Abstract
Postbiotics are functional bioactive substances manufactured during fermentation in a food matrix, which can be used to improve human health, but their influence on the adhesion potential and physicochemical cell surface of probiotics is still unclear. We examined the postbiotic influence produced by Escherichia coli Nissle 1917 in functional labneh on cell surface properties (auto-aggregation, hydrophobicity, and co-aggregation) and the adhesion capacities of three probiotic strains. The most commonly detected effects of probiotics, particularly Lsyn−7, were an increase in auto-aggregation, hydrophobicity, co-aggregation, and adhesion ability of the tested strains. Lactobacillus rhamnosus with Lsyn−7 (59%) presented the highest hydrophobicity, whereas the least adhesion to xylene was detected in L. rhamnosus with LHM. Lactobacillus casei with Lsyn−7 showed the highest auto-aggregation after 24 h (60.55%). Moreover, it also has a strong adhesion to Caco-2 cells and effectively prevents the binding of Salmonella Typhimurium to Caco-2 cells. Lactobacillus plantarum with Lsyn−7 presented the strongest co-aggregation with Staphylococcus aureus (85.1%), S. typhimurium (85. 02%) and Listeria monocytogenes (77.4%). The adherence potential of tested probiotic strains was highly correlated with auto-aggregation, hydrophobicity, co-aggregation, and competitive inhibition of L. monocytogenes and S. typhimurium. The findings suggest that Lsyn−7 can be a candidate to promote the adhesion potential of selected probiotic strains. For the reason that the application of probiotic strains has been more interested in their positive influences in the gastrointestinal tract, it is essential to use some functional compounds, such as postbiotics, to improve adhesion abilities and cell surface properties in terms of bacterial binding.
1. Introduction
The word “-biotics” indicates strategies that fit with proper nutrition that can be used to modulate the colonic microflora towards healthy benefit on the host. The word ‘biotic’ is formed from the Greek term “biotikós”, which means “relating to life”. It can be used to describe the biological ecosystem consisting of organism communities joined with their natural environment []. Probiotic strains modify the intestinal microbiota and were particularly used to improve the health of humans and animals []. Presently, many well-characterized strains of bifidobacterial and lactobacilli are available for human use for the benefit of the host []. Besides lactic acid bacteria, Escherichia coli Nissle 1917 (EcN) have also been used as a probiotic and therapeutic agent for over a century []. It was isolated by Alfred Nissle in 1917 from a German soldier who remained healthy, whereas his companions were subjected to infections caused by Shigella spp. []. Nissle is the unique probiotic active ingredient in Mutaflor, a licensed pharmaceutical broadly available in Australia, Europe and Canada [].
EcN has immunomodulatory influences, including upregulation of beneficial responses, as well as suppression of immune-mediated damage, and it can enhance the function of the intestinal barrier []. Moreover, it could produce some novel type of colicin that decrease the growth of pathogens []. The efficacy and safety of EcN have been widely studied. Several previous studies have presented that EcN is efficient for the prevention and treatment of gastrointestinal disorders, including chronic constipation, ulcerative colitis, irritable bowel syndrome, and Crohn’s disease. It has also been exhibited to be safe when administered to infants []. Although EcN is able to utilize lactose [], there was no evidence in the literature that reported the possibility using of EcN as starter culture for the production of fermented milk.
Prebiotics refers to compounds that are particularly used by probiotics to confer health benefits []. Prebiotics can modulate the colonic microflora composition by activating the growth of the probiotic, thereby promoting health benefits in the host []. These compounds included phytochemicals, phenolics, polyunsaturated fatty acids, conjugated linoleic acids, human milk oligosaccharides, many oligosaccharides, and several dietary fiber types []. Galactooligosaccharides (GOS) are lactose-derived ingredients that are synthesized during hydrolysis by the enzyme β-galactosidase in a side reaction called transglycosylation []. GOS are particularly used in human nutrition as they are compositionally identical to a complicated variety of structures in breast milk []. GOS are extensively utilized in infant formulas because bovine milk has small concentration of oligosaccharides (0.005%) []. However, human colostrum includes about 2.0–2.5% g/L, which decreases to 0.5 to 2.0% in mature breast milk []. Lactose is used as a precursor for GOS by β-galactosidases produced by fungi, bacteria, or yeast []. When probiotics and prebiotics are integrated, their influences are considered in the form of synergism, and they are denominated synbiotics [].
The newest member of the biotics family, postbiotics, refers to bioactive ingredients produced by probiotic strains during the process of fermentation. Postbiotics contain metabolites and cell components [].
Historically, the knowledge and discovery of beneficial microbes or probiotics are associated with commonly consumed fermented foods, and it was approved that probiotic strains have been transferred to us since the first food raw materials were subjected to fermentation process []. Cultured dairy products were probably the first functional food supplemented with probiotics. They are the best carrier of probiotic strains in the production of dairy products. Several studies presented the idea that probiotic cultures have been effectively used in milk fermentation and where they induced preferable sensory and textural properties, apart from beneficials effects on human health []. Labneh is an artisanal culture milk product. It is a popular dairy product in different parts of the world, particularly in the regions of the Middle East, in which it plays an essential role in the family diet [].
There were many studies dealing with the effect of prebiotics on probiotic strains. However, so far, only a few studies have highlighted the correlation between the adherence of probiotics and prebiotics addition [,].
Prebiotics could enhance the adherence of probiotics, proposing that the production of novel synbiotic dairy products might be a potential agent to increase probiotic colonization in the human gut []. Moreover, prebiotics could also present as decoy receptors, preventing the attachment of the pathogen to the intestinal mucosa, as reported by Hickey []. To our knowledge, no prior findings have investigated the influence of postbiotics on the adhesion of probiotics.
This study aimed to investigate the effect of postbiotics on the adhesion abilities of many probiotics, thus increasing our knowledge about their interactions. This was conducted by examining the influence of postbiotics manufactured by E. coli Nissle 1917 in functional labneh enriched with GOS, the adhesion potential and physicochemical cell surface features of three probiotic strains.
2. Materials and Methods
2.1. Production of Hydrolyzed Lactose Cow’s Milk
Hydrolyzed lactose milk was used in this study as a prebiotic, which was produced by treating with 0.1% (w/w) β-galactosidase (3.2.1.23; Sigma-Aldrich, St. Louis, MO, USA) at 37 °C for 60 min. The resultant milk contained galactooligosaccharides (GOS) that were synthesized during the reaction with β-galactosidase []. The lyophilization of the hydrolyzed milk (LHM) was considered a control.
2.2. Production of Labneh
Labneh was manufactured as previously described with slight modification []. Briefly, hydrolyzed lactose cow’s milk was heated for 15 min at 94 °C and then inoculated with 2% of E. coli Nissle 1917 at 45 °C, then the mixture was incubated at 37 °C for complete coagulation. The resultant curd was then slowly stirred and put into cloth bags stacked on top of each other and left for 24 h at 5 °C. Labneh was obtained from cloth bags, mixed with 2% NaCl, put into plastic containers, and stored at 5 °C for two weeks. Samples for the preparation of lyophilized supernatant were taken at zero-time (Lsyn−0), 7 (Lsyn−7), and 15 (Lsyn−15) days during refrigerated storage. Subsequently, sterile distilled water (2.5 mL) was added to the 10 g labneh sample, then adjusted to pH 4 with 0.1-NHCl, followed by ten minutes of incubation at 45 °C. The crude extract (CE) was centrifuged (4000× g, 15 min, 5 °C). NaOH (0.1 N) was used to adjust the pH of supernatants to 7.0, followed by re-centrifugation (4000× g, 15 min, 5 °C) for further salts and protein precipitation. The supernatants were obtained and filtered with sterilized 0.22 µm Millipore filters. The final supernatants were lyophilized and used within ten hours of preparation [].
2.3. Culture Condition of Selected Probiotic Strains
All used microbial strains were obtained from the food microbiology laboratory at the dairy department, Faculty of Agriculture, Mansoura University, Mansoura, Egypt (Escherichia coli Nissle 1917, Lactobacillus rhamnosus DML182, Lactobacillus casei DML191, Lactobacillus plantarum DML23, Listeria monocytogenes DML34, Salmonella Typhimurium DML113, and Staphylococcus aureus ATCC 25923).
The selected strains were grown overnight in Lactobacillus MRS broth o/w dextrose (dextrose is not involved in this culture media). 2-gL−1 postbiotics (Lsyn−0, Lsyn−7, and Lsyn−15) or LHM (control) were added to culture media as a carbon source followed by incubation at 37 °C. However, E. coli Nissle 1917 and pathogenic strains of S. aureus, L. monocytogenes and S. typhimurium were cultivated in tryptone soy broth supplemented with 0.1% yeast extract for overnight at 37 °C.
The pellets were obtained by centrifugation for ten minutes at 5000× g at laboratory temperature. The phosphate buffer (pH 6.6) was used to wash the pellets thrice and it was also used to adjust the optical density 0.60–0.65 at 600 nm (A0, H0).
2.4. Bacterial Adhesion to Solvents (BATS)
The adhesion of tested bacterial strains to solvent assay was conducted to investigate the features of physicochemical cell surface []. Briefly, adherence to a polar solvent (xylene) describes bacterial cell surface hydrophobicity. The affinities to basic solvent (ethyl acetate) and acidic solvent (chloroform) show the electron acceptor features and electron donor of the cell surface of bacteria, respectively. Cell pellets were washed thrice and resuspended in PBS (phosphate-buffered saline; 7.2) to obtain suspensions of 108 CFU mL−1. The 1.5 mL bacterial cell suspension was mixed with postbiotic solution (1.5 mL; 5% (wt/vol)), followed by addition into a solvent (1 mL); ethyl acetate as an electron acceptor, chloroform as an electron donor and xylene as an apolar solvent. The blend was mixed by vortexing for two minutes and incubated at room temperature (20 °C) without agitation for 30 min to split into two layers. The absorbance of the collected aqueous layer was measured at 600 nm. The affinities to various solvents were expressed as adhesion percentages based on the subsequent formula
where A and A0 are the absorbances after and before extraction using different solvents, respectively.
BATS (%) = (1 − A/A0 min) × 100,
2.5. Assessment of Auto-Aggregation of Selected Probiotics
The determination of the auto-aggregation was performed following the Zuo et al. [] with minor modifications. Concisely, for the assay of the auto-aggregation, each bacterial suspension (2 mL) was mixed using postbiotic solution (2 mL; 5% (wt/vol)) at laboratory temperature for 20 s, followed by incubation at 37 °C. The absorbance of mixtures was measured at 600 nm after 3, 6, and 24 h (At). The values of auto-aggregation were calculated depending on the subsequent equation:
Auto-aggregation (%) = (1 − At/A0) × 100
2.6. Co-Aggregation Determination of Pathogens with Lactobacilli Strains
The co-aggregation ability of tested strains was determined according to the method previously described [] with slight modification. Briefly, suspensions of tested strains were prepared as reported for auto-regulation analysis. Equal cell volumes (1.5 mL) of the different pathogens and probiotics were mixed with postbiotic solution (3 mL; 5% (wt/vol)) and incubated for three hours at 37 °C without agitation. The mixture’s absorbance was measured after three hours at 600-nm. The co-aggregation (%) was calculated by subsequent equation:
Co-aggregation (%) = [1 − Amix/(Aprobiotic + Apathogen)/2] × 100
2.7. Adhesion of Probiotic or Pathogenic Strains to Caco-2 Cells
The influence of postbiotics on the adhesive activity of foodborne pathogens or probiotic strains were determined using an intestinal epithelial cell model (Caco-2 cells) as an intestinal epithelial cell model based on the previous study reported by Nueno-Palop and Narbad [] with minor modification. Briefly, Dulbecco’s modified Eagle’s minimal essential medium was used as culture media for the Caco-2 cell line (ATCC HTB-39). This medium fortified with L-glutamine (2 mmol L−1), 10% fetal bovine serum (Sigma-Aldrich), penicillin (100-U ml−1) and streptomycin (100-µg mL−1) and then incubated (37 °C, 5% CO2 (v/v) and 95% (v/v) humidified air) until the formation of confluent monolayers.
The Caco-2 cell monolayers on the 24-well plates were washed with phosphate-buffered saline (PBS) at pH 7.2 before adhesion assay of selected strains. Bacterial pellets were obtained using centrifugation at 7500× g for 20 min at 7 °C, followed by washing thrice with PBS (pH 7.2). The bacterial pellets were resuspended in Dulbecco’s modified Eagle’s minimal essential medium (fetal-bovine-serum-free and antibiotic-free). In total, 500 mL of bacterial suspension were blended with an identical volume solution of postbiotics (5%). The mixture was poured into the 24-well plates, followed by incubation for one hour at 37 °C in 5% CO2. The free or nonattached bacterial cells were eliminated from Caco-2 cells by washing each plate’s well with PBS (pH 7.2). Finally, 1 mL Triton X-100 (1% (v/v) was put to each well. Then, the mixture was mixed to separate the attached microbial cells from monolayers of Caco-2 cells. A series of sequential dilutions of S. typhimurium, S. aureus, L. monocytogenes, and lactobacilli strains were plated on xylose lysine deoxycholate agar, Baird-parker agar, modified oxford agar, and MRS, respectively, followed by incubation at 37 °C for 48 h. The adhesion percentage was calculated using the adhesion potential of pathogens or probiotics numbers to Caco-2 cells when matched with the inoculum level.
2.8. Determination of Competitive Inhibition of Pathogens Binding
To evaluate the ability of competitive inhibition of pathogen binding, 250 µL pathogenic bacterial suspension was mixed with lactobacilli strains at a ratio of 1:1. Five hundred microliters of postbiotics solution (5%) was added. The combination of lactobacilli-pathogens with/without postbiotics was added to 24-well plates, including monolayers of Caco-2 cells. The plates were incubated at 37 °C for one hour. These steps are the same steps that are used for the adherence potential of probiotics or pathogenic strains to Caco-2 cells. The inhibition of pathogenic strains adhesion was determined by the number of pathogens attached to Caco-2 cells compared with the number of pathogens attached to Caco-2 without lactobacilli strains [].
2.9. Statistical Analysis
All experiments were conducted in triplicate. ANOVA test with a significance level of p < 0.05 was used to assess the parameters of physicochemical cell surface and adhesion ability of tested strain. The results are indicated as average ± standard deviation. Significant divergences among mean values were evaluated using Duncan’s multiple range tests. SPSS Statistics software version 16 (SPSS Inc., Chicago, IL, USA) was applied to assess all statistical tests in this study. Principal component analysis (PCA) was conducted to present the relationship between selected lactobacilli strains’ cell surface properties and their adhesion to Caco-2 cells.
3. Results
3.1. The Effect of Postbiotics on Cell Surface Properties of Probiotic Strains
Microbial adhesion to hydrocarbon (MATH) methods are the main prevalently used protocol for determining the cell surface hydrophobicity of probiotics []. This protocol appreciates hydrophobicity as the adhesion of selected probiotic strains to apolar solvent (e.g., xylene, toluene, and hexane). In this study, the affinity of tested strain to xylene was performed to determine bacterial hydrophobicity. The adhesive features of tested strains associated with postbiotics (Lsyn−0, Lsyn−7, and Lsyn−15) to aromatic hydrocarbon (xylene), halogenated aliphatic hydrocarbon (chloroform), and aliphatic ester (ethyl acetate) are indicated in Figure 1. All tested probiotics grown in MRS broth enriched with Lsyn−7 showed significantly (p ˂ 0.05) higher hydrophobicity than other treatments (Figure 1). The hydrophobicity of all probiotic strains grown with postbiotics was in the order Lsyn−7 > Lsyn−15 > Lsyn−0 > control (Figure 1). The high hydrophobicity (˃50%) was detected for the L. rhamnosus with Lsyn−7 (59.53%), L.plantarum with Lsyn−7 (52.91%), and L.casei with Lsyn−7 (52.19%) (Figure 1). For the test strains with different postbiotics, non-significant (p ˂ 0.05) differences in affinities with chloroform and ethyl acetate, except for L. rhamnosus associated with Lsyn−7, where its affinity with chloroform was shown to be relatively higher than control (Figure 1). The adhesion rate of tested strains to an aliphatic ester (ethyl acetate) was lower than xylene or chloroform, within a range between 6.2% and 8.9%.
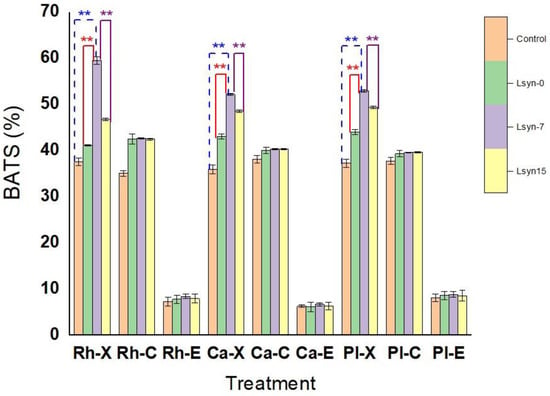
Figure 1.
The influence of postbiotics (Lsyn−0, Lsyn−7 and Lsyn−15) on affinities of selected probiotics (L. rhamnosus (Rh), L. casei (Ca) and L. plantarum (Pl) to xylene (X), chloroform (C), and ethyl acetate (E). ** p ˂ 0.01.
3.2. The Influence of Postbiotics on Auto-Aggregation Activity of Probiotic Strains
The auto-aggregation features of selected probiotics are shown in Figure 2. In the recent work, after incubation for 24 h, all selected probiotic strains showed a high ability of auto-aggregation compared with their ability incubated at three or six hours (Figure 2). A time-dependent rise in this auto-aggregation capacity was detected (Figure 2). All strains associated with Lsyn−7 presented significantly (p ˂ 0.05) higher values of auto-aggregation, within a range from 56.1% to 60.5%, than other treatments (Figure 2). L. casei exhibited the greatest auto-aggregative ability at the end of incubation (24 h), whereas L. rhamnosus showed the lowest ability of auto-aggregative (56.1%) (Figure 2).
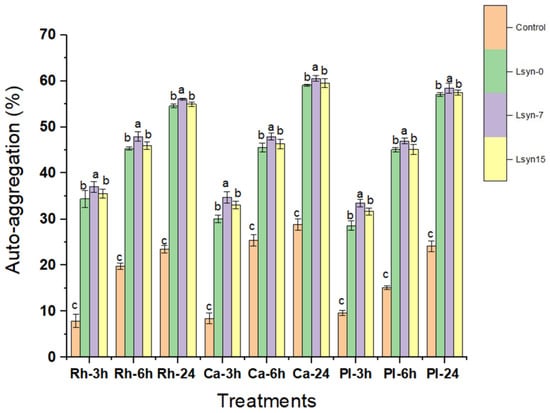
Figure 2.
The effect of postbiotics (Lsyn−0, Lsyn−7 and Lsyn−15) on auto-aggregative properties of L. rhamnosus (Rh), L. casei (Ca) and L. plantarum (Pl) after incubated at 37 °C for 3 h, 6 h, and 24 h. Different letters show significant differences at p < 0.05. Error bars indicate the standard division of the sample’s mean.
3.3. The Effect of Postbiotics on Co-Aggregation Activity of Pathogens with Probiotic Strains
The co-aggregation between foodborne pathogens and probiotics is indicated in Figure 3. The high co-aggregation (˃70%) was observed for almost all probiotics associated with Lsyn−7 and Lsyn−15 against foodborne pathogens (Figure 3). Among the tested strains, L. plantarum with Lsyn−7 showed significantly (p ˂ 0.05) highest abilities of co-aggregation with L. monocytogenes (85.02%) and S. aureus (85.1%), but no significant difference between the effects of L. plantarum and L. rhamnosus are associated with Lsyn−7 against S. Typhimurim (Figure 3). The probiotic strains associated with LHM demonstrated the least co-aggregation capacity against all foodborne pathogens. Predominantly, the co-aggregation was related to auto-aggregation and hydrophobicity.
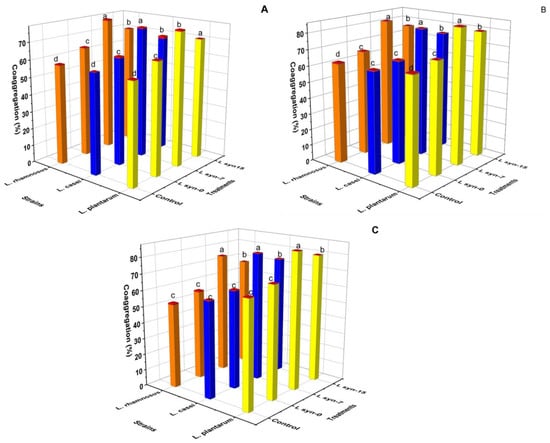
Figure 3.
The effect of postbiotics (Lsyn−0, Lsyn−7 and Lsyn−15) on the ability of selected probiotic strains to co-aggregate with foodborne pathogenic bacteria, L. monocytogenes (A), S. typhimurium (B), S. aureus (C) after three hours incubation at 37 °C. Different letters indicate significant differences at p < 0.05. Redcaps of the columns show the standard division of the sample’s mean.
3.4. In Vitro Assessment of Postbiotics on Adhesion of Probiotic or Pathogenic Strains
The effect of postbiotics on adhesion capacities of selected strains of foodborne pathogens and probiotics in Caco-2 cells is indicated in Figure 4 and Table S1. The adherence ability highly varied between the tested strains. Generally, all selected strains can bind to Caco-2 cells under in vitro conditions; their ability of adherence ranged between 5.76% and 26.82%, with L. casei, L. plantarum, and L. rhamnosus associated with Lsyn−7 adhering most effectively at 26.82%, 26.69%, and 23.66%, respectively (Figure 4; Table S1). In contrast S. Typhimurim, S. aureus and L. monocytogenes associated with Lsyn−7 were least efficient, with adhesion ability of 5.7%, 6.01%, and 6.3%, respectively (Figure 4). In our research, Lsyn−7 induced a two-way adjusting influence on the adherence to Caco-2 cells, where Lsyn−7 significantly increased the adhesion potential of lactobacilli strains to Caco-2 cells compared with other treatments. Still, the same treatment significantly reduced the adhesion potential of pathogens to Caco-2 compared with other treatments (Figure 4).
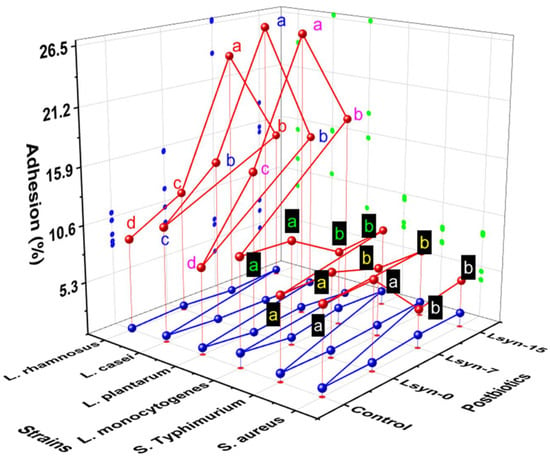
Figure 4.
The influence of postbiotics (Lsyn−0, Lsyn−7 and Lsyn−15) on adherence ability (red sphere) of strains of foodborne pathogenic bacteria and probiotics to Caco-2 cells. Different letters indicate significant differences at p < 0.05. The blue sphere shows the standard division of the sample’s mean.
3.5. In Vitro Evaluation of Postbiotics on Competitive Inhibition of Pathogens Adhesion to Caco-2 Cells by Probiotic Strains
The competitive inhibition of adherence of S. aureus, S. typhimurium, L. monocytogenes to Caco-2 cells by selected lactobacilli strains is indicated in Figure 5. The selected lactobacilli strain enhanced inhibition of the adhesion abilities of S. aureus (1.3%–4.8%), S. Typhimurim (5.3%–27.55%) and L. monocytogenes (1.8%–31.18%) to Caco-2 cells. Lsyn−7 significantly increases the rate of competitive inhibition of the adhesion of L. monocytogenes and S. typhimurium compared with other treatments, regardless of the lactobacilli strain, was used in this test. However, the competitive inhibition of S aureus was not significantly observed in the presence of selected lactobacilli strains associated with different postbiotics (Figure 5).
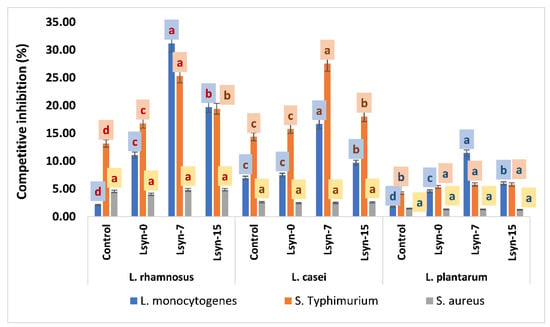
Figure 5.
The influence of postbiotics (Lsyn−0, Lsyn−7 and Lsyn−15) on competitive inhibition to adhere S. aureus, S. typhimurium and L. monocytogenes to Caco-2 cells by lactobacilli strains. Different letters indicate significant differences at p < 0.05. The Error bars show the standard division of the sample’s mean.
3.6. Multivariate Analysis of Adhesion Potential and Cell Surface Characteristics of Selected Lactobacilli Strains
PCA of the influence of postbiotics on cell surface properties (bacterial adhesion to solvent, auto-aggregation, and co-aggregation) and adhesion potential of selected probiotic strains explained 78.06% of the variability on two PC (Figure 6A). PC1 (56.31%) comprised bacterial adhesion to xylene (%BATS-x), bacterial adhesion to chloroform (%BATS-C), auto-aggregation (Agg) after 3, 6, and 24 h, co-aggregation, (Coagg), adhesion to Caco-2 cells (Adhesion) and competitive inhibition of L. monocytogenes (L. inhibition). However, the second dimension (21.75%) was primarily associated with competitive inhibition of S. typhimurium and S. aureus (Figure 6A). Four groups, groups (2 and 3) were positioned at the left side of PC1. However, group one and four were put on the positive value side of PC1. Group one presented high values of cell surface properties and adhesion potential of selected strain. In contrast, group four is characterized by high values of all parameters except the competitive inhibition of S. typhimurium and S. aureus (Figure 6A). All values of tested parameters are decreased in groups two and three compared with other groups, but the rate of competitive inhibition of S. typhimurium and S. aureus in group three is higher than group four (Figure 6A). Figure 6B is used to define the relationship between cell surface properties of selected strains and their adhesion to Caco-2 cells. The gastrointestinal tract adhesion (Caco-2 cells) is strongly correlated with hydrophobicity and co-aggregation. The auto-aggregation (3, 6, and 24 h) and competitive inhibition of L. monocytogenes is strongly correlated with the adhesion rate, whereas the rate of adhesion is moderately correlated with bacterial adhesion to chloroform. Finally, the degree of correlation between adhesion of tested strains and bacterial adhesion to ethyl acetate and competitive inhibition of S. typhimurium is weak. The competitive inhibition of S. aureus is negatively correlated with the rate of adhesion to Caco-2 cells (Figure 6B).
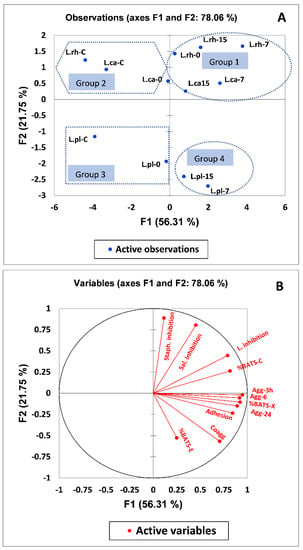
Figure 6.
(A) Principal component (PC) analysis biplot of physicochemical cell surface properties and adhesion potential of L. rhamnosus (Lrh), L. casei (Lca) and L. plantarum (Lpl) tested strain. (B) The correlation of principal component analysis variable. The following abbreviation C, 0, 7, 15, Agg and Coagg refer to control, zero-time, 7 and 15 days of storage periods, auto-aggregation, and co-aggregation respectively.
4. Discussion
An optimization of the enzymatic synthesis was performed to result the maximum contents of GOS, the enzymatic treatment was performed with ß-galactosidase concentration of 0.1% (w/w) and incubated at 37 °C for 60 min. generally, such as in previous studies, similar enzymatic synthesis process for GOS productions was performed [,]. Oh, Kim, Oh and Kim [] reported also that catalyzed reaction of transgalactosylation resulted GOS with different linkages such as ß1-6, ß1-4 and ß1-3. Moreover, the concentration of residual lactose after 60 min of enzymatic reaction had reduced by 98.16% and the total content of GOS was 0.71 g/100 g. GOS mainly comprised allolactose, 6-galactobiose and 6-galactosyllactose.
To the best of the authors’ knowledge, no study has been conducted on determining the postbiotics effect on the cell surface properties (auto-aggregation, hydrophobicity, and co-aggregation) and the adhesion potential of selected probiotic strains.
This study determines the effect of postbiotics on adhesion abilities, and cell surface characteristics, of selected probiotics. Previous studies have confirmed that higher adhesion to Caco-2 cells because of higher hydrophobicity of the cell surface and vice versa [].
The basic solvent (ethyl acetate), non-polar (Xylene) and acidic solvent (chloroform) were used to determine the electron acceptor, hydrophobic/hydrophilic, and electron donor properties of bacterial cell surface, respectively (Figure 1), which resulted from Lewis’s acid-base and carboxylic groups interactions [].
The hydrophilic and hydrophobic features are attributed to polysaccharides and proteins on the bacterial surface []. The selected strains exhibited weak adhesion to ethyl acetate, which demonstrated the particularity of selected strains having the nonacidic and poor electron acceptor property [], compared with their affinities to an acidic solvent (chloroform), which showed the specialty of selected strains to have an acidic character and essential electron donor.
The saccharide concentration in the culture medium plays a critical role in measuring the hydrophobicity of cell surface. When the presence of lactose as a sole carbon source in culture media positively influences hydrophobicity of cell surface []. The culture media enriched with prebiotic levan also enhances the hydrophobicity of the probiotic strain L. acidophilus La5 []. In contrast contrary, the presence of hydrochloric acid and bile salts is inversely proportional to the hydrophobicity of bacterial cell surface []. Substantially, the MATH method does not determine the cell surface’s hydrophobicity; instead, it reflects van der Waals and electrostatic forces as substantial factors interacting with overall adhesion capacity []. The relationship between hydrophobicity and adhesion has been detected in many studies with conflicting results because cell adhesion features are affected by different factors such as pH, temperature, and culture media composition. [,,].
According to Falah et al. [], investigation of hydrophobicity may be considered a preliminary test of the ability of probiotics to bind to epithelial cells. They also consider hydrophobicity as one of the significant features enhancing the first contact between host cells and probiotic strains. Some authors also consider that probiotics with high hydrophobicity have better potency to bind to the human intestinal [].
Bacterial auto-aggregation is an approach by which bacterial strains physically react with each other and precipitate from static liquid suspension []. Bacterial ability to form cellular aggregates through the bacterial aggregation of the same strain (auto-aggregation) can also participate in colonization in the intestine. Furthermore, aggregation may act antagonistically against potentially pathogenic microorganisms [,]. The auto-aggregation ability of probiotics is directly proportional to their adhesion to intestinal mucosa []. To succeed in the required advantage of probiotic bacteria, they need to create an adequately large biomass through aggregation []. Some studies have found a relationship between adhesion and aggregation []. However, other studies did not find such a link and correlation between these two characteristics []. Depending on these results, probiotic strains had perfect auto-aggregation characteristics, usually having good adhesion abilities to cell culture, usually had good auto-aggregation properties. This correlation could have been seen in the case of tested strains, particularly when they are associated with Lsyn−7. A strong correlation was found between hydrophobicity and auto-aggregation of selected strains in this study (Figure 6B). This finding corresponds to the study by Tuo et al. [], who reported that auto-aggregation levels of the 15 L. plantarum strains strongly correlated with hydrophobicity. However, our findings do not correspond to the study by Vlková et al. [], who described that no significant correlations existed between hydrophobicity and auto-aggregation ability of bifidobacteria. The correlation between hydrophobicity and auto-aggregation may be species-specific.
Co-aggregation of probiotics plays a critical role in preventing pathogenic strains from surface colonization []. It was recently reported that probiotics aggregation efficiently produced various antimicrobial compounds [], suggesting that co-aggregation and auto-aggregation are closely associated with the antagonistic influence of probiotic strains.
Co-aggregation of probiotics with a potential pathogen allows them to produce antimicrobial compounds in very close proximity to pathogens, which may prevent the growth of the pathogenic strain in the gastrointestinal tract []. It was reported that the ability of co-aggregation with potential pathogens and auto-aggregation properties can be used for preliminary selection of probiotic bacteria [,]. Co-aggregation and auto-aggregation are significant in biofilm formation to protect the host from colonization by potential pathogenic strains []. The co-aggregation of the selected strains was highly correlated with auto-aggregation and hydrophobicity. These results are inconsistent with those by Collado et al. [], who reported that auto-aggregation properties are related to co-aggregation capacities. Xu, Jeong, Lee and Ahn [] also showed that the co-aggregative was strongly correlated with hydrophobicity.
Adherence to epithelial cells and following colonization of the gastrointestinal tract is a characteristic that gives a significant advantage to probiotic strains and supports them to proliferate and compete in the gut effectively. The enterocyte-like Caco-2 cell model is particularly used to determine the competitive inhibition, displacement, adhesion rate because the ability of adhesion to epithelial cells is significantly considered an important standard to select while isolating novel probiotic bacteria [,]. Kadlec and Jakubec [] reported that Enterococcus durans, Bifidobacterium animalis ssp. lactis and L. rhamnosus exhibited increases in adhesion abilities after prebiotic (vivinal or P95) addition. Different results from multiple combinations of prebiotics and probiotic strains suggest that different adhesion mechanisms in the probiotic strains play a critical role. These modes of action are reported only in general terms, and so far, no evidence occurs as to why two kinds of prebiotics can have a different influence on adhesion properties of the same strain.
Many previous studies have investigated probiotic strains for their adhesion abilities; however, their studies are hardly comparable because of differences in tissue models []. Previous studies have shown a correlation between hydrophobicity and adhesion ability in some lactobacilli strains []. In our study, the adhesion of the selected strains was highly correlated with co-aggregation, auto-aggregation, and hydrophobicity, as indicated in Figure 6B. These results are not in accordance with the results of other studies [,].
The influence of these three postbiotics (Lsyn−0, Lsyn−7, and Lsyn−15) on the adhesion ability of selected probiotic strains were generally positive and, as mentioned above, particularly the addition of Lsyn−7 to tested strains. It is, thence, potential that hydrolyzed GOS, particularly Lsyn−7, is better at enhancing adhesion ability because raw, prebiotic GOS in this study had the lowest effect on cell surface features and adhesion potential of all monitored probiotics compared with other treatments. The highest impact of postbiotic on cell surface features and adhesion abilities of selected strains up to day seven (Lsyn−7) and then a reduction after day 15 (Lsyn−15) but L syn−15 is still higher than Lsyn−0 or control. This finding is consistent with previous reports []. The positive effect of Lsyn−7 on adhesion abilities of selected probiotic strains might be attributed to high concentration of calcium and other divalent cations []. The concentration of diffusible calcium in fermented milk increased along with progress of cold storage period compared with casein-bound calcium []. Additionally, a previous study reported that prebiotics was added to probiotic strain (L. rhamnosus), the ζ-potential was increased by about 10 mV. Modification in negative charge intensity is probably one of the essential factors affecting adhesion ability [].
All selected lactobacilli strains associated with Lsyn−7 significantly (p ˂ 0.05) inhibited the adhesion of L. monocytogenes and S. typhimurium (Figure 6). This finding suggests a correlation between adhesion of probiotic strains to Caco-2 cells and competitive inhibition, which is competitively preventing pathogenic bacteria. The mechanism of bacterial adhesion to the gastrointestinal tract is more complicated, including extracellular and cell surface receptors []. The adhesion of foodborne pathogens may also be affected by the direct effect of postbiotics against invading pathogens or indirect effect by stimulating probiotic metabolites, such as bacteriocins, polysaccharides, peptides, hydrogen peroxide, and organic acids, where many studies have reported the effect of probiotic metabolites on the adhesion of pathogens [,]. The decline in the adhesion of L. monocytogenes and S. typhimurium by selected probiotic strains with or without postbiotics may be due to steric hindrance that selected lactobacilli strains to compete with foodborne pathogens for adhesion sites (adherence-receptor interactions) []. The general correlation between in vivo colonization and in vitro adhesion ability has been proposed [].
Therefore, the most important finding in the current study was that postbiotic (Lsyn−7) can promote GI colonization by probiotic strains. Once a stable and balanced gut flora is attained, the microflora and their metabolites may contribute to the colonization and homeostasis with competitive inhibition of pathogens in the gastrointestinal tract. This study would provide significant information for the effect of postbiotic on the adhesion of probiotic strains, according to the correlation between cell surface properties and adhesion to Caco-2 cells. Therefore, further studies are required to understand the use of postbiotic as an improvement tool for probiotic colonization and how to select a suitable postbiotic for variations in strains, species, and genera of probiotics in terms of adhesion, cell surface properties, and competitive inhibition of pathogens in GIT.
5. Conclusions
Because the usage of probiotic bacteria has been more interested with their beneficial influences in the gastrointestinal tract, it is important to investigate essential factors that effect on physicochemical cell surface and adhesive features of selected probiotic strains.
We found, under the present study conditions, that the influences of postbiotics produced by EcN, particularly Lsyn−7 on cell surface and adhesive properties of selected probiotic strains are positive and that they could improve on the competitive inhibition of foodborne pathogen adhesion to Caco-2 cells by probiotics. The effect of postbiotic on the adhesion of probiotics is strongly strain-specific and each incorporation should be investigated before application. However, the mechanisms of the effects of postbiotics on auto-aggregation, adhesion, co-aggregation, cell surface characteristics of probiotic strains should be further studied.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation8030122/s1, Table S1: Competitive inhibition of adherence of food borne pathogens to the Caco-2 cells by selected probiotic strains.
Author Contributions
Conceptualization M.S.D. and E.K.; methodology E.K., M.S.D., M.S.G., E.G.E. and A.E.; software E.K., M.S.D., M.S.G., E.G.E. and A.E. validation, E.K., M.S.D., M.S.G., E.G.E. and A.E.; formal analysis, E.K., M.S.D., M.S.G., E.G.E. and A.E.; investigation, E.K., M.S.D., M.S.G., E.G.E. and A.E.; resources, E.K., M.S.D., M.S.G., E.G.E. and A.E.; data curation E.K., M.S.D., M.S.G., E.G.E. and A.E.; writing—original draft preparation, M.S.D. and E.K. review and editing, E.K., M.S.D., M.S.G., E.G.E. and A.E.; visualization, E.K., M.S.D., M.S.G., E.G.E. and A.E.; supervision, M.S.D., M.S.G. and E.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Taif University Researchers supporting project number (TURSP-2020/307), Taif University, Taif, Saudi Arabia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are available from the corresponding author on reasonable request.
Acknowledgments
Taif University Researchers supporting project number (TURSP-2020/307), Taif University, Taif, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wegh, C.A.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and their potential applications in early life nutrition and beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, J.H.; Choi, S.H.; Park, S.W.; Choi, N.-J.; Kim, Y.; Kim, S.H. Synbiotic impact of tagatose on viability of Lactobacillus rhamnosus strain GG mediated by the phosphotransferase system (PTS). Food Microbiol. 2013, 36, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Weiss Alison, A.; Barbour Alan, G. Probiotic Properties of Escherichia coli Nissle in Human Intestinal Organoids. mBio 2020, 11, e01470-20. [Google Scholar] [CrossRef] [PubMed]
- Henker, J.; Laass, M.W.; Blokhin, B.M.; Maydannik, V.G.; Bolbot, Y.K.; Elze, M.; Wolff, C.; Schreiner, A.; Schulze, J. Probiotic Escherichia coli Nissle 1917 Versus Placebo for Treating Diarrhea of Greater Than 4 Days Duration in Infants and Toddlers. Pediatr. Infect. Dis. J. 2008, 27, 494–499. [Google Scholar] [CrossRef]
- Ramachandran, C.; Rani, R.S.; Usha, A. Evaluation of safety, antimicrobial activity and probiotic properties of Escherichia coli Nissle 1917 isolated from Idli batter. Res. J. Biotechnol. Vol. 2016, 11, 7. [Google Scholar]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Vyas, U.; Ranganathan, N. Probiotics, prebiotics, and synbiotics: Gut and beyond. Gastroenterol. Res. Pract. 2012, 2012, 872716. [Google Scholar] [CrossRef] [Green Version]
- Torres, D.P.M.; Gonçalves, M.d.P.F.; Teixeira, J.A.; Rodrigues, L.R. Galacto-oligosaccharides: Production, properties, applications, and significance as prebiotics. Compr. Rev. Food Sci. Food Saf. 2010, 9, 438–454. [Google Scholar] [CrossRef] [Green Version]
- Oh, N.S.; Kim, K.; Oh, S.; Kim, Y. Enhanced production of galactooligosaccharides enriched skim milk and applied to potentially synbiotic fermented milk with Lactobacillus rhamnosus 4B15. Food Sci. Anim. Resour. 2019, 39, 725. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Ferez, A.; Guadix, A.; Guadix, E.M. Recovery of caprine milk oligosaccharides with ceramic membranes. J. Membr. Sci. 2006, 276, 23–30. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Zhang, G.; Sadiq, F.A.; Simal-Gandara, J.; Xiao, J.; Sang, Y. Probiotics in the dairy industry—Advances and opportunities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3937–3982. [Google Scholar] [CrossRef] [PubMed]
- Saad, S.; Salama, H.; El-Sayed, H. Manufacture of functional Labneh using Uf-retentate and artichoke puree. Int. J. Dairy Sci. 2015, 10, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Kavanaugh, D.W.; O’Callaghan, J.; Butto, L.F.; Slattery, H.; Lane, J.; Clyne, M.; Kane, M.; Joshi, L.; Hickey, R.M. Exposure of Bifidobacterium longum subsp. infantis to milk oligosaccharides increases adhesion to epithelial cells and induces a substantial transcriptional response. PLoS ONE 2013, 8, e67224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celebioglu, H.U.; Olesen, S.V.; Prehn, K.; Lahtinen, S.J.; Brix, S.; Abou Hachem, M.; Svensson, B. Mucin-and carbohydrate-stimulated adhesion and subproteome changes of the probiotic bacterium Lactobacillus acidophilus NCFM. J. Proteom. 2017, 163, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Hickey, R.M. The role of oligosaccharides from human milk and other sources in prevention of pathogen adhesion. Int. Dairy J. 2012, 22, 141–146. [Google Scholar] [CrossRef]
- Aly, A.A.; Refaey, M.M.; Hameed, A.M.; Sayqal, A.; Abdella, S.A.; Mohamed, A.S.; Hassan, M.; Ismail, H.A. Effect of addition sesame seeds powder with different ratio on microstructural and some properties of low fat Labneh. Arab. J. Chem. 2020, 13, 7572–7582. [Google Scholar] [CrossRef]
- Shori, A.; Baba, A. Antioxidant activity and inhibition of key enzymes linked to type-2 diabetes and hypertension by Azadirachta indica-yogurt. J. Saudi Chem. Soc. 2013, 17, 295–301. [Google Scholar] [CrossRef]
- Xu, H.; Jeong, H.; Lee, H.; Ahn, J. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Lett. Appl. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef]
- Zuo, F.; Yu, R.; Feng, X.; Chen, L.; Zeng, Z.; Khaskheli, G.B.; Ma, H.; Chen, S. Characterization and in vitro properties of potential probiotic Bifidobacterium strains isolated from breast-fed infant feces. Ann. Microbiol. 2016, 66, 1027–1037. [Google Scholar] [CrossRef]
- Nueno-Palop, C.; Narbad, A. Probiotic assessment of Enterococcus faecalis CP58 isolated from human gut. Int. J. Food Microbiol. 2011, 145, 390–394. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, O.; Muthaiyan, A.; Moreno, F.J.; Montilla, A.; Sanz, M.L.; Ricke, S.C. Effect of prebiotic carbohydrates on the growth and tolerance of Lactobacillus. Food Microbiol. 2012, 30, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; O’Sullivan, D.J. Production of galactooligosaccharides using a hyperthermophilic β-galactosidase in permeabilized whole cells of Lactococcus lactis. J. Dairy Sci. 2014, 97, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauvière, G.; Coconnier, M.-H.; Kerneis, S.; Darfeuille-Michaud, A.; Joly, B.; Servin, A.L. Competitive exclusion of diarrheagenic Escherichia coli (ETEC) from human enterocyte-like Caco-2 cells by heat-killed Lactobacillus. FEMS Microbiol. Lett. 1992, 91, 213–217. [Google Scholar] [CrossRef]
- Pelletier, C.; Bouley, C.; Cayuela, C.; Bouttier, S.; Bourlioux, P.; Bellon-Fontaine, M.N. Cell surface characteristics of Lactobacillus casei subsp. casei, Lactobacillus paracasei subsp. paracasei, and Lactobacillus rhamnosus strains. Appl. Environ. Microbiol. 1997, 63, 1725–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakirova, L.; Grube, M.; Gavare, M.; Auzina, L.; Zikmanis, P. Lactobacillus acidophilus La5 and Bifidobacterium lactis Bb12 cell surface hydrophobicity and survival of the cells under adverse environmental conditions. J. Ind. Microbiol. Biotechnol. 2013, 40, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Martienssen, M.; Reichel, O.; Kohlweyer, U. Surface Properties of Bacteria from Different Wastewater Treatment Plants. Acta Biotechnol. 2001, 21, 207–225. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; de Cadiñanos, L.P.G.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Falah, F.; Vasiee, A.; Behbahani, B.A.; Yazdi, F.T.; Moradi, S.; Mortazavi, S.A.; Roshanak, S. Evaluation of adherence and anti-infective properties of probiotic Lactobacillus fermentum strain 4–17 against Escherichia coli causing urinary tract infection in humans. Microb. Pathog. 2019, 131, 246–253. [Google Scholar] [CrossRef]
- de Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum Strains Isolated from Mozzarella Cheese: Probiotic Potential, Safety, Acidifying Kinetic Parameters and Viability under Gastrointestinal Tract Conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Sorroche, F.G.; Spesia, M.B.; Zorreguieta, Á.; Giordano, W. A positive correlation between bacterial autoaggregation and biofilm formation in native Sinorhizobium meliloti isolates from Argentina. Appl. Environ. Microbiol. 2012, 78, 4092–4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 2008, 226, 1065–1073. [Google Scholar] [CrossRef]
- Krausova, G.; Hyrslova, I.; Hynstova, I. In Vitro Evaluation of Adhesion Capacity, Hydrophobicity, and Auto-Aggregation of Newly Isolated Potential Probiotic Strains. Fermentation 2019, 5, 100. [Google Scholar] [CrossRef] [Green Version]
- Tuo, Y.; Yu, H.; Ai, L.; Wu, Z.; Guo, B.; Chen, W. Aggregation and adhesion properties of 22 Lactobacillus strains. J. Dairy Sci. 2013, 96, 4252–4257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlková, E.; Rada, V.; Šmehilová, M.; Killer, J. Auto-aggregation and co-aggregation ability in bifidobacteria and clostridia. Folia Microbiol. 2008, 53, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.L.; Grześkowiak, Ł.; Collado, M.C.; Salminen, S. In Vitro Evaluation of Lactobacillus gasseri Strains of Infant Origin on Adhesion and Aggregation of Specific Pathogens. J. Food Prot. 2011, 74, 1482–1487. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.-R.; Patra, J.K.; Kim, W.J.; Kang, S.-S. Antagonistic Activities and Probiotic Potential of Lactic Acid Bacteria Derived From a Plant-Based Fermented Food. Front. Microbiol. 2018, 9, 1963. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Surono, I.; Meriluoto, J.; Salminen, S. Indigenous dadih lactic acid bacteria: Cell-surface properties and interactions with pathogens. J. Food Sci. 2007, 72, M89–M93. [Google Scholar] [CrossRef]
- Kadlec, R.; Jakubec, M. The effect of prebiotics on adherence of probiotics. J. Dairy Sci. 2014, 97, 1983–1990. [Google Scholar] [CrossRef]
- Ehrmann, M.A.; Kurzak, P.; Bauer, J.; Vogel, R.F. Characterization of lactobacilli towards their use as probiotic adjuncts in poultry. J. Appl. Microbiol. 2002, 92, 966–975. [Google Scholar] [CrossRef]
- Mathara, J.M.; Schillinger, U.; Guigas, C.; Franz, C.; Kutima, P.M.; Mbugua, S.K.; Shin, H.K.; Holzapfel, W.H. Functional characteristics of Lactobacillus spp. from traditional Maasai fermented milk products in Kenya. Int. J. Food Microbiol. 2008, 126, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zárate, G.; De Ambrosini, V.M.; Chaia, A.P.; González, S. Some factors affecting the adherence of probiotic Propionibacterium acidipropionici CRL 1198 to intestinal epithelial cells. Can. J. Microbiol. 2002, 48, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.O.; Mahgoub, S.A.; Sitohy, M.Z. Hindering milk quality storage deterioration by mild thermization combined with methylated chickpea protein. Int. Food Res. J. 2014, 21, 693–701. [Google Scholar]
- Lee, Y.K.; Lim, C.Y.; Teng, W.L.; Ouwehand, A.C.; Tuomola, E.M.; Salminen, S. Quantitative approach in the study of adhesion of lactic acid bacteria to intestinal cells and their competition with enterobacteria. Appl. Environ. Microbiol. 2000, 66, 3692–3697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.K.; Puong, K.Y. Competition for adhesion between probiotics and human gastrointestinal pathogens in the presence of carbohydrate. Br. J. Nutr. 2002, 88, S101–S108. [Google Scholar] [CrossRef] [Green Version]
- Guglielmetti, S.; Tamagnini, I.; Mora, D.; Minuzzo, M.; Scarafoni, A.; Arioli, S.; Hellman, J.; Karp, M.; Parini, C. Implication of an Outer Surface Lipoprotein in Adhesion of Bifidobacterium bifidum to Caco-2 Cells. Appl. Environ. Microbiol. 2008, 74, 4695–4702. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).