

The Potential of Traditional ‘Gaja’ and New Breed Lines of Waxy, Blue and Purple Wheat in Wholemeal Flour Fermentation
 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
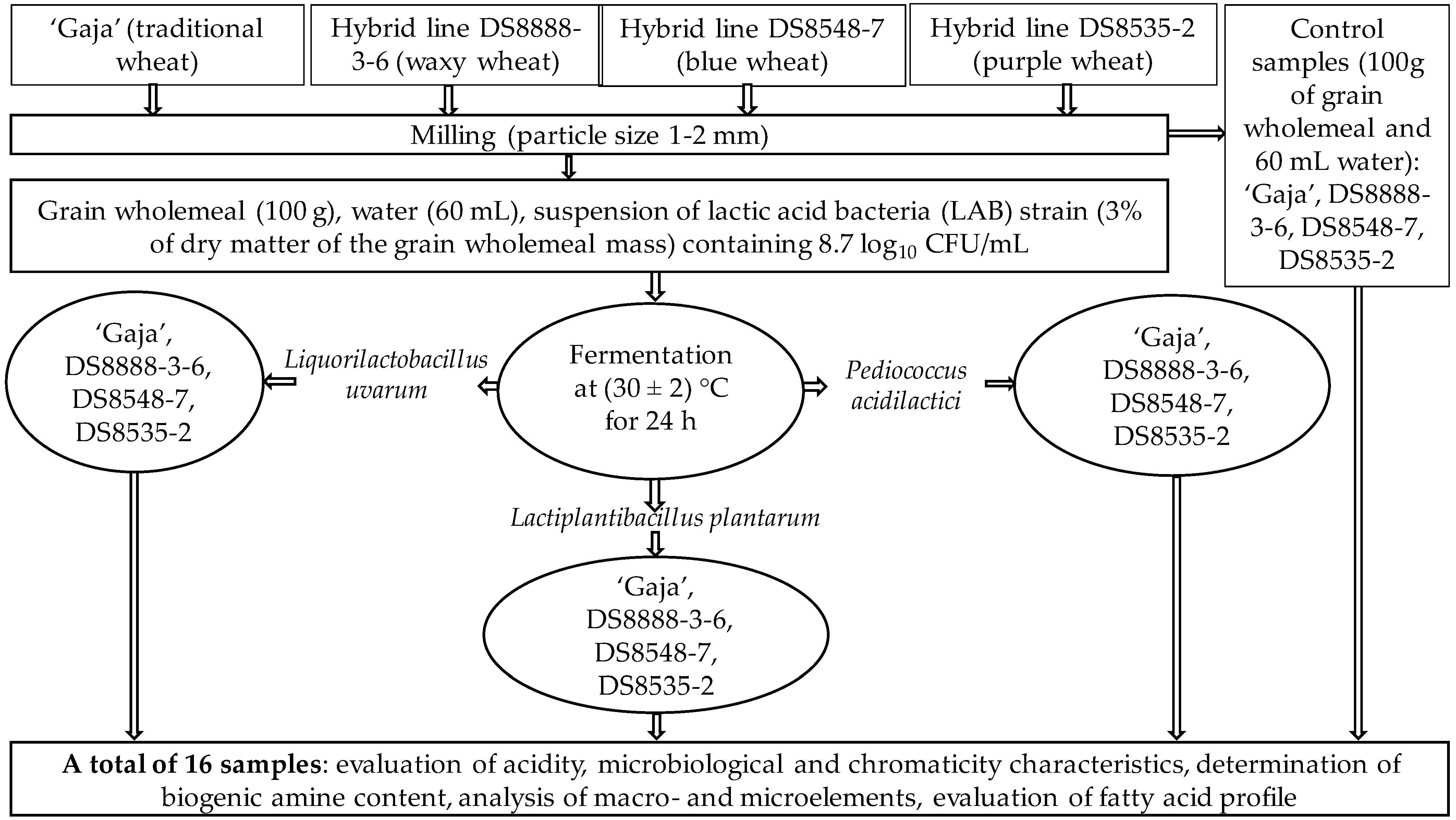
2. Materials and Methods
2.1. Wheat Varieties Used in the Experiments
2.2. Lactic Acid Bacteria Strains (LAB) Used in the Experiments and Wholemeal Wheat Flour Fermentation
2.3. Evaluation of Total Titratable Acidity (TTA), pH, Microbiological and Colour Characteristics of Wholemeal Wheat Flour Samples
2.4. Analysis of the Amino Acids (AA) and Gamma-Aminobutyric Acid (GABA) of Wholemeal Wheat Flour Samples
2.5. Analysis of the Biogenic Amine (BA) Content of Wholemeal Wheat Flour Samples
2.6. Analysis of Macro- and Microelements Content of Wholemeal Wheat Flour Samples
2.7. Fatty Acid (FA) Composition Analysis of Wholemeal Wheat Flour Samples
2.8. Statistical Analysis
3. Results and Discussion
3.1. Acidity, Microbiological and Chromaticity Characteristics of Wholemeal Wheat Flour Samples
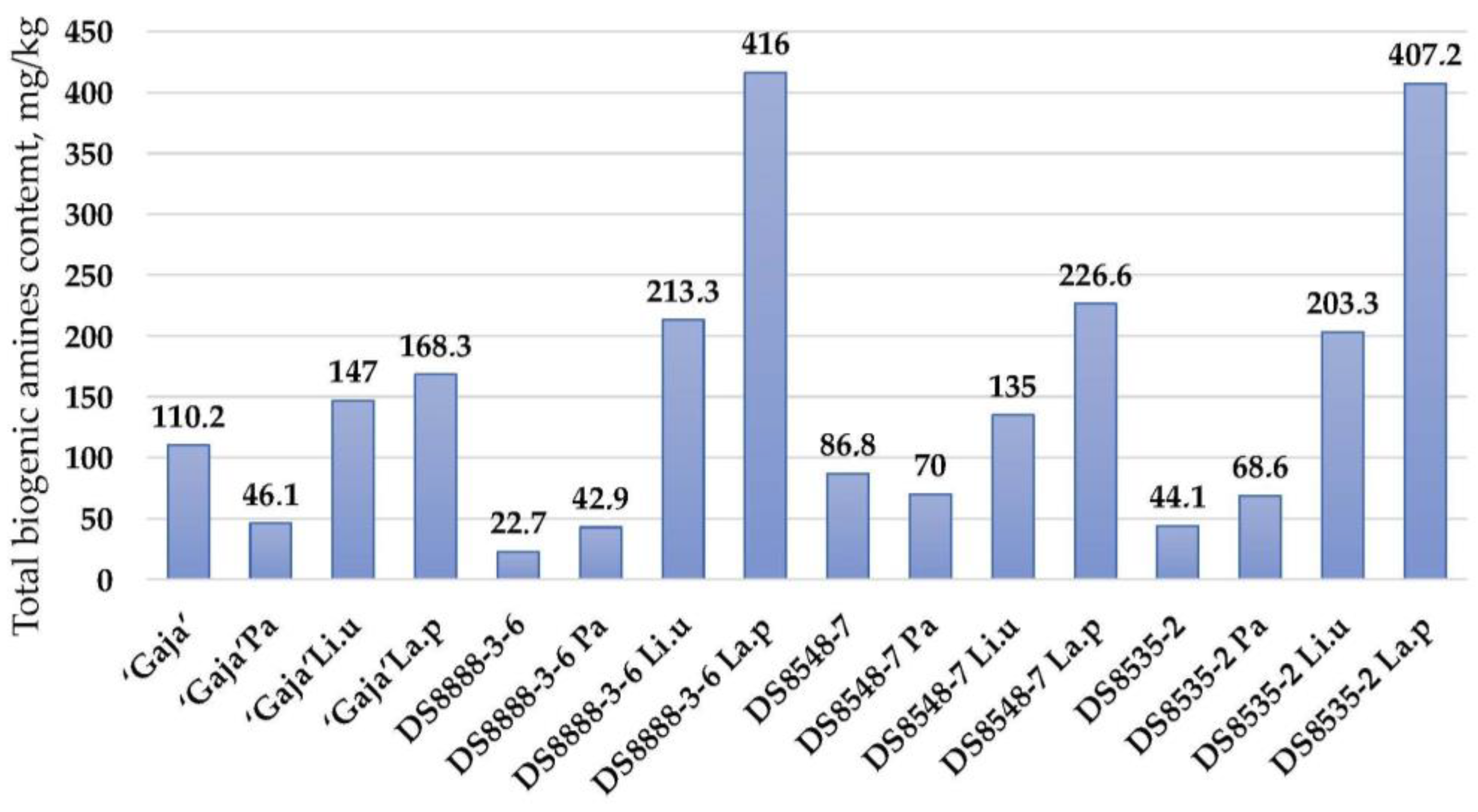
3.2. Amino Acids (AA), γ-Aminobutyric Acids (GABA) and Biogenic Amines (BA) of Wholemeal Wheat Flour Samples
3.3. Macro- and Microelements of Wholemeal Wheat Flour Samples
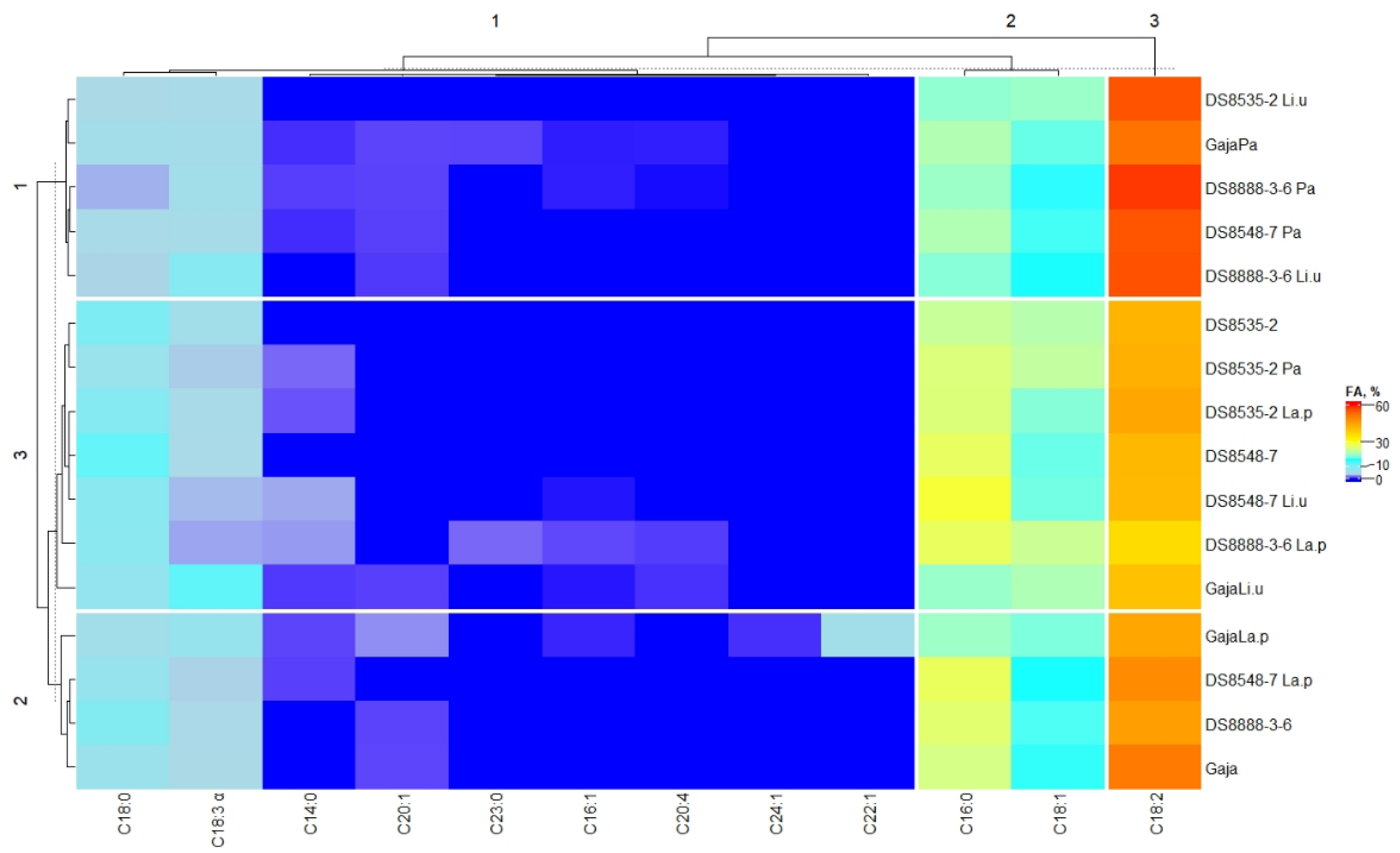
3.4. Fatty Acid Composition of Wholemeal Wheat Flour Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nakamura, T.; Yamamori, M.; Hirano, H.; Hidaka, S.; Nagamine, T. Production of Waxy (Amylose-Free) Wheats. Mol. Gen. Genet. MGG 1995, 248, 253–259. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Lai, H.-M. Noodle Quality Affected by Different Cereal Starches. J. Food Eng. 2010, 97, 135–143. [Google Scholar] [CrossRef]
- Baik, B.-K.; Lee, M.-R. Effects of Starch Amylose Content of Wheat on Textural Properties of White Salted Noodles. Cereal Chem. 2003, 80, 304–309. [Google Scholar] [CrossRef]
- Sasaki, T.; Yasui, T.; Matsuki, J. Effect of Amylose Content on Gelatinization, Retrogradation, and Pasting Properties of Starches from Waxy and Nonwaxy Wheat and Their F1 Seeds. Cereal Chem. 2000, 77, 58–63. [Google Scholar] [CrossRef]
- Blazek, J.; Copeland, L. Pasting and Swelling Properties of Wheat Flour and Starch in Relation to Amylose Content. Carbohydr. Polym. 2008, 71, 380–387. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Xu, C.; Zhou, X. Morphological Features and Physicochemical Properties of Waxy Wheat Starch. Int. J. Biol. Macromol. 2013, 62, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Sun, Y.; Yao, Y.; Li, H.; He, J. Study of Noodle Quality Based on Protein Properties of Three Wheat Varieties. J. Food Qual. 2022, 2022, e6383080. [Google Scholar] [CrossRef]
- Rausch, K.D.; Singh, V. Crops–Cereals. In Food Processing Principles and Applications; Wiley-Blackwell: New York, NY, USA, 2014; pp. 293–304. [Google Scholar]
- Song, J.M.; Liu, A.F.; You, M.S.; Li, B.Y.; Wu, X.Y.; Zhan, Z.D.; Liu, G.T. Effects of Waxy Flour Blending on Starch Pasting Properties and Noodle Quality of Non Waxy Flour. Sci. Agric. Sin. 2004, 37, 1838–1842. [Google Scholar]
- Aifeng, L.; Ran, H.; Dungong, C.; Haosheng, L.; Xinyou, C.; Jianmin, S.; Cheng, L.; Jun, G.; Faji, L.; Shengnan, Z.; et al. Applicability of Waxy Wheat Variety for Improving the Quality of Noodle and Steamed Bread. Int. J. Nutr. Food Sci. 2021, 10, 72. [Google Scholar] [CrossRef]
- Gupta, R.; Meghwal, M.; Prabhakar, P.K. Bioactive Compounds of Pigmented Wheat (Triticum Aestivum): Potential Benefits in Human Health. Trends Food Sci. Technol. 2021, 110, 240–252. [Google Scholar] [CrossRef]
- Lachman, J.; Martinek, P.; Kotíková, Z.; Orsák, M.; Šulc, M. Genetics and Chemistry of Pigments in Wheat Grain—A Review. J. Cereal Sci. 2017, 74, 145–154. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Zivcak, M.; Abdelly, C.; Cerda, A.; Brestic, M. Anthocyanins of Coloured Wheat Genotypes in Specific Response to Salstress. Molecules 2018, 23, 1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paznocht, L.; Kotíková, Z.; Šulc, M.; Lachman, J.; Orsák, M.; Eliášová, M.; Martinek, P. Free and Esterified Carotenoids in Pigmented Wheat, Tritordeum and Barley Grains. Food Chem. 2018, 240, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Kumari, A.; Chunduri, V.; Kaur, S.; Banda, J.; Goyal, A.; Garg, M. Anthocyanin Biofortified Black, Blue and Purple Wheat Exhibited Lower Amino Acid Cooking Losses than White Wheat. LWT 2022, 154, 112802. [Google Scholar] [CrossRef]
- Sharma, S.; Chunduri, V.; Kumar, A.; Kumar, R.; Khare, P.; Kondepudi, K.K.; Bishnoi, M.; Garg, M. Anthocyanin Bio-Fortified Colored Wheat: Nutritional and Functional Characterization. PLoS ONE 2018, 13, e0194367. [Google Scholar]
- Sharma, N.; Tiwari, V.; Vats, S.; Kumari, A.; Chunduri, V.; Kaur, S.; Kapoor, P.; Garg, M. Evaluation of Anthocyanin Content, Antioxidant Potential and Antimicrobial Activity of Black, Purple and Blue Colored Wheat Flour and Wheat-Grass Juice against Common Human Pathogens. Molecules 2020, 25, 5785. [Google Scholar] [CrossRef]
- Sharma, S.; Kapoor, P.; Kaur, S.; Kumari, A.; Sharma, N.; Kumar, A.; Chunduri, V.; Garg, M. Changing Nutrition Scenario: Colored Wheat–A New Perspective. In Physiological, Molecular, and Genetic Perspectives of Wheat Improvement; Springer: Berlin/Heidelberg, Germany, 2021; pp. 71–88. [Google Scholar]
- Sytar, O.; Bośko, P.; Živčák, M.; Brestic, M.; Smetanska, I. Bioactive Phytochemicals and Antioxidant Properties of the Grains and Sprouts of Colored Wheat Genotypes. Molecules 2018, 23, 2282. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Chen, Z.; Wei, Y. Measurement of Colour-Grained Wheat Nutrient Compounds and the Application of Combination Technology in Dough. J. Cereal Sci. 2018, 83, 63–67. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Pugajeva, I.; Krungleviciute, V.; Mayrhofer, S.; Domig, K. The Contribution of P. Acidilactici, L. Plantarum, and L. Curvatus Starters and L-(+)-Lactic Acid to the Acrylamide Content and Quality Parameters of Mixed Rye—Wheat Bread. LWT 2017, 80, 43–50. [Google Scholar] [CrossRef]
- Bartkiene, E.; Starkute, V.; Katuskevicius, K.; Laukyte, N.; Fomkinas, M.; Vysniauskas, E.; Kasciukaityte, P.; Radvilavicius, E.; Rokaite, S.; Medonas, D.; et al. The Contribution of Edible Cricket Flour to Quality Parameters and Sensory Characteristics of Wheat Bread. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and Their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2020, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badaras, S.; Ruzauskas, M.; Gruzauskas, R.; Zokaityte, E.; Starkute, V.; Klupsaite, D.; Mockus, E.; Klementaviciute, J.; Vadopalas, L.; Zokaityte, G.; et al. Different Creep Compound Feed Formulations for New Born Piglets: Influence on Growth Performance and Health Parameters. Front. Vet. Sci. 2022, 9, 971783. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Vizbickiene, D.; Bartkevics, V.; Pugajeva, I.; Krungleviciute, V.; Zadeike, D.; Zavistanaviciute, P.; Juodeikiene, G. Application of Pediococcus Acidilactici LUHS29 Immobilized in Apple Pomace Matrix for High Value Wheat-Barley Sourdough Bread. LWT-Food Sci. Technol. 2017, 83, 157–164. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Lele, V.; Pugajeva, I.; Zavistanaviciute, P.; Mickiene, R.; Zadeike, D.; Juodeikiene, G. A Concept of Mould Spoilage Prevention and Acrylamide Reduction in Wheat Bread: Application of Lactobacilli in Combination with a Cranberry Coating. Food Control 2018, 91, 284–293. [Google Scholar] [CrossRef]
- Cai, H.-L.; Zhu, R.-H.; Li, H.-D. Determination of Dansylated Monoamine and Amino Acid Neurotransmitters and Their Metabolites in Human Plasma by Liquid Chromatography–Electrospray Ionization Tandem Mass Spectrometry. Anal. Biochem. 2010, 396, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gigirey, B.; Vieites Baaptista de Sousa, J.M.; Villa, T.G.; Barros-Velazquez, J. Histamine and Cadaverine Production by Bacteria Isolated from Fresh and Frozen Albacore (Thunnus alalunga). J. Food Prot. 1999, 62, 933–939. [Google Scholar] [CrossRef]
- Evans, J.D. Straightforward Statistics for the Behavioral Sciences; Thomson Brooks/Cole Publishing Co: Belmont, CA, USA, 1996; pp. xxii, 600. ISBN 0-534-23100-4. [Google Scholar]
- Kati, K.; Kaisa, P.; Karin, A. Influence and Interactions of Processing Conditions and Starter Culture on Formation of Acids, Volatile Compounds, and Amino Acids in Wheat Sourdoughs. Cereal Chem. 2004, 81, 598–610. [Google Scholar] [CrossRef]
- Zhu, F. Anthocyanins in Cereals: Composition and Health Effects. Food Res. Int. 2018, 109, 232–249. [Google Scholar] [CrossRef]
- Enaru, B.; Drețcanu, G.; Pop, T.D.; Stǎnilǎ, A.; Diaconeasa, Z. Anthocyanins: Factors Affecting Their Stability and Degradation. Antioxidants 2021, 10, 1967. [Google Scholar] [CrossRef]
- Suo, B.; Chen, X.; Wang, Y. Recent Research Advances of Lactic Acid Bacteria in Sourdough: Origin, Diversity, and Function. Curr. Opin. Food Sci. 2021, 37, 66–75. [Google Scholar] [CrossRef]
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.-M.; Weckx, S. Microbial Ecology of Sourdough Fermentations: Diverse or Uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Tomičić, Z.; Pezo, L.; Spasevski, N.; Lazarević, J.; Cabarkapa, I.; Tomicic, R. Diversity of Amino Acids Composition in Cereals. Food Feed Res. 2022, 49, 11–22. [Google Scholar] [CrossRef]
- Ferreira, R.R.; Varisi, V.A.; Meinhardt, L.W.; Lea, P.J.; Azevedo, R.A. Are High-Lysine Cereal Crops Still a Challenge? Braz. J. Med. Biol. Res. 2005, 38, 985–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramadas, S.; Kumar, T.M.K.; Singh, G.P. Wheat Production in India: Trends and Prospects; IntechOpen: London, UK, 2019; ISBN 978-1-78985-450-3. [Google Scholar]
- Ufaz, S.; Galili, G. Improving the Content of Essential Amino Acids in Crop Plants: Goals and Opportunities. Plant Physiol. 2008, 147, 954–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caire-Juvera, G.; Vázquez-Ortíz, F.A.; Higuera-Ciapara, I.; Hernández, G. Composición de aminoácidos, calificación química y digestibilidad. Nutr. Hosp. 2013, 28, 365–371. [Google Scholar] [CrossRef]
- Siddiqi, R.A.; Singh, T.P.; Rani, M.; Sogi, D.S.; Bhat, M.A. Diversity in Grain, Flour, Amino Acid Composition, Protein Profiling, and Proportion of Total Flour Proteins of Different Wheat Cultivars of North India. Front. Nutr. 2020, 7, 141. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (Ed.) Dietary Protein Quality Evaluation in Human Nutrition: Report of an FAO Expert Consultation, 31 March–April 2011, Auckland, New Zealand; FAO food and nutrition paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 978-92-5-107417-6. [Google Scholar]
- Knežević, D.; Đukić, N.; Madic, M.; Paunović, A.; Zecevic, V. Comparison of Amino Acids Contents in Barley and Wheat. Res. J. Agric. Sci. 2007, 39, 71–76. [Google Scholar]
- Wilson, D.C.; Rafii, M.; Ball, R.O.; Pencharz, P.B. Threonine Requirement of Young Men Determined by Indicator Amino Acid Oxidation with Use of L-[1-13C]Phenylalanine. Am. J. Clin. Nutr. 2000, 71, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Zafar, S.; Naz, N.; Nazir, S.; Abbas, M.; Khan, A.M. Analysis of Selected Amino Acids in Different Varieties of Wheat Available in Punjab, Pakistan. Chromatogr. Res. Int. 2014, 2014, e867070. [Google Scholar] [CrossRef] [Green Version]
- Gheller, M.; Bender, E.; Thalacker-Mercer, A. Safety of Graded-Doses of Histidine in Healthy Adults (P08-062-19). Curr. Dev. Nutr. 2019, 3, nzz044.P08-062-19. [Google Scholar] [CrossRef] [Green Version]
- Moro, J.; Tomé, D.; Schmidely, P.; Demersay, T.-C.; Azzout-Marniche, D. Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species. Nutrients 2020, 12, 1414. [Google Scholar] [CrossRef] [PubMed]
- Laze, A.; Arapi, V.; Ceca, E.; Gusho, K.; Pezo, L.; Brahushi, F.; Kneževic, D. Chemical Composition and Amino Acid Content in Different Genotypes of Wheat Flour. Period. Polytech. Chem. Eng. 2019, 63, 618–628. [Google Scholar] [CrossRef]
- Jackman, S.R.; Witard, O.C.; Philp, A.; Wallis, G.A.; Baar, K.; Tipton, K.D. Branched-Chain Amino Acid Ingestion Stimulates Muscle Myofibrillar Protein Synthesis Following Resistance Exercise in Humans. Front. Physiol. 2017, 8, 390. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Tian, J.; Hao, Z.; Zhang, W. Protein Content and Amino Acid Composition in Grains of Wheat-Related Species. Agric. Sci. China 2008, 7, 272–279. [Google Scholar] [CrossRef]
- Bandegan, A.; Golian, A.; Kiarie, E.; Payne, R.L.; Crow, G.H.; Guenter, W.; Nyachoti, C.M. Standardized Ileal Amino Acid Digestibility in Wheat, Barley, Pea and Flaxseed for Broiler Chickens. Can. J. Anim. Sci. 2011, 91, 103–111. [Google Scholar] [CrossRef]
- Mayer, R.R.; Cherry, J.H.; Rhodes, D. Effects of Heat Shock on Amino Acid Metabolism of Cowpea Cells 1. Plant Physiol. 1990, 94, 796–810. [Google Scholar] [CrossRef] [Green Version]
- Aurisano, N.; Bertani, A.; Reggiani, R. Anaerobic Accumulation of 4-Aminobutyrate in Rice Seedlings; Causes and Significance. Phytochemistry 1995, 38, 1147–1150. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Lavermicocca, P.; De Vincenzi, M.; Giovannini, C.; Faccia, M.; Gobbetti, M. Proteolysis by Sourdough Lactic Acid Bacteria: Effects on Wheat Flour Protein Fractions and Gliadin Peptides Involved in Human Cereal Intolerance. Appl. Environ. Microbiol. 2002, 68, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Gänzle, M.G.; Loponen, J.; Gobbetti, M. Proteolysis in Sourdough Fermentations: Mechanisms and Potential for Improved Bread Quality. Trends Food Sci. Technol. 2008, 19, 513–521. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the Sourdough May Affect the Functional Features of Leavened Baked Goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.; Piggott, J.R. Flavour in Sourdough Breads: A Review. Trends Food Sci. Technol. 2006, 17, 557–566. [Google Scholar]
- Bhanwar, S.; Bamnia, M.; Ghosh, M.; Ganguli, A. Use of Lactococcus Lactis to Enrich Sourdough Bread with γ-Aminobutyric Acid. Int. J. Food Sci. Nutr. 2013, 64, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Diana, M.; Rafecas, M.; Quílez, J. Free Amino Acids, Acrylamide and Biogenic Amines in Gamma-Aminobutyric Acid Enriched Sourdough and Commercial Breads. J. Cereal Sci. 2014, 60, 639–644. [Google Scholar] [CrossRef]
- Okada, T.; Sugishita, T.; Murakami, T.; Murai, H.; Saikusa, T.; Horino, T.; Onoda, A.; Kajimoto, O.; Takahashi, R.; Takahashi, T. Effect of the Defatted Rice Germ Enriched with GABA for Sleeplessness, Depression, Autonomic Disorder by Oral Administration. Nippon Shokuhin Kagaku Kogaku Kaishi J. Jpn. Soc. Food Sci. Technol. 2000, 47, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Cassone, A.; Di Cagno, R.; Gobbetti, M. Synthesis of Angiotensin I-Converting Enzyme (ACE)-Inhibitory Peptides and γ-Aminobutyric Acid (GABA) during Sourdough Fermentation by Selected Lactic Acid Bacteria. J. Agric. Food Chem. 2008, 56, 6936–6943. [Google Scholar] [CrossRef]
- Villegas, J.M.; Brown, L.; de Giori, G.S.; Hebert, E.M. Optimization of Batch Culture Conditions for GABA Production by Lactobacillus Brevis CRL 1942, Isolated from Quinoa Sourdough. LWT-Food Sci. Technol. 2016, 67, 22–26. [Google Scholar] [CrossRef]
- Apetrei, I.M.; Apetrei, C. Amperometric Biosensor Based on Diamine Oxidase/Platinum Nanoparticles/Graphene/Chitosan Modified Screen-Printed Carbon Electrode for Histamine Detection. Sensors 2016, 16, 422. [Google Scholar] [CrossRef] [Green Version]
- Biji, K.B.; Ravishankar, C.N.; Venkateswarlu, R.; Mohan, C.O.; Gopal, T.K. Biogenic Amines in Seafood: A Review. J. Food Sci. Technol. 2016, 53, 2210–2218. [Google Scholar] [CrossRef]
- Askar, A.; Treptow, H. Biogene Amine in Lebensmitteln: Vorkommen, Bedeutung Und Bestimmung; E. Ulmer: Stuttgart, Germany, 1986; ISBN 3-8001-2132-8. [Google Scholar]
- Glória, M.B.A.; Tavares-Neto, J.; Labanca, R.A.; Carvalho, M.S. Influence of Cultivar and Germination on Bioactive Amines in Soybeans (Glycine max L. Merril). J. Agric. Food Chem. 2005, 53, 7480–7485. [Google Scholar] [CrossRef]
- Thamm, M.; Scholl, C.; Reim, T.; Grübel, K.; Möller, K.; Rössler, W.; Scheiner, R. Neuronal Distribution of Tyramine and the Tyramine Receptor AmTAR1 in the Honeybee Brain. J. Comp. Neurol. 2017, 525, 2615–2631. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, C.; Gómez-Tamayo, J.C.; Nebel, J.-C.; Pardo, L.; Gonzalez, A. Identifying Human Diamine Sensors for Death Related Putrescine and Cadaverine Molecules. PLoS Comput. Biol. 2018, 14, e1005945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardozo, M.; Lima, K.S.; Franca, T.C.; Lima, A.L.S. Aminas Biogênicas: Um Problema de Saúde Pública. Rev. Virtual Quím. 2013, 5, 149–168. [Google Scholar]
- Montet, D.; Ray, R.C. Fermented Foods, Part I: Biochemistry and Biotechnology; Taylor & Francis Group: New York, NY, USA, 2020; ISBN 978-0-367-73745-0. [Google Scholar]
- Silva, T.M.; Sabaini, P.S.; Evangelista, W.P.; Gloria, M.B.A. Occurrence of Histamine in Brazilian Fresh and Canned Tuna. Food Control 2011, 22, 323–327. [Google Scholar] [CrossRef]
- Oliveira, R.B.A.; Evangelista, W.P.; Sena, M.J.; Gloria, M.B.A. Tuna Fishing, Capture and Post-Capture Practices in the Northeast of Brazil and Their Effects on Histamine and Other Bioactive Amines. Food Control 2012, 25, 64–68. [Google Scholar] [CrossRef]
- Gomes, M.B.; Pires, B.A.D.; Fracalanzza, S.A.P.; Marin, V.A. The Risk of Biogenic Amines in Food. Cienc. Saude Colet. 2014, 19, 1123. [Google Scholar] [CrossRef] [Green Version]
- Gomes Müller, D.; Quadro Oreste, E.; Grazielle Heinemann, M.; Dias, D.; Kessler, F. Biogenic Amine Sensors and Its Building Materials: A Review. Eur. Polym. J. 2022, 175, 111221. [Google Scholar] [CrossRef]
- Dabrowski, W.M.; Sikorski, Z.E. Toxins in Food; CRC Press: Boca Raton, FL, USA, 2004; ISBN 0-203-50235-3. [Google Scholar]
- U.S. Department of Health and Human Services Food and Drug Administration. Fish and Fishery Products Hazards and Controls Guidance; US Department of Health and Human Services: Rockville, MD, USA, 2011.
- Santos, M.H.S. Biogenic Amines: Their Importance in Foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Jākobsone, I.; Kantāne, I.; Zute, S.; Jansone, I.; Bartkevičs, V. Macro-Elements and Trace Elements in Cereal Grains Cultivated in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2015, 69, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Underwood, B.A.; Smitasiri, S. Micronutrient Malnutrition: Policies and Programs for Control and Their Implications. Annu. Rev. Nutr. 1999, 19, 303–324. [Google Scholar] [CrossRef] [Green Version]
- Pietola, L.; Salo, T. Response of P, K, Mg and NO3-N Contents of Carrots to Irrigation, Soil Compaction, and Nitrogen Fertilisation. Agric. Food Sci. Finl. 2000, 9, 319–331. [Google Scholar] [CrossRef]
- Hattori, H.; Chino, M. Growth, Cadmium, and Zinc Contents of Wheat Grown on Various Soils Enriched with Cadmium and Zinc. In Plant Nutrition: Food Security and Sustainability of Agro-Ecosystems through Basic and Applied Research; Horst, W.J., Schenk, M.K., Bürkert, A., Claassen, N., Flessa, H., Frommer, W.B., Goldbach, H., Olfs, H.-W., Römheld, V., Sattelmacher, B., et al., Eds.; Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 2001; pp. 462–463. ISBN 978-0-306-47624-2. [Google Scholar]
- Kashian, S.; Fathivand, A.A. Estimated Daily Intake of Fe, Cu, Ca and Zn through Common Cereals in Tehran, Iran. Food Chem. 2015, 176, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhang, Z.; Xu, P.; Guo, Y. Analysis of Nutrient Composition of Purple Wheat. Cereal Res. Commun. 2012, 41, 293–303. [Google Scholar] [CrossRef]
- Kapur, D.; Nath Agarwal, K.; Kumari Agarwal, D. Nutritional Anemia and Its Control. Indian J. Pediatr. 2002, 69, 607–616. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for Micronutrients in Staple Food Crops from a Human Nutrition Perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef]
- Choi, Y.; Jeong, H.-S.; Lee, J. Antioxidant Activity of Methanolic Extracts from Some Grains Consumed in Korea. Food Chem. 2007, 103, 130–138. [Google Scholar] [CrossRef]
- Milder, I.E.; Arts, I.C.; van de Putte, B.; Venema, D.P.; Hollman, P.C. Lignan Contents of Dutch Plant Foods: A Database Including Lariciresinol, Pinoresinol, Secoisolariciresinol and Matairesinol. Br. J. Nutr. 2005, 93, 393–402. [Google Scholar] [CrossRef]
- Hosseinian, F.S.; Beta, T. Saskatoon and Wild Blueberries Have Higher Anthocyanin Contents than Other Manitoba Berries. J. Agric. Food Chem. 2007, 55, 10832–10838. [Google Scholar] [CrossRef]
- Li, W.; Beta, T. Flour and Bread from Black-, Purple-, and Blue-Colored Wheats. In Flour and Breads and Their Fortification in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2011; pp. 59–67. [Google Scholar]
- He, Y.Z.; Ning, J.F. Analysis of Nutrition Composition in the Special Purple Grain Wheat “Qinhei 1” Containing Rich Fe and Zn. J. Northwest Agric. For. Univ. 2003, 31, 87–90. [Google Scholar]
- Blanco, A. Quimica Biologica/Biological Chemistry; AbeBooks: Victoria, BC, Canada, 2006; ISBN 13: 9789500204224. [Google Scholar]
- Hurley, S.W.; Johnson, A.K. The Biopsychology of Salt Hunger and Sodium Deficiency. Pflüg. Arch.-Eur. J. Physiol. 2015, 467, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Moll, R.; Davis, B. Iron, Vitamin B12 and Folate. Medicine 2017, 45, 198–203. [Google Scholar] [CrossRef]
- Weaver, C.M.; Heaney, R.P. Calcium and Human Health, 1st ed.; Humana Press Inc: Totowa, NJ, USA, 2006. [Google Scholar]
- EFSA Dietary Reference Values|EFSA. Available online: https://www.efsa.europa.eu/en/topics/topic/dietary-reference-values (accessed on 27 September 2022).
- Fotedar, A.; Bhasin, J.S.; Chakravarty, A.; Kulkarni, A.; Bhalla, G.; Anwar, F.; Rao, S. Effectiveness of Iron-Fortified Infant Cereals on Hemoglobin Levels of Children Aged 12–24 Months: A Cross-Sectional Study from New Delhi, India. J. Fam. Med. Prim. Care 2018, 7, 77. [Google Scholar]
- Banerjee, P.; Bhattacharya, P. Investigating Cobalt in Soil-Plant-Animal-Human System: Dynamics, Impact and Management. J. Soil Sci. Plant Nutr. 2021, 21, 2339–2354. [Google Scholar] [CrossRef]
- Chasapis, C.T.; Ntoupa, P.-S.A.; Spiliopoulou, C.A.; Stefanidou, M.E. Recent Aspects of the Effects of Zinc on Human Health. Arch. Toxicol. 2020, 94, 1443–1460. [Google Scholar] [CrossRef]
- Rosentrater, K.A.; Evers, A.D. Chapter 4—Chemical Components and Nutrition. In Kent’s Technology of Cereals, 5th ed.; Rosentrater, K.A., Evers, A.D., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Cambridge, UK, 2018; pp. 267–368. ISBN 978-0-08-100529-3. [Google Scholar]
- Lafiandra, D.; Masci, S.; Sissons, M.; Dornez, E.; Delcour, J.; Courtin, C.; Caboni, M.F. Kernel Components of Technological Value. In Durum Wheat Chemistry and Technology; AACC International Press: Washington, DC, USA, 2012; pp. 85–124. [Google Scholar]
- Russo, G.L. Dietary N-6 and n-3 Polyunsaturated Fatty Acids: From Biochemistry to Clinical Implications in Cardiovascular Prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on Risk Factors, Molecular Pathways, and Clinical Events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [Green Version]
- Armanino, C.; De Acutis, R.; Rosa Festa, M. Wheat Lipids to Discriminate Species, Varieties, Geographical Origins and Crop Years. Anal. Chim. Acta 2002, 454, 315–326. [Google Scholar] [CrossRef]
- Beleggia, R.; Platani, C.; Nigro, F.; De Vita, P.; Cattivelli, L.; Papa, R. Effect of Genotype, Environment and Genotype-by-Environment Interaction on Metabolite Profiling in Durum Wheat (Triticum durum Desf.) Grain. J. Cereal Sci. 2013, 57, 183–192. [Google Scholar] [CrossRef]
- Nejadsadeghi, L.; Maali-Amiri, R.; Zeinali, H.; Ramezanpour, S.; Sadeghzade, B. Membrane Fatty Acid Compositions and Cold-Induced Responses in Tetraploid and Hexaploid Wheats. Mol. Biol. Rep. 2015, 42, 363–372. [Google Scholar] [CrossRef]
- Bottari, E.; De Acutis, R.; Festa, M.R. On the Lipid Constituents of Wheat of Different Species, Variety, Origin and Crop Year. Ann. Chim. 1999, 89, 849–862. [Google Scholar]
- Upchurch, R.G. Fatty Acid Unsaturation, Mobilization, and Regulation in the Response of Plants to Stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Rocha, J.M.; Kalo, P.J.; Malcata, F.X. Fatty acid composition of non-starch and starch neutral lipid extracts of portuguese sourdough bread. J. Am. Oil Chem. Soc. 2012, 89, 2025–2045. [Google Scholar] [CrossRef]
- Rocha, J.M.; Kalo, P.J.; Malcata, F.X. Composition of neutral lipid classes and content of fatty acids throughout sourdough breadmaking. Eur. J. Lipid Sci. Technol. 2012, 114, 294–305. [Google Scholar] [CrossRef]
- Rocha, J.M.; Kalo, P.J.; Malcata, F.X. Neutral lipids in free, bound and starch lipid extracts of flours, sourdough and portuguese sourdough bread, determined by NP-HPLC-ELSD. Cereal Chem. 2011, 88, 400–408. [Google Scholar] [CrossRef]
- Rocha, J.M.; Kalo, P.J.; Malcata, F.X. Neutral lipids in non-starch lipid and starch lipid extracts from portuguese sourdough bread. Eur. J. Lipid Sci. Technol. 2010, 112, 1138–1149. [Google Scholar] [CrossRef]
- Rocha, J.M.; Kalo PJOllilainen, V.; Malcata, F.X. Separation and identification of neutral cereal lipids by normal phase high-performance liquid chromatography, using evaporative light-scattering and electrospray mass spectrometry for detection. J. Chromatogr. A 2010, 1217, 3013–3025. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Wholemeal Wheat Samples | pH | TTA, °N | Colour Coordinates, NBS | LAB Viable Counts, log10 CFU/g | ||
|---|---|---|---|---|---|---|
| L* | a* | b* | ||||
| Traditional Wheat | ||||||
| ‘Gaja’ | 6.01 ± 0.01 d,A | 1.23 ± 0.11 a,B | 41.94 ± 0.12 a,A | 4.39 ± 0.09 a,C | 9.07 ± 0.11 a,A | 4.54 ± 0.32 a,A |
| ‘Gaja’ Pa | 4.44 ± 0.02 c,B | 4.91 ± 0.09 b,A | 44.11 ± 0.14 b,B | 4.86 ± 0.07 b,C | 9.16 ± 0.09 a,C | 8.48 ± 0.18 b,A,B |
| ‘Gaja’ Li.u | 4.37 ± 0.02 b,D | 4.90 ± 0.06 b,A | 52.11 ± 0.23 c,D | 4.88 ± 0.06 b,C | 16.01 ± 0.13 b,D | 8.52 ± 0.21 b,A,B |
| ‘Gaja’ La.p | 4.01 ± 0.01 a,C | 5.00 ± 0.05 b,A | 54.78 ± 0.15 d,C | 5.89 ± 0.08 c,C | 18.93 ± 0.14 c,D | 8.63 ± 0.14 b,A |
| Waxy Wheat | ||||||
| DS8888-3-6 | 6.11 ± 0.03 d,B | 1.20 ± 0.03 a,B | 52.29 ± 0.21 c,D | 3.06 ± 0.03 a,A | 14.78 ± 0.22 c,D | 4.19 ± 0.27 a,A |
| DS8888-3-6 Pa | 4.09 ± 0.02 b,A | 5.22 ± 0.05 b,B | 48.48 ± 0.35 a,C | 3.36 ± 0.05 b,A | 10.77 ± 0.18 a,D | 8.08 ± 0.14 b,A |
| DS8888-3-6 Li.u | 4.20 ± 0.01 c,B | 5.43 ± 0.04 c,B | 49.06 ± 0.21 b,C | 3.54 ± 0.06 c,B | 11.79 ± 0.21 b,C | 8.48 ± 0.11 c,A |
| DS8888-3-6 La.p | 3.99 ± 0.02 a,C | 5.61 ± 0.09 d,B | 52.29 ± 0.19 c,B | 3.06 ± 0.09 a,A | 14.78 ± 0.16 c,C | 8.61 ± 0.16 c,A |
| Blue Wheat | ||||||
| DS8548-7 | 6.18 ± 0.02 d,C | 1.02 ± 0.11 a,A | 49.51 ± 0.14 d,C | 4.10 ± 0.05 d,B | 12.68 ± 0.15 d,C | 4.69 ± 0.31 a,A |
| DS8548-7 Pa | 4.13 ± 0.02 b,A | 5.23 ± 0.06 b,B | 44.04 ± 0.23 b,B | 3.76 ± 0.11 c,B | 7.09 ± 0.09 a,B | 8.16 ± 0.23 b,A |
| DS8548-7 Li.u | 4.27 ± 0.02 c,C | 5.42 ± 0.10 c,B | 48.21 ± 0.17 c,B | 2.93 ± 0.09 a,A | 8.76 ± 0.08 c,B | 8.49 ± 0.16 b,A |
| DS8548-7 La.p | 3.55 ± 0.01 a,A | 6.10 ± 0.14 d,C | 41.69 ± 0.25 a,A | 3.34 ± 0.04 b,B | 7.65 ± 0.11 b,A | 8.94 ± 0.22 c,A |
| Purple Wheat | ||||||
| DS8535-2 | 5.98 ± 0.02 d,A | 1.00 ± 0.05 a,A | 45.14 ± 0.16 d,B | 5.93 ± 0.12 b,D | 11.16 ± 0.12 d,B | 4.41 ± 0.13 a,A |
| DS8535-2 Pa | 4.40 ± 0.02 c,B | 5.52 ± 0.07 b,C | 38.68 ± 0.22 b,A | 6.08 ± 0.08 b,D | 6.55 ± 0.07 b,A | 8.74 ± 0.19 b,B |
| DS8535-2 Li.u | 3.72 ± 0.03 b,A | 6.21 ± 0.05 c,C | 38.00 ± 0.13 a,A | 5.67 ± 0.06 a,D | 5.08 ± 0.08 a,A | 8.92 ± 0.21 b,B |
| DS8535-2 La.p | 3.61 ± 0.01 a,B | 6.30 ± 0.11 c,C | 41.98 ± 0.16 c,A | 5.95 ± 0.09 b,C | 8.10 ± 0.11 c,B | 8.99 ± 0.17 b,A |
| Wholemeal Wheat Samples | Concentration Essential of Amino Acids in Sample, µmol/g | ||||||
|---|---|---|---|---|---|---|---|
| Threonine | Methionine | Valine | Phenylalanine | Leucine/ Isoleucine | Lysine | Histidine | |
| Traditional Wheat | |||||||
| ‘Gaja’ | 0.460 ± 0.026 a,A | <0.020 | 0.385 ± 0.020 a,A | <0.020 | 0.185 ± 0.010 a,B,C | 0.360 ± 0.021 a,C | <0.020 |
| ‘Gaja’ Pa | 0.823 ± 0.077 d,C | 0.303 ± 0.014 c,C | 2.43 ± 0.16 d,C | 1.18 ± 0.07 b,C | 5.71 ± 0.22 d,C | 1.51 ± 0.14 d,B | 0.169 ± 0.007 b,C |
| ‘Gaja’ Li.u | 0.624 ± 0.034 b,A | 0.043 ± 0.003 a,A | 1.95 ± 0.10 c,C | 1.25 ± 0.10 b,C | 4.17 ± 0.25 c,B | 1.16 ± 0.05 c,B | <0.020 |
| ‘Gaja’ La.p | 0.780 ± 0.074 c,A | 0.146 ± 0.008 b,B | 1.21 ± 0.04 b,A | 0.767 ± 0.034 a,B | 2.95 ± 0.17 b,B | 0.839 ± 0.044 b,B | 0.083 ± 0.008 a,A |
| Waxy Wheat | |||||||
| DS8888-3-6 | 0.423 ± 0.040 a,A | <0.020 | 0.343 ± 0.034 a,A | <0.020 | 0.188 ± 0.012 a,C | 0.285 ± 0.020 a,B | <0.020 |
| DS8888-3-6 Pa | 0.768 ± 0.029 c,B | 0.117 ± 0.010 a,A | 2.01 ± 0.17 d,B | 0.813 ± 0.058 b,A | 4.67 ± 0.26 c,B | 1.07 ± 0.06 c,A | 0.024 ± 0.001 a,A |
| DS8888-3-6 Li.u | 0.591 ± 0.035 b,A | 0.146 ± 0.008 b,C | 1.65 ± 0.06 b,B | 0.788 ± 0.065 a,b,A | 3.53 ±0.32 b,A | 1.02 ± 0.04 c,B | 0.031 ± 0.003 b,A |
| DS8888-3-6 La.p | 0.715 ± 0.054 c,A | <0.020 | 1.99 ± 0.08 c,C | 0.695 ± 0.065 a,A | 3.01 ± 0.26 b,B,C | 0.583 ± 0.045 b,A | <0.020 |
| Blue Wheat | |||||||
| DS8548-7 | 0.420 ± 0.019 a,A | <0.020 | 0.422 ± 0.019 a,B | <0.020 | 0.162 ± 0.013 a,B | 0.224 ± 0.014 a,A | <0.020 |
| DS8548-7 Pa | 0.656 ± 0.024 c,A | 0.154 ± 0.009 b,B | 1.63 ± 0.13 c,A | 0.857 ± 0.078 b,A,B | 3.73 ± 0.14 d,A | 0.946 ± 0.056 d,A | 0.093 ± 0.004 b,B |
| DS8548-7 Li.u | 0.593 ± 0.044 b,A | 0.061 ± 0.005 a,B | 1.37 ± 0.09 b,A | 0.743 ± 0.074 a,b,A | 3.20 ± 0.11 c,A | 0.702 ± 0.059 c,A | 0.027 ± 0.002 a,A |
| DS8548-7 La.p | 0.685 ± 0.060 c,A | <0.020 | 1.49 ± 0.13 c,B | 0.641 ± 0.028 a,A | 2.47 ± 0.22 b,A | 0.671 ± 0.065 b,A | <0.020 |
| Purple Wheat | |||||||
| DS8535-2 | 0.440 ± 0.021 a,A | <0.020 | 0.430± 0.035 a,B | <0.020 | 0.103 ± 0.006 a,A | 0.321 ± 0.028 a,B,C | <0.020 |
| DS8535-2 Pa | 1.23 ± 0.09 d,D | 0.477 ± 0.015 c,D | 3.28 ± 0.31 c,D | 0.978 ± 0.083 b,B | 5.46 ± 0.20 c,C | 2.23 ± 0.20 d,C | 0.297 ± 0.013 c,D |
| DS8535-2 Li.u | 0.791 ± 0.025 b,B | 0.135 ± 0.007 b,C | 1.76 ± 0.08 b,B | 1.07 ± 0.10 b,B | 3.47 ± 0.30 b,A | 1.16 ± 0.06 c,B | 0.145 ± 0.005 b,B |
| DS8535-2 La.p | 0.909 ± 0.038 c,B | 0.100 ± 0.006 a,A | 1.87 ± 0.11 b,C | 0.853 ± 0.030 a,C | 3.54 ± 0.35 b,C | 0.629 ± 0.049 b,A | 0.116 ± 0.008 a,B |
| Wholemeal Wheat Samples | Concentration Nonessential of Amino Acids in Sample, µmol/g | GABA, µmol/g | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Arginine | Glutamine | Serine | Aspartic Acid | Glutamic Acid | Glycine | Alanine | Proline | Tyrosine | ||
| Traditional Wheat | ||||||||||
| ‘Gaja’ | 0.521 ± 0.018 b,C | 1.95 ± 0.14 c,A | 0.971 ± 0.061 a,A | 0.793 ± 0.068 b,A | 0.581 ± 0.057 b,D | 0.895 ± 0.040 a,C | 1.74 ± 0.16 a,A | 0.285 ± 0.015 a,A | <0.020 | 2.43 ± 0.13 a,C |
| ‘Gaja’ Pa | 0.221 ± 0.013 a,B | 3.07 ± 0.15 d,C | 1.47 ± 0.05 b,B | 1.38 ± 0.09 c,C | 0.780 ± 0.023 c,C | 1.84 ± 0.12 c,B | 5.98 ± 0.32 c,C | 1.55 ± 0.06 d,B | 0.246 ± 0.010 b,B | 5.32±0.30 b,C |
| ‘Gaja’ Li.u | 1.09 ± 0.10 c,C | 1.12±0.05 b,A | 1.16 ± 0.08 a,B | 0.686 ± 0.066 a,B | 0.477 ± 0.016 a,B | 1.56 ± 0.13 b,B | 4.28 ± 0.16 b,A | 1.31 ± 0.07 c,A | 0.279 ± 0.019 b,B | 6.37 ± 0.41 c,D |
| ‘Gaja’ La.p | 0.987 ± 0.071 c,D | 0.452 ± 0.023 a,B | 1.42 ± 0.13 a,b,B | 0.626 ± 0.020 a,C | 0.470 ± 0.038 a,C | 1.76 ± 0.11b, c,B | 4.54 ± 0.40 b,C | 0.972 ± 0.042 b,A | 0.055 ± 0.003 a,A | 6.51 ± 0.42 c,B |
| Waxy Wheat | ||||||||||
| DS8888-3-6 | 0.578 ± 0.047 b,C | 1.81 ± 0.10 a,A | 1.49 ± 0.11 a,b,B | 0.906 ± 0.036 c,B | 0.199 ± 0.019 a,A | 0.527a ± 0.033 A | 2.11 ± 0.10 a,B | 0.300 ± 0.025 a,A,B | <0.020 | 2.51 ± 0.09 a,C |
| DS8888-3-6 Pa | 0.236 ± 0.015 a,B | 2.66 ± 0.18 b,B | 1.35 ± 0.11 a,b,B | 1.15 ± 0.09 c,B | 0.585 ± 0.045 c,A | 1.39b ± 0.07 A | 4.27 ± 0.18 b,B | 1.39 ± 0.07 b,A | 0.246 ± 0.007 c,B | 4.53 ± 0.39 b,B |
| DS8888-3-6 Li.u | 0.722 ± 0.054 c,B | 1.75 ± 0.11 a,B | 1.26 ± 0.09 a,B | 0.546 ± 0.051 b,A | 0.409 ± 0.030 b,A | 1.33b ± 0.07 A | 4.07 ± 0.31 b,A | 1.35 ± 0.05 b,A | 0.225 ± 0.007 b,A | 4.87 ± 0.17 b,C |
| DS8888-3-6 La.p | 0.227 ± 0.007 a,B | 1.97 ± 0.18 a,C | 1.30 ± 0.11 a,b,B | 0.425 ± 0.027 a,B | 0.392 ± 0.033 b,B | 2.06 ± 0.17 c,C | 4.51 ± 0.31 b,C | 2.22 ± 0.17 c,C | 0.191 ± 0.007 a,C | 7.60 ± 0.24 c,C |
| Blue Wheat | ||||||||||
| DS8548-7 | 0.370 ± 0.033 b,A | 2.52 ± 0.23 b,C | 1.03 ± 0.09 b,A | 0.893 ± 0.057 c,B | 0.368 ± 0.024 b,C | 0.494 ± 0.020 a,A | 3.03 ± 0.25 a,C | 0.330 ± 0.028 a,B | <0.020 | 2.27 ± 0.21 a,B |
| DS8548-7 Pa | 0.337 ± 0.011 a,C | 2.32 ± 0.22 b,A | 0.919 ± 0.031 a,A | 0.954 ± 0.043 d,A | 0.677 ± 0.048 c,B | 1.25 ± 0.12 b,A | 3.56 ± 0.21 a,b,A | 1.26 ± 0.08 b,A | 0.198 ± 0.008 b,A | 3.25 ± 0.29 b,A |
| DS8548-7 Li.u | 0.352 ± 0.035 a,b,A | 2.37 ± 0.08 b,C | 0.960 ± 0.038 a, b,A | 0.782 ± 0.07 b,C | 0.621 ± 0.026 c,D | 1.18 ± 0.10 b,A | 3.73 ± 0.37 b,A | 1.33 ± 0.07 b,A | 0.210 ± 0.010 b,A | 3.35 ± 0.13 b,A |
| DS8548-7 La.p | 0.846 ± 0.048 c,C | 0.134 ± 0.004 a,A | 1.13 ± 0.05 b,A | 0.147 ± 0.013 a,A | 0.154 ± 0.009 a,A | 1.30 ± 0.047 b,A | 4.15 ± 0.13 c,B | 1.65 ± 0.15 c,B | 0.108 ± 0.009 a,B | 6.06 ± 0.47 c,B |
| Purple Wheat | ||||||||||
| DS8535-2 | 0.475 ± 0.041 c,B | 2.01 ± 0.11 a,B | 1.05 ± 0.06 a,A | 0.739 ± 0.036 a,A | 0.245 ± 0.008 a,B | 0.663 ± 0.054 a,B | 1.72 ± 0.06 a,A | 0.336 ± 0.031 a,B | <0.020 | 1.56 ± 0.14 a,A |
| DS8535-2 Pa | 0.192 ± 0.007 b,A | 4.31 ± 0.37 d,D | 1.85 ± 0.11 c,C | 1.72 ± 0.09 c,D | 1.22 ± 0.114 d,D | 2.41 ± 0.12 c,C | 8.08 ± 0.31 d,D | 2.16 ± 0.13 c,C | 0.243 ± 0.023 a,B | 5.61 ± 0.56 d,C |
| DS8535-2 Li.u | 1.21 ± 0.05 d,C | 3.26 ± 0.30 c,D | 1.22 ± 0.06 b,B | 0.894 ± 0.059 b,D | 0.522 ± 0.021 b,C | 1.72 ± 0.16 b,B | 4.28 ± 0.16 c,A | 1.72 ± 0.15 b,B | 0.268 ± 0.008 b,B | 3.97 ± 0.34 c,B |
| DS8535-2 La.p | 0.105 ± 0.007 a,A | 2.77 ± 0.17 b,D | 1.10 ± 0.10 a,b,A | 0.849 ± 0.062 b,D | 0.783 ± 0.055 c,D | 1.78 ± 0.11 b,B | 2.68 ± 0.21 b,A | 1.58 ± 0.15 b,B | 0.227 ± 0.009 a,D | 2.82 ± 0.24 b,A |
| Wholemeal Wheat Samples | Biogenic Amines, mg/kg | |||||||
|---|---|---|---|---|---|---|---|---|
| Tryptamine | Phenyl- ethylamine | Putrescine | Cadaverine | Histamine | Tyramine | Spermidine | Spermine | |
| Traditional Wheat | ||||||||
| ‘Gaja’ | <0.1 | 58.5 ± 3.7 b,B | 19.9 ± 1.1 | <0.1 | <0.1 | <0.1 | 31.8 ± 2.9 B | <0.1 |
| ‘Gaja’ Pa | <0.1 | 46.1 ± 2.9 a,A | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| ‘Gaja’ Li.u | <0.1 | 52.6 ± 4.1 a,b,B | <0.1 | 94.4 ± 5.4 a,C | <0.1 | <0.1 | <0.1 | <0.1 |
| ‘Gaja’ La.p | <0.1 | 52.0 ± 4.8 a,b,C | <0.1 | 116.3 ± 8.1 b,B | <0.1 | <0.1 | <0.1 | <0.1 |
| Waxy Wheat | ||||||||
| DS8888-3-6 | <0.1 | 57.3 ± 4.5 b,B | <0.1 | <0.1 | <0.1 | <0.1 | 22.7 ± 1.6 A | <0.1 |
| DS8888-3-6 Pa | <0.1 | 42.9 ± 3.8 a,A | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| DS8888-3-6 Li.u | <0.1 | 45.9 ± 4.1 a,A,B | 103.8 ± 7.2a | 63.6 ± 4.9 a,B | <0.1 | <0.1 | <0.1 | <0.1 |
| DS8888-3-6 La.p | <0.1 | 41.8 ± 3.5 a,B | 226.2 ± 14.1 b,C | 127.9 ± 8.2 b,B | 20.1 ± 1.7 A | <0.1 | <0.1 | <0.1 |
| Blue Wheat | ||||||||
| DS8548-7 | <0.1 | 55.9 ± 4.8 b,B | <0.1 | <0.1 | <0.1 | <0.1 | 30.9 ± 2.5 a,B | <0.1 |
| DS8548-7 Pa | <0.1 | 40.5 ± 3.1 a,A | <0.1 | <0.1 | <0.1 | <0.1 | 29.5 ± 2.2 a,B | <0.1 |
| DS8548-7 Li.u | <0.1 | 42.0 ± 2.9 a,A | <0.1 | 52.4 ± 4.7 a,A | <0.1 | <0.1 | 40.6 ± 3.1 b,B | <0.1 |
| DS8548-7 La.p | <0.1 | 39.7 ± 3.0 a,B | 27.2 ± 1.7 A | 84.1 ± 5.3 b,A | 49.9 ± 3.8 B | <0.1 | 25.7 ± 1.3 a,A | <0.1 |
| Purple Wheat | ||||||||
| DS8535-2 | <0.1 | 44.1 ± 3.5 b,A | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| DS8535-2 Pa | <0.1 | 44.1 ± 4.2 b,A | <0.1 | <0.1 | <0.1 | <0.1 | 24.5 ± 2.2 a,A | <0.1 |
| DS8535-2 Li.u | <0.1 | 50.9 ± 4.7 b,B | <0.1 | 122.6 ± 8.4 a,D | <0.1 | <0.1 | 29.8 ± 2.8 b,A | <0.1 |
| DS8535-2 La.p | <0.1 | 30.9 ± 2.9 a,A | 87.3 ± 6.3 B | 115.1 ± 9.1 a,B | 66.3 ± 5.1 C | 35.6 ± 3.2 | 51.8 ± 4.9 c,B | 20.2 ± 1.3 |
| Wholemeal Wheat Samples | Macroelements, Dry Matter (d.m.) | |||
|---|---|---|---|---|
| Na, g/100g | Mg, g/kg | K, g/kg | Ca, g/kg | |
| Traditional Wheat | ||||
| ‘Gaja’ | <0.002 | 0.682 ± 0.053 | 2.54 ± 0.22 a,B | 0.210 ± 0.019 a,B |
| ‘Gaja’ Pa | 0.010 ± 0.002 a,C | 0.638 ± 0.051 a,B | 2.32 ± 0.19 a,C | 0.199 ± 0.018 a,B |
| ‘Gaja’ Li.u | 0.008 ± 0.001 a,B | 0.669 ± 0.042 a,C | 2.38 ± 0.13 a,B | 0.192 ± 0.019 a,A,B |
| ‘Gaja’ La.p | 0.007 ± 0.002 a,A | 0.641 ± 0.050 a,C | 2.37 ± 0.21 a,B | 0.189 ± 0.020 a,A |
| Waxy Wheat | ||||
| DS8888-3-6 | <0.002 | 0.448 ± 0.045 a,A | 1.98 ± 0.17 a,A | 0.345 ± 0.031 a,C |
| DS8888-3-6 Pa | 0.013 ± 0.001 a,C | 0.501 ± 0.041 a,A | 2.38 ± 0.23 a,C | 0.310 ± 0.026 a,C |
| DS8888-3-6 Li.u | 0.011 ± 0.002 a,B | 0.438 ± 0.040 a,A | 2.20 ± 0.18 a,B | 0.321 ± 0.027 a,C |
| DS8888-3-6 La.p | 0.014 ± 0.001 a,B | 0.509 ± 0.051 a,A,B | 2.37 ± 0.23 a,B | 0.322 ± 0.030 a,C |
| Blue Wheat | ||||
| DS8548-7 | <0.002 | 0.510 ± 0.048 a,A | 1.93 ± 0.16 a,A | 0.213 ± 0.018 a,B |
| DS8548-7 Pa | 0.003 ± 0.001 a,A | 0.511 ± 0.027 a,A | 1.84 ± 0.11 a,A | 0.205 ± 0.014 a,B |
| DS8548-7 Li.u | 0.002 ± 0.002 a,A | 0.513 ± 0.022 a,A,B | 1.88 ± 0.09 a,A | 0.210 ± 0.016 a,B |
| DS8548-7 La.p | 0.004 ± 0.001 a,A | 0.534 ± 0.043 a,B | 1.82 ± 0.10 a,A | 0.214 ± 0.012 a,B |
| Purple Wheat | ||||
| DS8535-2 | <0.002 | 0.482 ± 0.041 a,A | 1.75 ± 0.16 a,A | 0.173 ± 0.016 a,A |
| DS8535-2 Pa | 0.006 ± 0.001 a,B | 0.458 ± 0.039 a,A | 1.86 ± 0.20 a,B | 0.162 ± 0.015 a,A |
| DS8535-2 Li.u | 0.006 ± 0.002 a,A,B | 0.495 ± 0.036 a,A | 1.77 ± 0.14 a,A | 0.163 ± 0.012 a,A |
| DS8535-2 La.p | 0.006 ± 0.001 a,A | 0.438 ± 0.034 a,A | 1.84 ± 0.11 a,A | 0.162 ± 0.013 a,A |
| Wholemeal Wheat Samples | Essential Microelements, Dry Matter (d.m.) | ||||
|---|---|---|---|---|---|
| Cr | Mn | Fe | Cu | Zn | |
| mg/100 g | mg/kg | ||||
| Traditional Wheat | |||||
| ‘Gaja’ | <0.010 | 12.8 ± 1.3 a,B | 24.8 ± 2.1 a,B | 2.19 ± 0.22 a,C | 6.56 ± 0.61 a,A |
| ‘Gaja’ Pa | <0.010 | 11.7 ± 1.2 a,B | 22.4 ± 2.0 a,C | 2.01 ± 0.18 a,C | 6.46 ± 0.59 a,A |
| ‘Gaja’ Li.u | <0.010 | 11.7 ± 1.2 a,B | 25.0 ± 2.3 a | 2.14 ± 0.16 a,C | 6.03 ± 0.42 a,A,B |
| ‘Gaja’ La.p | <0.010 | 11.5 ± 1.2 a,B | 24.8 ± 2.2 a,B | 2.23 ± 0.18 a,D | 6.21 ± 0.42 a,A |
| Waxy Wheat | |||||
| DS8888-3-6 | 0.151 ± 0.014 a | 7.54 ± 0.71 a,A | 15.6 ± 1.4 a,A | 1.14 ± 0.11 a,A,B | 5.51 ± 0.55 a,A |
| DS8888-3-6 Pa | 0.161 ± 0.007 a | 7.77 ± 0.80 a,A | 14.6 ± 1.3 a,A | 1.27 ± 0.13 a,B | 5.81 ± 0.57 a,A |
| DS8888-3-6 Li.u | 0.145 ± 0.025 a | 7.69 ± 0.92 a,A | 15.2 ± 1.7 a,A | 1.23 ± 0.12 a,A,B | 5.87 ± 0.42 a,A |
| DS8888-3-6 La.p | 0.158 ± 0.014 a | 7.71 ± 1.0 a,A | 15.9 ± 1.6 a,A | 1.22 ± 0.19 a,C | 5.74 ± 0.53 a,A |
| Blue Wheat | |||||
| DS8548-7 | <0.010 | 15.6 ± 1.3 a,C | 16.2 ± 1.6 a,A | 0.973 ± 0.097 a,A | 6.65 ± 0.63 a,A |
| DS8548-7 Pa | <0.010 | 15.4 ± 1.2 a,C | 15.0 ± 1.1 a,A | 0.926 ± 0.073 a,A,B | 6.73 ± 0.50 a,A |
| DS8548-7 Li.u | <0.010 | 14.1 ± 1.1 a,C | 16.7 ± 1.2 a,A | 0.938 ± 0.074 a,A | 6.38 ± 0.51 a,A,B |
| DS8548-7 La.p | <0.010 | 15.3 ± 1.0 a,C | 16.5 ± 1.2 a,A | 0.921 ± 0.071 a,B | 6.43 ± 0.58 a,A,B |
| Purple Wheat | |||||
| DS8535-2 | <0.010 | 17.8 ± 1.5 a,C | 16.6 ± 1.7 a,A | 0.805 ± 0.081 a,A | 8.31 ± 0.79 a,B |
| DS8535-2 Pa | <0.010 | 16.1 ± 1.3 a,C | 17.3 ± 1.8 a,A,B | 0.797 ± 0.080 a,A | 7.94 ± 0.75 a,A,B |
| DS8535-2 Li.u | <0.010 | 17.9 ± 1.2 a,D | 16.7 ± 1.5 a,A | 0.775 ± 0.078 a,A | 7.57 ± 0.72 a,B |
| DS8535-2 La.p | <0.010 | 17.4 ± 1.2 a,C | 17.0 ± 1.3 a,A | 0.764 ± 0.070 a,A | 7.54 ± 0.61 a,B |
| Wholemeal Wheat Samples | Non-Essential Microelements, mg/kg (d.m.) | |||||
|---|---|---|---|---|---|---|
| As | Rb | Sr | Cd | Ba | Pb | |
| Traditional Wheat | ||||||
| ‘Gaja’ | 0.007 ± 0.001 a | 1.44 ± 0.14 a | 0.963 ± 0.091 a,A | 0.054 ± 0.005 a,C | 2.35 ± 0.24 a,A | 0.012 ± 0.001 a,A |
| ‘Gaja’ Pa | 0.009 ± 0.002 a | 1.36 ± 0.12 a | 0.976 ± 0.083 a,A | 0.053 ± 0.006 a,D | 2.21 ± 0.19 a,A | 0.014 ± 0.002 a,A |
| ‘Gaja’ Li.u | 0.008 ± 0.001 a | 1.41 ± 0.10 a | 0.984 ± 0.074 a,A | 0.057 ± 0.004 a,D | 2.33 ± 0.22 a,A | 0.013 ± 0.001 a,A |
| ‘Gaja’ La.p | 0.008 ± 0.002 a | 1.43 ± 0.11 a | 0.962 ± 0.087 a,A | 0.058 ± 0.005 a,D | 2.36 ± 0.23 a,A | 0.012 ± 0.001 a,A |
| Waxy Wheat | ||||||
| DS8888-3-6 | <0.005 | <1 | 1.50 ± 0.13 a,B | 0.029 ± 0.003 a,B | 2.22 ± 0.21 a,A | <0.010 |
| DS8888-3-6 Pa | <0.005 | <1 | 1.46 ± 0.12 a,A | 0.033 ± 0.003 a,C | 2.45 ± 0.25 a,A,B | <0.010 |
| DS8888-3-6 Li.u | <0.005 | <1 | 1.42 ± 0.13 a,B | 0.035 ± 0.004 a,C | 2.23 ± 0.22 a,A | <0.010 |
| DS8888-3-6 La.p | <0.005 | <1 | 1.52 ± 0.14 a,B | 0.034 ± 0.004 a,C | 2.35 ± 0.26 a,A | <0.010 |
| Blue Wheat | ||||||
| DS8548-7 | <0.005 | <1 | 2.58 ± 0.26 D | 0.021 ± 0.002 a,A | 2.90 ± 0.29 a,B | <0.010 |
| DS8548-7 Pa | <0.005 | <1 | 2.59 ± 0.20 C | 0.020 ± 0.001 a,B | 2.78 ± 0.26 a,B | <0.010 |
| DS8548-7 Li.u | <0.005 | <1 | 2.66 ± 0.19 D | 0.022 ± 0.001 a,B | 2.83 ± 0.20 a,B | <0.010 |
| DS8548-7 La.p | <0.005 | <1 | 2.63 ± 0.21 D | 0.023 ± 0.002 a,B | 2.69 ± 0.25 a,A | <0.010 |
| Purple Wheat | ||||||
| DS8535-2 | <0.005 | <1 | 2.00 ± 0.20 a,C | 0.019 ± 0.003 a,A | <2 | 0.032 ± 0.003 a,B |
| DS8535-2 Pa | <0.005 | <1 | 1.97 ± 0.19 a,B | 0.016 ± 0.002 a,A | <2 | 0.029 ± 0.002 a,B |
| DS8535-2 Li.u | <0.005 | <1 | 1.90 ± 0.14 a,C | 0.017 ± 0.003 a,A | <2 | 0.031 ± 0.003 a,B |
| DS8535-2 La.p | <0.005 | <1 | 1.87 ± 0.17 a,C | 0.016 ± 0.002 a,A | <2 | 0.028 ± 0.003 a,B |
| Wholemeal Wheat Samples | Fatty Acid Composition, % from the Total Fatty Acid Content | |||||
|---|---|---|---|---|---|---|
| SFA | MUFA | PUFA | Omega-3 (ω-3) | Omega-6 (ω-6) | Omega-9 (ω-9) | |
| Traditional Wheat | ||||||
| ‘Gaja’ | 29.85 ± 0.09 c,A | 16.07 ± 0.08 a,B | 54.08 ± 0.25 c,D | 3.22 ± 0.06 a,A | 50.86 ± 0.32 c,C | 16.07 ± 0.11 a,A |
| ‘Gaja’ Pa | 26.38 ± 0.06 b,C | 17.51 ± 0.05 b,C | 56.11 ± 0.21 d,B | 4.48 ± 0.11 b,C | 51.63 ± 0.29 d,B | 17.51 ± 0.06 b,C |
| ‘Gaja’ Li.u | 26.31 ± 0.04 b,C | 21.34 ± 0.12 c,D | 52.35 ± 0.17 b,B | 11.85 ± 0.23 d,D | 40.50 ± 0.25 a,A | 21.34 ± 0.14 c,D |
| ‘Gaja’ La.p | 24.73 ± 0.11 a,A | 24.84 ± 0.14 d,D | 50.44 ± 0.22 a,C | 6.20 ± 0.10 c,D | 44.24 ± 0.23 b,B | 24.84 ± 0.13 d,D |
| Waxy Wheat | ||||||
| DS8888-3-6 | 34.54 ± 0.12 c,C | 16.54 ± 0.11 b,C | 48.92 ± 0.11 b,C | 3.39 ± 0.09 b,B | 45.53 ± 0.26 b,B | 16.54 ± 0.14 b,B |
| DS8888-3-6 Pa | 22.26 ± 0.08 b,A | 15.10 ± 0.10 a,A | 62.64 ± 0.23 c,D | 4.78 ± 0.11 c,D | 57.86 ± 0.19 d,D | 15.10 ± 0.13 a,A |
| DS8888-3-6 Li.u | 21.06 ± 0.14 a,A | 15.23 ± 0.12 a,A | 63.71 ± 0.35 d,D | 8.18 ± 0.14 d,C | 55.52 ± 0.22 c,C | 15.23 ± 0.10 a,A |
| DS8888-3-6 La.p | 37.73 ± 0.25 d,D | 23.91 ± 0.15 c,C | 38.37 ± 0.28 a,A | 2.17 ± 0.08 a,A | 36.20 ± 0.14 a,A | 23.91 ± 0.19 c,C |
| Blue Wheat | ||||||
| DS8548-7 | 38.14 ± 0.32 c,D | 16.81 ± 0.22 c,C | 45.04 ± 0.26 b,A | 3.46 ± 0.13 c,B | 41.58 ± 0.21 a,A | 16.81 ± 0.14 c,B |
| DS8548-7 Pa | 24.77 ± 0.18 a,B | 16.31 ± 0.12 b,B | 58.92 ± 0.33 d,C | 3.80 ± 0.09 d,B | 55.12 ± 0.19 c,C | 16.31 ± 0.12 b,B |
| DS8548-7 Li.u | 38.97 ± 0.15 d,D | 17.23 ± 0.09 d,B | 43.80 ± 0.19 a,A | 2.53 ± 0.07 a,A | 41.27 ± 0.22 a,B | 17.23 ± 0.15 d,B |
| DS8548-7 La.p | 33.87 ± 0.12 b,B | 15.12 ± 0.10 a,A | 51.02 ± 0.17 c,D | 2.90 ± 0.14 b,B | 48.11 ± 0.25 b,C | 15.12 ± 0.10 a,A |
| Purple Wheat | ||||||
| DS8535-2 | 32.01 ± 0.03 c,B | 21.33 ± 0.13 c,A | 46.66 ± 0.15 b,B | 4.71 ± 0.15 c,C | 41.95 ± 0.15 a,A | 21.33 ± 0.11 c,C |
| DS8535-2 Pa | 31.99 ± 0.14 b,D | 22.38 ± 0.09 d,D | 45.63 ± 0.21 a,A | 2.80 ± 0.10 a,A | 42.83 ± 0.22 b,A | 22.38 ± 0.13 d,D |
| DS8535-2 Li.u | 21.86 ± 0.12 a,B | 19.16 ± 0.15 b,C | 58.99 ± 0.16 d,C | 3.73 ± 0.09 b,B | 55.26 ± 0.31 d,C | 19.15 ± 0.18 b,C |
| DS8535-2 La.p | 34.30 ± 0.24 d,C | 17.98 ± 0.13 a,B | 47.72 ± 0.32 c,B | 3.66 ± 0.07 b,C | 44.06 ± 0.14 c,B | 17.98 ± 0.15 a,B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mockus, E.; Starkute, V.; Zokaityte, E.; Klupsaite, D.; Bartkevics, V.; Borisova, A.; Rocha, J.M.; Ruibys, R.; Liatukas, Z.; Ruzgas, V.; et al. The Potential of Traditional ‘Gaja’ and New Breed Lines of Waxy, Blue and Purple Wheat in Wholemeal Flour Fermentation. Fermentation 2022, 8, 563. https://doi.org/10.3390/fermentation8100563
Mockus E, Starkute V, Zokaityte E, Klupsaite D, Bartkevics V, Borisova A, Rocha JM, Ruibys R, Liatukas Z, Ruzgas V, et al. The Potential of Traditional ‘Gaja’ and New Breed Lines of Waxy, Blue and Purple Wheat in Wholemeal Flour Fermentation. Fermentation. 2022; 8(10):563. https://doi.org/10.3390/fermentation8100563
Chicago/Turabian StyleMockus, Ernestas, Vytaute Starkute, Egle Zokaityte, Dovile Klupsaite, Vadims Bartkevics, Anastasija Borisova, João Miguel Rocha, Romas Ruibys, Zilvinas Liatukas, Vytautas Ruzgas, and et al. 2022. "The Potential of Traditional ‘Gaja’ and New Breed Lines of Waxy, Blue and Purple Wheat in Wholemeal Flour Fermentation" Fermentation 8, no. 10: 563. https://doi.org/10.3390/fermentation8100563
APA StyleMockus, E., Starkute, V., Zokaityte, E., Klupsaite, D., Bartkevics, V., Borisova, A., Rocha, J. M., Ruibys, R., Liatukas, Z., Ruzgas, V., & Bartkiene, E. (2022). The Potential of Traditional ‘Gaja’ and New Breed Lines of Waxy, Blue and Purple Wheat in Wholemeal Flour Fermentation. Fermentation, 8(10), 563. https://doi.org/10.3390/fermentation8100563