1. Introduction
Biomass materials are promising carbon sources for the production of various value-added products. Among these products, lactic acid (LA) is an essential organic acid that provides a broad range of applications in food, cosmetic, pharmaceutical, and chemical conversion industries [
1]. It is also an important monomer for the production of polylactic acid (PLA) that has the potential to serve as promising biodegradable commodity plastics used for food packaging, textiles, trays, plates, and trash bags [
2]. Besides, this polymer has recently achieved a great interest as an additive manufacturing technology, for 3D printing, manufacturing of implants as biomedical components [
2,
3]. However, the high production cost of LA is hampering the commercial availability of PLA materials. The main expenses lie within the efficient fermentation processes (either microbial strain or utilized substrate) or the downstream processes of LA production [
4].
Industrially, LA can be manufactured by chemical or microbial fermentation routes. The chemical route is not preferred as it produces a racemic mixture of LA and involves the utilization of petrochemical resources [
5]. Therefore, the microbial route is preferred not only because of the high optical purity enantiomers of LA that can be achieved via fermentation, but also due to the utilization of renewable substrates that significantly improved environmental sustainability and serve as relatively cheap raw materials [
6].
The utilization of expensive raw materials has a significant effect on the overall LA production cost. Pure and crop sugars are utilized for production of LA; however, these sugars are considered as non-ideal substrates as compared to cellulosic materials, i.e., lignocellulosic and algal biomass act as the second and third-generation feedstock [
7]. Cellulosic materials are preferred due to their low price, abundance, sustainability, and does not compete with food crops [
8]. However, several technical limitations must be overcome to efficiently convert lignocellulosic biomass to LA. Of those, the direct utilization of these biomasses is difficult owing to their complex structure [
4]. The main carbohydrates present in microalgae and macroalgae are significantly different from those of plant biomass. The main structural polysaccharides are composed of cellulose and hemicellulose with varied ratios based on the source and type of biomass [
9,
10]. These biomasses are mostly pretreated and saccharified for the release of fermentable sugars that yield mixtures of hexoses and pentoses including glucose, cellobiose, and xylose with little amounts of arabinose, mannose, and oligosaccharides [
11,
12].
Lactic acid-producing bacteria are efficient consumers for glucose while xylose could be utilized by few strains [
4]. However, within fermentation of mixed sugars, most of these strains consume glucose preferentially to other sugars via a phenomenon known as carbon catabolite repression (CCR) resulting in extended fermentation time, wasting available substrates, lower LA titer, yield, and productivities [
11,
13]. Therefore, efficient utilization of all types of biomass-derived sugars in biomass hydrolysates is essential for economic conversion. In literature, very few wild type LA-producing strains could achieve efficient utilization of mixed-sugars without CCR achievable high LA titers, yields, and productivities [
11,
12,
14]. Various approaches have been done to rescue the CCR effect for the ultimate goal of relaxed control of sugar utilization and enhanced LA fermentations including genetic engineering and utilization of co-cultures [
15,
16].
Most reported studies on fermentative LA production from biomasses are focused on strain isolation and selection, mutagenesis or molecular breeding, technologies for pretreatment of cellulosic biomass, or optimization of LA fermentation from the hydrolysate regardless of their sugar composition [
1,
4,
6,
7,
8,
11,
12,
14,
15,
16,
17]. However, the present investigation focused on the effect of various sugar concentrations on LA production in order to select the best sugar composition alleviating CCR. This would greatly help to design the best biomass hydrolysate composition that achieve the highest sugar utilization and LA production in term of titer, yield, and productivity. This step is highly essential for further modification of the sacharification processes feasible for the microbial strain to establish an adaptive process maximizing LA fermentation from biomass substrates.
We have recently reported a new thermophilic and homofermentative xylose utilizing LA producer,
Bacillus coagulans Azu-10 that could ferment xylose efficiently at pH 7.0 and 50 °C [
17]. This strain showed the capability of utilizing various sugars existing in the biomass hydrolysates such as glucose, cellobiose, xylose and arabinose as a sole carbon source. Besides, it showed high resistance to most microbial inhibitors derived from biomass pretreatments including furans, carboxylic acids, and phenolic compounds [
17]. Therefore, the current investigation aimed to study the effect of various mixed-sugars derived from cellulosic biomass on the sugar utilization, CCR, and LA fermentation by the strain Azu-10 for the ultimate goal of establishing the optimally designed biomass hydrolysate suitable for efficient fermentation for economical LA industry-scale production.
4. Discussion
Cellulose biomasses are varied greatly in their sugar content and therefore, yield diverse ratios of sugar mixtures upon hydrolysis that should affect sugar utilization and LA fermentation efficiency owing to the carbon catabolite repression effect [
14,
15]. Biomass hydrolysates are composed of a mixture of sugars; therefore, co-utilization of various sugars is essential for economically feasible LA fermentation processes. However, LA fermentation by microbial producers is associated with several challenges including the un-consumption of existed sugars especially xylose. Otherwise, when sugars have been consumed, the utilization of preferred carbon and energy source such as glucose represses the consumption of alternative sugars that is known as carbon catabolite repression (CCR). Simultaneous utilization of biomass-derived mixed sugars (glucose, cellobiose, and xylose) results from partial or complete saccharification process has been demonstrated by means of few strains [
11,
12,
15]. Therefore, there is an urgent need to select the optimal substrates (sugars mixture) suitable for specific microbial strain for an effective LA fermentation process. This is known as “designed biomass” that can be used to establish the consolidated bioprocessing systems [
18].
We have recently isolated a robust LA-producer strain,
Bacillus coagulans Azu-10, capable of utilizing xylose homofermentatively and could tolerate several biomass-derived microbial inhibitors [
17]. This study firstly investigated the effect of different xylose concentrations on the LA fermentation in order to determine the optimal substrate concentration and the substrate inhibition. Strain Azu-10 has efficiently utilized up to 100 g/L xylose to LA at a yield of 1.0 g/g in a homofermentative pattern.
Numerous cellulosic biomasses have been widely used for LA fermentation, including woody biomasses, algal biomass, and grass biomass. In general, woody biomass contains higher cellulose content (40–55% cellulose, and 8–25% hemicellulose) than grass biomass (25–50% cellulose, 20–50% hemicellulose) [
19] while the structural polysaccharides of algae are different. These varying contents result in differential performances in enzymatic hydrolysis and LA fermentation. For example, 82% glucose was obtained from enzymatically hydrolyzed NaOH-pretreated Napier grass [
20], whereas 46.2% glucose was obtained from NaOH-pretreated rice straw [
21], 28% glucose from NaOH pretreated phragmites, and 33% glucose from switchgrass [
22].
Besides, one of the major challenges in fermentative LA production using cellulosic biomass is the release of microbial growth and LA production inhibitors such as furfural and 5-hydroxy methyl furfural, vanillin, acetic acid, and formic acid at concentrations ranging from 0.08 g/L to 5.3 g/L [
23]. Interestingly, strain Azu-10 achieved efficient LA production in the presence of different inhibitory compounds [
17]. This would overcome the extra required steps for substrate preparation including the removal of the inhibitors using chemicals, such as over-liming [
15,
24] or conversion of these inhibitory materials into less-toxic substances with a view to simplify the overall fermentation processes in an eco-friendly method. Some
B. coagulans strains have been reported to be resistant to certain levels of these inhibitors [
24,
25,
26,
27]; for example,
Bacillus sp. P38 could tolerate up to 6 g/L 2-furfural in corn stover hydrolysate [
27].
A recent study showed that the pretreatment methods affected the sugar ratio. Papayannakos et al. [
28] compared various chemical and physicochemical pretreatment methods (alkali, microwave-assisted acid, organosolv, hydrothermal treatment, and combination of the organosolv and hydrothermal treatment) using 15% solid loading of cotton stalks. Cellulose, hemicellulose, and lignin content of the pretreated materials were varied depending on the type of pretreatment applied, which might affect the fermentable sugar release. The order of pretreatments of cotton stalks was organosolv-hydrothermal, organosolv, alkali, microwave-assisted acid, and hydrothermal treatments with cellulose contents of 79.6, 77.0, 55.3, 48.6, and 47.3 g/100 g of pretreated cotton stalks, whereas the hemicellulose contents were 0.4, 1.4, 21.2, 9.6, and 3.9 g/100 g of pretreated cotton stalks, respectively [
28]. The cellulose hydrolysis yield of wheat straw was enhanced using a combination of steam explosion pretreatment and physical mill or chlorite pretreatment [
29].
The concentration of glucose/cellobiose/xylose mixture in cellulosic hydrolysate is greatly varied based on the hydrolysis process. Zhao et al., [
30] obtained cellobiose (7.29 g/L)/glucose (5.40 g/L)/xylose (10.2 g/L)/arabinose (1.89 g/L) mixture in separate hydrolysis process and cellobiose (5.31 g/L)/glucose (3.22 g/L)/xylose (10.3 g/L)/arabinose (1.91 g/L) mixture in repeated hydrolysis process from H
2SO
4 pretreated rice straw that hydrolysed by 2.6 U/g-substrate of cellulase while they obtained glucose (14.8 g/L)/xylose (9.7 g/L)/arabinose (1.31 g/L) mixture using 2.6 U/g-substrate of cellulase and 42.1 U/g-substrate of β-glucosidase. Moradi et al. [
31] obtained glucose (5.40 g/L)/xylose (1.2 g/L)/arabinose (0.3 g/L) mixture from 85% (
v/
v) H
3PO
4-pretreated rice straw that treated with 25 U/g-substrate of cellulase and 50.0 U/g-substrate of β-glucosidase while they obtained glucose (7.30 g/L)/xylose (2.20 g/L)/arabinose (0.400 g/L) mixture from 12% (
v/
v) NaOH-pretreated rice straw. On the other hand, Amiri et al. [
21] obtained glucose (15.0 g/L)/xylose (5.5 g/L)/arabinose (1.0 g/L) mixture from ethanol organosolve-pretreated rice straw using 25 U/g-substrate of cellulase and 40.0 U/g-substrate of β-glucosidase. He et al., [
20] obtained only glucose (34.8 g/L)/xylose (16.4 g/L) mixture from enzymatically hydrolyzed NaOH-pretreated Napier grass using 40 U/g-substrate of cellulase. Additionally, Gao and Rehmann [
32] obtained glucose (35.1 g/L)/xylose (20.1 g/L) mixture from enzymatically hydrolyzed NaOH-pretreated corncob using 15 U/g-substrate of cellulase. Amiri and Karimi [
33] obtained glucose (15.8 g/L)/xylose (6.47 g/L)/arabinose (0.89 g/L) mixture from ethanol organosolve-pretreated elm wood using 25 U/g-substrate of cellulase and 40.0 U/g-substrate of β-glucosidase while they obtained glucose (5.8 g/L)/xylose (1.07 g/L)/galactose (0.06 g/L)/mannose (2.30 g/L) mixture from pine using same hydrolysis process. Zhang et al., [
34] obtained cellobiose (1.2 g/L)/glucose (35.1 g/L)/xylose (3.0 g/L)/arabinose (10.1 g/L) mixture from Ca(OH)
2-pretreted corncob and enzymatically hydrolyzed using 48.0 U/g-substrate of cellulase and 20.0 U/g-substrate of β-glucosidase. Gao et al., [
22] obtained glucose (40.4 g/L)/xylose (16.5 g/L) mixture from NaOH-pretreated switchgrass (
Panicum virgatum) using 15 U/g-substrate of cellulase while they obtained glucose (35.7 g/L)/xylose (15.2 g/L) mixture from NaOH-pretreated phragmites (
Phragmites australis) using 15 U/g-substrate of cellulase. Zhang et al., [
35] obtained cellobiose (1.68 g/L)/glucose (39.5 g/L) from steam explosion-pretreated eucalyptus using 175 U/g-substrate of cellulose. Besides, various strategies can be used to control specific sugar concentrations to be used for specific fermentation processes including using a specific hydrolysis process, mixing different biomasses, and adding other resources. A semi-hydrolysis process using cellulase with low ß-glucosidase loading can control the disaccharide cellobiose concentration in a sugar mixture [
30]. The inhibition of cellulases by cellobiose (as a potent inhibitor) was controlled using multifunctional cellulases such as Umcel9y-1, Td2F2, and CoGH1A [
36]. High solid loading can be also used for generating a satisfactory amount of sugar. A starchy slurry was mixed with salix hydrolysate that improved the xylose consumption from 29 to 81% and enhanced the yield of fermentation end-product yield [
37].
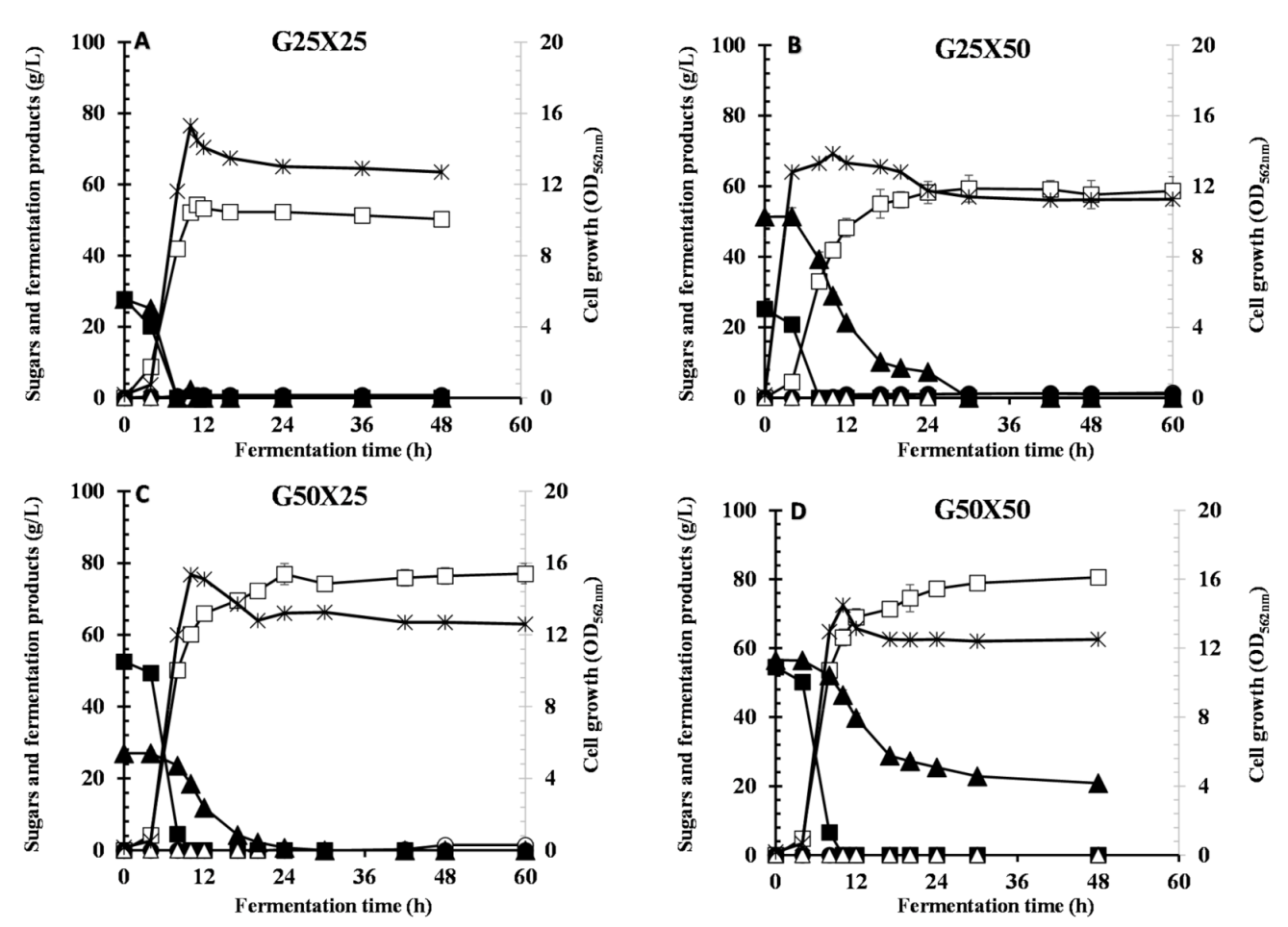
Another problem with the use of mixed sugars in cellulosic biomass is carbon catabolite repression (CCR). Interestingly, when strain Azu-10 was cultivated in different ratios of glucose–xylose mixtures, CCR was alleviated where both sugars were consumed simultaneously. The only limitation of G/X fermentation is the total substrate concentration greater than 75 g/L of mixed sugars. The intensities of CCR were previously reported as dependent on the concentration of the preferred sugar [
12]. In contrast, high LA yield was achieved when xylose concentration was 25 g/L while decreased yield was obtained at 50 g/L xylose. Our results suggest that the by-products (acetate, format, and ethanol) are not increased concerning the substrate conversion to cell mass rather than metabolic shift.
Lb. brevis ATCC 367 was reported to possess a relaxed CCR mechanism with glucose/xylose mixture; however, it produced LA at low yield (0.52 g/g) and high by-product formation [
15]. At concentrations higher than G25X25, glucose is preferentially consumed while xylose is consumed at a lower rate until completely utilized. At concentrations higher than 75 g/L (G50X50), only glucose was consumed while high residual xylose was left. This may be attributed to the limitation of some nutritional elements and/or accumulation of fermentation end-products post-glucose depletion, that slow and incomplete fermentation of non-glucose sugars [
23,
38]. Our results demonstrate that the initial concentration of glucose is a key factor determining xylose utilization for LA fermentation from mixed sugars using strain Azu-10, and G25X25 exhibited the best LA fermentation efficiency.
Enterococcus mundtii QU 25 achieved homo-LA production with low by-product formation and simultaneous glucose/xylose utilization by maintaining the concentrations of xylose over 10 g/L and glucose below 25 g/L [
14]. Besides high glucose concentration (50 g/L) did not repress enzymes responsible for xylose metabolism by strain Azu-10 where 76.9 g/L of LA could be produced from G50X25 after 24 h. In contrast, only 55.9 g/L of LA with a yield of 0.776 g/g was produced after 72 h using G50X25 by
E. mundtii QU 25 [
14]. On the other hand,
E. faecium QU 50 showed relaxed CCR in the utilization of G20X20 with the production of 41.0 g/L of LA at a yield of 1.01 g/g [
11].
B. coagulans JI12 utilized empty fruit bunches hydrolysate (containing glucose, 4.7 g/L; xylose, 48.8 g/L; and arabinose, 9.6 g/L) producing 59.2 g/L of LA at a yield of 0.97 g/g [
39]. Zhang et al., [
26] used membrane integrated repeated batch fermentation to mitigated the CCR feature of
B. coagulans IPE22 and the feedback inhibition during fermentation of wheat straw hydrolysates (29.72 g/L glucose, 24.69 g/L xylose and 5.14 g/L arabinose). They could achieve 54.5–56.4 g/L of LA with a yield of 0.94–96 g/g. A mixed sugar of sucrose, glucose, and/or fructose was also consumed for LA production without CCR by
E. faecalis RKY1 for LA production without CCR [
40]. Yoshida et al., [
41] integrated xylose-assimilating pathway into
Lb. plantarum and the strain produced homo LA from G75X25 without CCR, achieving 74.2 g/L of LA at a yield of 0.78 g/g. Similarly, Zhang et al., [
42] genetically modified
Lb. plantarum ldhL1-pCU-PxylAB to simultaneously utilize xylose/glucose mixture. The obtained strain achieved 20% more LA than the parent strain from cellulosic biomass. Lu et al., [
43] engineered
E. coli strain devoid of the glucose effect by the elimination of the ptsG gene encoding PTS enzyme for glucose transport, followed by metabolic evolution. The obtained strain co-utilized mixed sugars (G50X50) with the production of 83.0 g/L of LA [
43]. Other researchers used mixed culture to overcome CCR [
16,
35]. Co-cultivation of
Lb. plantarum ATCC 21,028 and
Lb. brevis enhanced LA production, yield (0.8 g/g-consumed sugar), and reduced by-products formation from mixed sugars. Additionally, co-cultivation of
Lb. rhamnosus and
Lb. brevis has been used for LA fermentation of xylose/glucose mixture and corn stover hydrolysate [
16].
In this study, the replacement of glucose by cellobiose achieved higher LA production titer than G/X sugar mixture without CCR. This might indicate that cellobiose has a lower effect than glucose on the enzymes responsible for xylose metabolism by Azu-10 strain. This might be attributed to the higher enzymatic activities of xylose isomerase and xylulokinase in cells grown in the presence of C/X sugar mixture than those obtained with cells grown in G/X sugar mixture as previously reported [
11]. This approach was also reported to rescue the apparent CCR by
E. mundtii QU 25 by enhancing xylose isomerase and xylulokinase activities in cellobiose/xylose grown cells. The strain produced 122 g/L of LA at a yield of 0.766 g/g from C100X60 compared to 71.2 g/L LA with a yield of 0.603 g/g obtained from G100X60 [
12]. Strain
E. faecium QU 50 exhibited no CCR during LA fermentation of C20X20 with the production of 43.6 g/L of LA at a yield of 1.05 g/g after 18 h [
11].
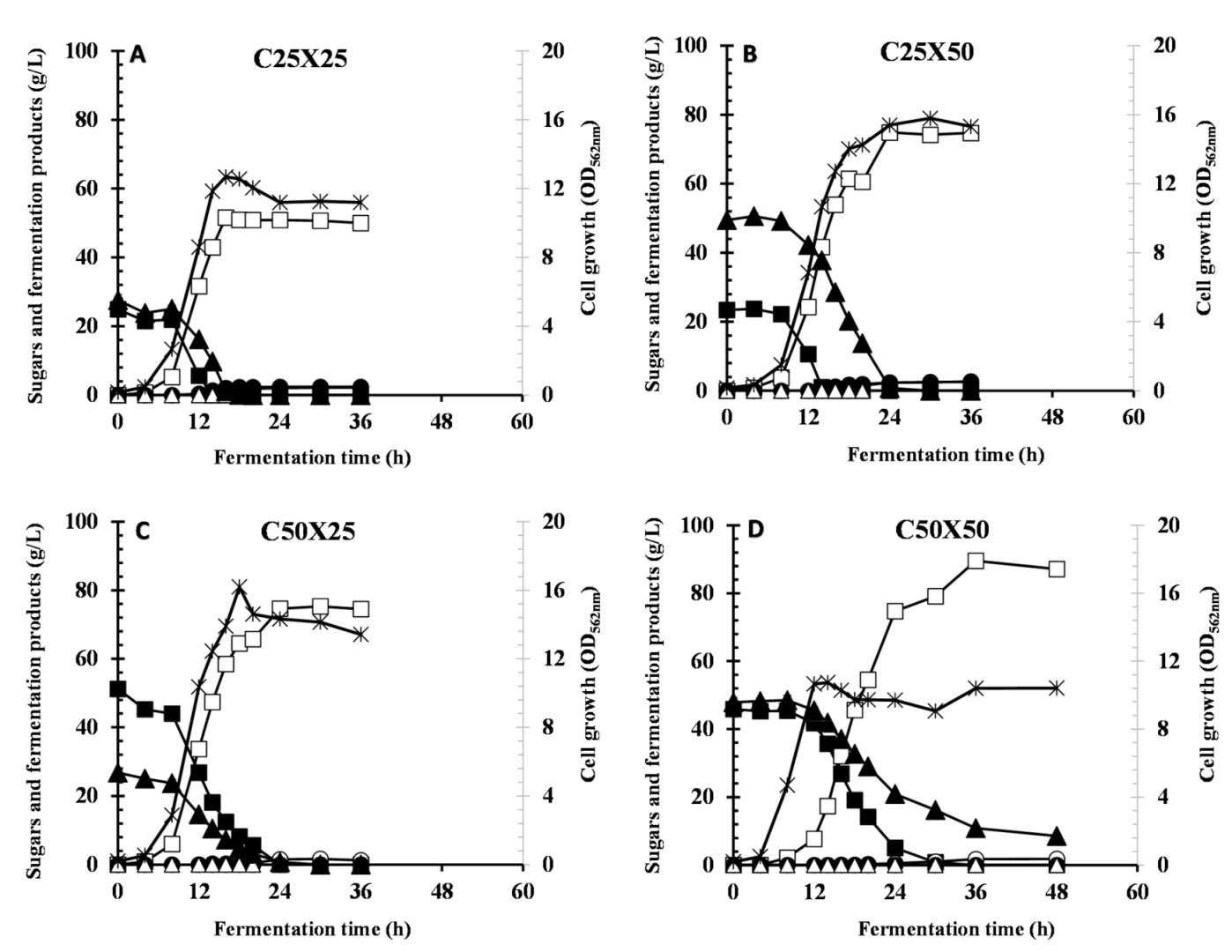
Cellulose fraction of cellulosic biomass can be hydrolyzed to glucose mainly in a complete hydrolysis process using cellulases and β-glucosidase or to a mixture of glucose and cellobiose in partial hydrolysis using cellulases only [
30]. To reveal the glucose effect of mixed-sugars released from cellulose saccharification of partial hydrolysis utilizing only cellulases without β-glucosidase (forming cellobiose and glucose). It was found that glucose concentration greater than 25 and total sugar concentration greater than 50 g/L are limiting factors for CCR by strain Azu-10 in G/C mixture. This might be attributed to the fact that glucose is a potent inhibitor for beta-glucosidase and cellulase activities [
44]. Supplementation of mannose and xylose during cellobiose hydrolysis did not show any inhibitory effects on beta-glucosidase activity as previously reported [
44]. Therefore, controlling the ratio of glucose to cellobiose is of great importance for effective LA fermentation by Azu-10. Our results indicated that the existence of cellobiose enhance glucose consumption but glucose retard cellobiose consumption at the tested concentrations. The results presented in this study indicate that initial glucose concentration >25.0 g/L is a limiting factor for efficient utilization of mixed glucose/cellobiose and high LA fermentation by
Bacillus coagulans Azu-10. Further studies should be conducted to improve the tolerance of β-glucosidase to glucose in order to maximize the sugar utilization and LA fermentation process.
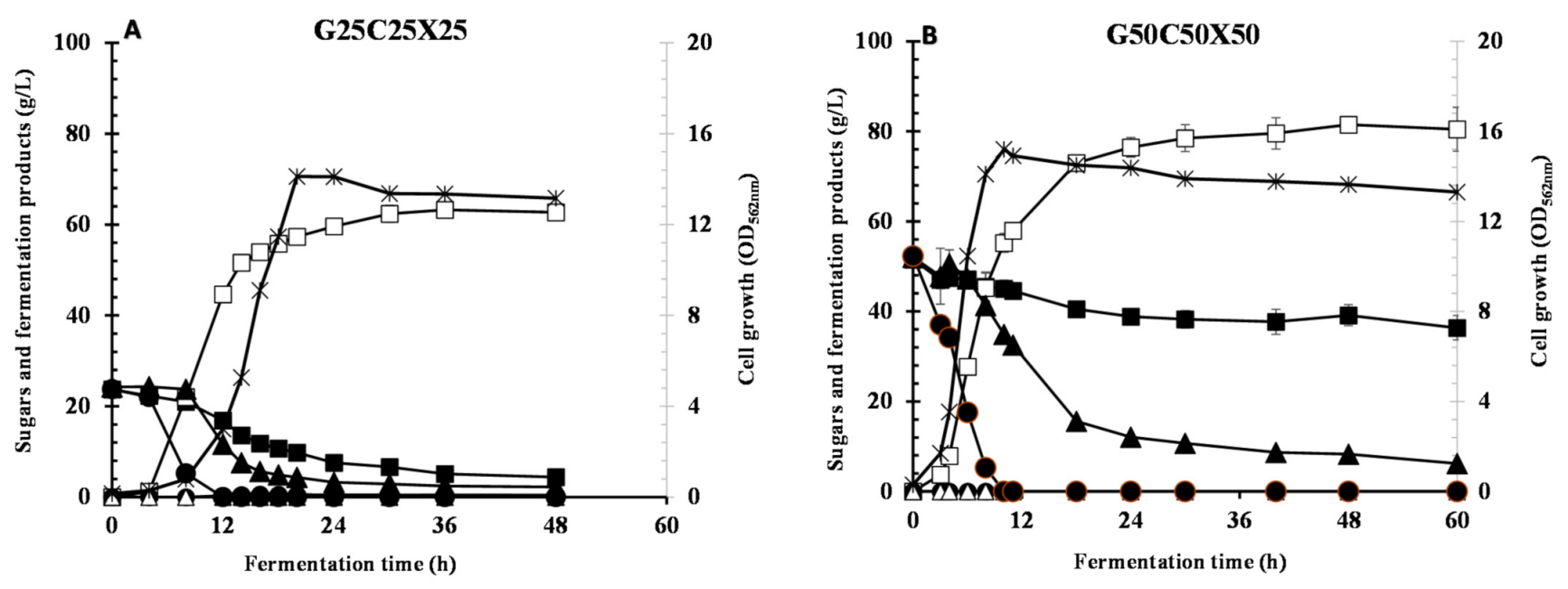
In this study, fermentation of glucose/cellobiose/xylose mixture was also found to be dependent on the glucose and total substrate concentrations. It is clear that glucose has more effect on cellobiose metabolism than xylose metabolism; therefore, Azu-10 preferentially utilized xylose next to glucose and then cellobiose. Our results indicated that the existence of cellobiose in the sugar mixture of lignocellulosic biomass hydrolysate should be controlled as it reduces the overall LA fermentation process. Thereby, it is recommended to use complete biomass hydrolysis by using cellulase(s) and β-glucosidase to yield high glucose concentration prior to LA fermentation by Azu-10. Mixing hemicellulose hydrolysate and cellulose hydrolysate would simplify and maximize LA fermentation by this strain. Taking together, fed-batch fermentation of glucose/xylose mixture resulted in homolactic fermentation with simultaneous sugar utilization. Further studies on the optimization of feeding solution and nutrient supplementation are required for achieving complete sugar conversion.
The homofermentative LA fermentation pattern and the obtained high yield by strain Azu-10 are mainly attributed to the metabolism of hexose by Embden–Meyerhof–Parnas pathway (EMP) and pentose sugars by the pentose phosphate pathway (PPP) [
45]. In the EMP pathway, hexose sugars are converted to pyruvate that is further reduced by LDH to produce LA [
8]. Each mole of glucose is converted to 2 mole LA; thus, the maximum theoretical yield of LA from glucose is 1 g/g. While in PPP, xylose is converted to xylulose xylulose 5-phosphate that is metabolized by transaldolase and transketolase to form GAP that is then converted to pyruvate and then to LA via EMP. In the PPP, every 3 moles of xylose are converted to 5 moles of LA, with a theoretical yield of 1 g/g [
8,
18,
46]. This characteristic is much better suited for increasing LA yield. De Clerck et al., [
47] investigated 15
B. coagulans strains and found that half of the strains can metabolize xylose. Besides, most of the reported
Bacillus strains achieved homolactic fermentation from xylose [
47,
48]. On the other hand, most of the reported LAB metabolize xylose hetermofermentatively via phosphoketolase (PK) pathway that produced LA and high byproducts (acetate or ethanol) with a theoretical LA yield of only 0.6 g/g (1 mol/mol) [
46]. To the best of our knowledge, only two LAB stains,
Enterococcus mundtii QU 25 and
E. faecium QU 50 were reported to metabolize xylose via PPP/EMP [
49,
50].


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}