósa, D.; Szilvássy, Z.; Fehér, P.; Ujhelyi, Z.; Kovács, G.; Német, I.; Pócsi, I.; Bácskay, I. Scientific and Pharmaceutical Aspects of Christensenella Minuta, a Promising Next-Generation Probiotic. Fermentation 2023, 9, 767. [Google Scholar] [CrossRef]
- Xu, C.; Jiang, H.; Feng, L.-J.; Jiang, M.-Z.; Wang, Y.-L.; Liu, S.-J. Christensenella Minuta Interacts with Multiple Gut Bacteria. Front. Microbiol. 2024, 15, 1301073. [Google Scholar] [CrossRef] [PubMed]
- Mazier, W.; Le Corf, K.; Martinez, C.; Tudela, H.; Kissi, D.; Kropp, C.; Coubard, C.; Soto, M.; Elustondo, F.; Rawadi, G.; et al. A New Strain of Christensenella Minuta as a Potential Biotherapy for Obesity and Associated Metabolic Diseases. Cells 2021, 10, 823. [Google Scholar] [CrossRef]
- Liu, C.; Du, M.-X.; Xie, L.-S.; Wang, W.-Z.; Chen, B.-S.; Yun, C.-Y.; Sun, X.-W.; Luo, X.; Jiang, Y.; Wang, K.; et al. Gut Commensal Christensenella Minuta Modulates Host Metabolism via Acylated Secondary Bile Acids. Nat. Microbiol. 2024, 9, 434–450. [Google Scholar] [CrossRef]
- Kropp, C.; Le Corf, K.; Relizani, K.; Tambosco, K.; Martinez, C.; Chain, F.; Rawadi, G.; Langella, P.; Claus, S.P.; Mar-tin, R. The Keystone Commensal Bacterium Christensenella Minuta DSM 22607 Displays Anti-Inflammatory Properties Both in Vitro and in Vivo. Sci. Rep. 2021, 11, 11494. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Zheng, S.; Zheng, W.; Shi, C.; Ning, K.; Zhang, Q.; Xie, Y.; Xiang, H.; Xie, Q. Christensenella Regulated by Huang-Qi-Ling-Hua-San Is a Key Factor by Which to Improve Type 2 Diabetes. Front. Microbiol. 2022, 13, 1022403. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-W.; Huang, H.-J.; Wang, X.-M.; Wei, R.-Q.; Niu, H.-Y.; Chen, H.-Y.; Luo, M.; Abdugheni, R.; Wang, Y.-L.; Liu, F.-L.; et al. Christensenella Strain Resources, Genomic/Metabolomic Profiling, and Association with Host at Species Lev-el. Gut Microbes 2024, 16, 2347725. [Google Scholar] [CrossRef] [PubMed]
- Relizani, K.; Le Corf, K.; Kropp, C.; Martin-Rosique, R.; Kissi, D.; Déjean, G.; Bruno, L.; Martinez, C.; Rawadi, G.; Elustondo, F.; et al. Selection of a Novel Strain of Christensenella Minuta as a Future Biotherapy for Crohn’s Disease. Sci. Rep. 2022, 12, 6017. [Google Scholar] [CrossRef]
- Akbuğa-Schön, T.; Suzuki, T.A.; Jakob, D.; Vu, D.L.; Waters, J.L.; Ley, R.E. The Keystone Gut Species Christensenella Minuta Boosts Gut Microbial Biomass and Voluntary Physical Activity in Mice. mBio 2023, 15, e02836-23. [Google Scholar] [CrossRef]
- Kumari, M.; Singh, P.; Nataraj, B.H.; Kokkiligadda, A.; Naithani, H.; Ali, S.A.; Behare, P.V.; Nagpal, R. Fostering next-generation probiotics in human gut by targeted dietary modulation: An emerging perspective. Food Res. Int. 2021, 150, 110716. [Google Scholar] [CrossRef]
- Aeron, G.; Morya, S. Immobilization and Microencapsulationr. J. Adv. Res. Biotechnol. 2017, 2, 3520033. [Google Scholar]
- Kowalska, E.; Ziarno, M.; Ekielski, A.; Żelaziński, T. Materials Used for the Microencapsulation of Probiotic Bacteria in the Food Industry. Molecules 2022, 27, 3321. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, Q.; Ferguson, L.R.; Shu, Q.; Weir, I.; Garg, S. Development of a Novel Probiotic Delivery System Based on Microencapsulation with Protectants. Appl. Microbiol. Biotechnol. 2012, 93, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Panjanapongchai, S.; Chavapradit, C.; Anal, A.K. Microencapsulation of Probiotics and Its Potential Industrial Ap-plications. In Probiotics, Prebiotics and Synbiotics; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2022; pp. 213–232. ISBN 978-1-119-70216-0. [Google Scholar]
- Chen, Y.; Meenu, M.; Baojun, X. A Narrative Review on Microencapsulation of Obligate Anaerobe Probiotics Bifidobacterium, Akkermansia Muciniphila, and Faecalibacterium Prausnitzii. Food Rev. Int. 2022, 38, 373–402. [Google Scholar] [CrossRef]
- Yao, M.; Xie, J.; Du, H.; McClements, D.J.; Xiao, H.; Li, L. Progress in Microencapsulation of Probiotics: A Review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 857–874. [Google Scholar] [CrossRef] [PubMed]
- Druart, C.; Plovier, H.; Van Hul, M.; Brient, A.; Phipps, K.R.; de Vos, W.M.; Cani, P.D. Toxicological Safety Evalua-tion of Pasteurized Akkermansia Muciniphila. J. Appl. Toxicol. 2021, 41, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Dwibedi, C.; Sundh, D.; Pradhan, M.; Kraft, J.D.; Caesar, R.; Tremaroli, V.; Lorentzon, M.; Bäckhed, F. Synergy and Oxygen Adaptation for Development of Next-Generation Probiotics. Nature 2023, 620, 381–385. [Google Scholar] [CrossRef]
- Noyola, A.; Morgan-Sagastume, J.M.; López-Hernández, J.E. Treatment of Biogas Produced in Anaerobic Reactors for Domestic Wastewater: Odor Control and Energy/Resource Recovery. Rev. Environ. Sci. Biotechnol. 2006, 5, 93–114. [Google Scholar] [CrossRef]
- Hwang, O.H.; Cho, S.B.; Han, D.W.; Lee, S.R.; Kwag, J.H.; Park, S.K. Effect of Storage Period on the Changes of Odorous Compound Concentrations and Bacterial Ecology for Identifying the Cause of Odor Production from Pig Slurry. PLoS ONE 2016, 11, e0162714. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Mazier, W.; Frederic, E.; Rinaldi, L.; Rasmussen, S.; Rawadi, G. Safety evaluation of Christensenella minuta as a novel microbiome-based biotherapy to treat obesity. Obesity 2021, 29, 158. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).