Abstract
Taiwan djulis (Chenopodium formosanum Koidz.) is a plant native to Taiwan and is a grain rich in nutrients, vitamins, and minerals with antioxidant properties. This paper aimed to use appropriate processing technology and incorporate probiotics, thus combining Taiwan’s high-quality milk sources to develop Taiwan djulis fermented dairy products. Later, FL83B cells have used to evaluate the glucose utilization ability after the administration of djulis. We first screened Lactiplantibacillus plantarum and combined it with the traditional yogurt strains Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus for cultivation. Further, the fermentation process was optimized where 7.5% djulis and an inoculum of 107 colony forming unit/mL were fermented at 40 °C for 18 h. Compared to fermented milk without djulis, the analysis of various nutrients and active ingredients showed that free radical scavenging abilities of DPPH and ABTS reached 2.3 and 2.0 times (752.35 ± 29.29 µg and 771.52 ± 3.79 µg TE/g, respectively). The free phenol content increased 2.5 times (169.90 ± 14.59 mg gallic acid/g); the total flavonoid content enhanced 4.8 times (3.05 ± 0.03 mg quercetin/g), and the gamma-aminobutyric acid content was 3.07 ± 0.94 mg/g. In a co-culture of mouse liver cells with fermented products, 100 ppm ethanol extract of fermented products effectively improved glucose utilization with increased glucose transporter expression. This functional fermented dairy product can be developed into the high value added local agricultural products and enhance multiple applications including medical and therapeutic fields.
1. Introduction
Chenopodium formosanum, also known as Taiwan djulis, is a plant endemic to Taiwan. Djulis has become a high-market-value food source gaining international attention because it is gluten-free, has a low glycemic index, and is rich in essential amino acids, fiber, lipids, carbohydrates, vitamins, and minerals. In addition to nutritional properties djulis has higher betalains and total polyphenol compounds. It is an important traditional crop and is one of the ingredients of the local wine brewed by aboriginal people in Taiwan [1,2]. With the migration of aboriginal people or changes in economic patterns, Taiwan’s djulis production is reduced with time. However, in recent years, the trend of healthy eating has become essential along with its influence in the international community, djulis has quickly become an important ingredient in food. The United Nations (UN) declared 2013 as the International Year of djulis (IYQ). The existence of this “miracle” plant was a well-kept secret from most of the world [3]. However, with its promotion, Taiwanese djulis has gradually attracted the attention of Chinese farmers. In 2015, the planting area was about 40 hectares, which expanded massively to 200 hectares in 2018. However, excessive plantation led to an imbalance between the supply and demand, which made Taiwan djulis unsaleable that year and its purchase price plummeted. Due to the impact of the COVID-19 epidemic, the Taiwan djulis market dropped by >50%. Therefore, devising new applications or products through processing methods can help to increase export value of Taiwan djulis.
Chenopodium formosanum, known as djulis, is a native plant in Taiwan [4]. Taiwanese aborigines have harvested djulis for hundreds of years. Recently, the biological effects of djulis have been investigated with properties in reducing cholesterol levels [5], preventing colon cancer [6], inhibiting hyperglycemia, hyperlipidemia [7], and adipogenecity [8]. It also possesses anti-oxidant [9], anti-inflammatory [10], anti-allergic [11], and hepatic protective properties [12]. The main functional components and dietary fiber of Taiwan djulis come from its shell. However, due to the strong and bitter taste of the shell, it is not accepted easily by the consumers. Currently, the commercially available Taiwan djulis is mostly in its deshelled form and consumed in combination with staple foods such as rice or brown rice. The main aim of this study was to use deshelled djulis to make fermented beverages with functional ingredients.
Recently, consumers have specialized demands for probiotic-related products, and this project aimed to develop health care products by targeting the physiological needs of specific consumer groups. Gamma-aminobutyric acid (GABA) is a non-protein amino acid with a four-carbon structure. It is an inhibitory neurotransmitter in the sympathetic nervous system with anti-diabetic and anti-hypertensive effects on humans. Additionally, it inhibits the proliferation of cancer cells, controls the lipid quality in serum, and reduces pain and anxiety. Past studies have found that lactic acid bacteria can be used to ferment plant milk to produce GABA [13,14,15]. Therefore, in this project, it is aimed that specific lactic acid bacteria strains can be identified through screening to degrade the protein-rich djulis, thereby increasing its GABA content to contribute to health benefits. In addition, Taiwan djulis contains active ingredients such as polyphenols or flavonoids. If lactic acid bacteria can ferment it, its rich protein can be degraded into functional peptides, and flavonoid biotransformation may synergize with other active ingredients to regulate blood sugar levels. In this regard, functional evaluations were conducted to assess the ability to reduce blood sugar through a cell culture system. The efficacy of blood sugar regulation or reducing the risk of type 2 diabetes is a pressing priority in developing health/therapeutic products. Thus, the study will benefit from a perspective of healthy fermented product development but also from increasing the value of local agricultural products.
2. Materials and Methods
2.1. Materials
Mouse glucose transporter 2 immunoassay kit was purchased from Jingmei (Jiangsu, China). Taiwan djulis planted in Pingtung County, Taiwan, were purchased from Lujinwang Biotechnology Co., Ltd. (Pingtung County, Taiwan). Ruisui Whole Cream Fresh Milk was purchased from Uni-President (Taipei, Taiwan). 2,2′-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS), Gallic acid, Folin–Ciocalteu phenol reagent, α,α-diphenyl-β-picrylhydrazyl (DPPH), Sodium hydroxide, Hydrogen chloride, Gallic acid, Sodium carbonate, Sodium nitrite, Aluminum chloride, Daidzin, Daidzein, Genistin, Genistein, Sodium tetraborate, Sodium dodecyl sulfate, 2-mercaptopropinoic acid, Sodium acetate, Petroleum ether, Sulfosalicylic acid, Iron (III) chloride hexahydrate, Sodium phytate, Oleanolic acid, Dimethyl sulfoxide, Glucose, Insulin, o-toludine, Trolox, Ammonium acetate, 2-Hydroxynaphthaldehyde, γ-Aminobutyric acid, 9-Fluorenylmethyloxycarbonyl chloride, o-phtalaldehyde, sodium phytate, Petroleum ether, Vanillin, and soy saponin were purchased from Sigma-Aldrich Co. (Merck, Darmstadt, Germany). The 95% Ethanol, Methanol, Acetone, and Sulfuric acid were purchased from Jingming Chemical Co., Ltd. (Taipei, Taiwan). Agar and Peptone were purchased from BioShop (BioShop Canda Inc., Ontario, QC, Canada). Cell counting kit-8 was purchased from Danjindo (Danjindo, Kumamoto, Japan). Fetal bovine serum, Ham’s F-12K (Kaighn’s) Medium, Trypsin, and KRB Buffer were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Glucose assay kit was purchased from Abbkine (Hardy Diagnostics, Santa Maria CA, USA). Lactobacilli MRS Broth was purchased from Hardy diagnostics (Santa Maria, CA, USA).
2.2. Microorganisms and Medium
Ten strains were selected to test the best combination of strains to ferment the dairy products. Bifidobacterium infantis BCRC 14602, Bifidobacterium adolescentis BCRC 14606, Bifidobacterium bifidum BCRC 14615, Bifidobacterium longum BCRC 14634, Bifidobacterium breve BCRC 11846, Lacticaseibacillus rhamnosus BCRC 16000, Lactobacillus delbrueckii subsp. bulgaricus BCRC 10696, Lactiplantibacillus plantarum BCRC 11697, Lactobacillus acidophilus BCRC 14079, Streptococcus salivarius subsp. thermophiles BCRC 14085 and FL83B hepatocyte cell line were purchased from Bioresource Collection and Research Center (BCRC, Hsinchu city, Taiwan) and cultured in MRS medium for routine use (Sigma-Aldrich, St. Louis, MO, USA). Bacteria were stored in MRS broth with 20% glycerol, at −80 °C, for long-term storage. For the activation of bacteria, all strains were thawed at room temperature, and 1% of bacteria (v/v) was inoculated into the MRS broth and cultured at a constant temperature of 37 °C, for 48 h, and sub-cultured twice a week.
2.3. Plate Counting
The samples were serially diluted ten-fold with 0.1% Peptone water to appropriate multiples, and 100 µL was applied to Lactobacilli MRS agar and incubated at 37 °C. The agar plates were incubated for 24 h, the number of bacteria was counted, and the logarithm was taken.
2.4. Fermentation and Fermentation Broth Freeze Drying
The fresh milk and different concentration of djulis were distributed evenly in the sterilized conical flask. The lactic acid bacteria were activated for 24 h, and 1% (v/v) (7 log colony forming unit (CFU)/mL) inoculum was added into the Taiwan djulis liquid. The Erlenmeyer flask was placed in a 37 °C incubator for 18 h. Of the fermented products, 1 mL was collected in a sterilized 1.5 mL microfuge tube (for the number of bacteria), 3 mL into a 10 mL centrifugation tube (for pH value), and 1 mL into a sterilized 1.5 mL microfuge tube (for DPPH, ABTS). Samples were stored at −20 °C until assessment.
The fermented products were placed in a freezer overnight for cooling, then transferred to a freezer vacuum dryer, cooled to −50 °C, and freeze-dried for 48 h. The fermented products dry powder weighed and stored at −20 °C. Subsequent experiments were carried out using the lyophilized samples.
2.5. DPPH Scavenging Activity
DPPH method was effective in describing the antioxidant ability of sample. The method for determination of the DPPH scavenging activity reported by Hu et al. [16] and Wang et al. [17] was modified. Fermented samples were freeze-dried and re-dissolved to 5 mg/mL in deionized water. A freshly prepared solution of DPPH in ethanol (100 µM) was mixed 1:1 with the sample, then stored in the darkroom for 30 min. The absorbance was measured at 517 nm, calculated to Trolox equivalent antioxidant capacity (TEAC) and presented as Trolox equivalent (µg/g).
2.6. ABTS Activity
ABTS method were reliable in determining the antioxidant ability of the sample. Freeze-dried fermentation samples were prepared at 5 mg/mL (in 80% MeOH); 7 mM ABTS and 2.45 mM potassium persulfate (K2S2O8) were prepared in deionized water, respectively. ABTS and potassium persulfate were mixed 1:1 in a dark environment, oxidized for 12 h (marked as ABTS stock solution), and diluted to 0.7 ± 0.02 at OD734 before use. In a 96-well plate, 20 µL of sample and 180 µL of ABTS solution were mixed, incubated for 6 min in a dark room, and the absorbance was measured at 734 nm, and calculated to Trolox equivalent antioxidant capacity (TEAC) and present as Trolox equivalent (µg/g) [18].
2.7. Preparation of Fermented Taiwan Djulis Functional Dairy Products
An appropriate amount of Taiwan djulis weighed, washed with water until no foaming, and 0.25 mL of water added per gram of Taiwan djulis, and filter sterilized. Fresh milk was added into sterilized serum bottles, heated at 65 °C for 20 min, sterilized, and stored at 4 °C for later use. In 100 mL of fresh milk, 7.5 g of sterilized Taiwan djulis was added, with 107 CFU/mL of Lactiplantibacillus plantarum BCRC 11697, Lactobacillus delbrueckii subsp. bulgaricus BCRC 10696 and Streptococcus salivarius subsp. thermophilus BCRC 14085, and cultured at 40 °C for 18 h. The samples were further freeze-dried to remove moisture and were stored at 4 °C.
2.7.1. pH and Titratable Acidity Determination
To 1 g of sample, 10 mL double distilled water was added, and its pH value was measured. Further, 10 g of sample was dissolved in 10 mL of ddH2O, and titrated to pH 8.1 with 0.1 N NaOH. The titratable acidity can be obtained according to the titratable acidity formula [19]. The formula is as follows:
- V: Titration of NaOH solution (mL);
- N: Normality of NaOH solution (N);
- F: potency of NaOH solution;
- B: Gram equivalents of organic acids, lactic acid is 90;
- W: sample weight (g).
2.7.2. Extraction and Determination of Total Phenols and Total Flavonoids
Phenols and flavonoids compound has various bioactive properties. Therefore, it is important to determine those compounds in the dairy products.
2.7.3. Extraction and Analysis of Free Phenol, Bound Phenol, and Total Phenol
Lyophilized sample powder (0.1 g) was weighed into 2 mL microfuge tube, to which 1 mL of acidic methanol (HCl)/water solution (50:50 v/v, pH 2) was added, shaken at room temperature for 1 h for extraction, followed by centrifugation at 2100× g at 4 °C for 10 min and filtration using a 0.22 mm filter to obtain the supernatant, which was stored at 4 °C in the dark. Further, 1 mL of acetone/water solution (70 mL) was added to the centrifuged pellet (30 v/v). The supernatant was obtained by extraction, centrifugation, and filtration under the same conditions and stored at 4 °C in the dark. Methanol/H2SO4 90:10 (v/v) was added (1 mL) to the centrifugation pellet, set to 85 °C, with oil bath extraction for 2 h, and centrifugation and filtration to obtain the supernatant, which was stored at 4 °C in the dark.
Gallic acid (1 mM) was prepared and diluted to 50, 125, 250, 500, 750, 1000 µM as a standard solution. The gallic acid standard solution (100 µL) of the above three extracts and blank group (water) were taken into 1.5 mL tube, and 100 µL of 50% Folin−Ciocalteu phenol reagent aqueous solution was added to each group. Sodium carbonate aqueous solution (5% w/v), incubated at 40 °C for 20 min, after cooling to room temperature, was taken (150 µL) into a 96-well plate, and the spectrophotometer was used to read the absorbance at 740 nm. The result is as follows: Gallic acid is expressed in equivalents (mg GAE g−1).
2.7.4. Extraction and Determination of Total Flavonoids
The lyophilized sample (0.5 g) was weighed, 5 mL of ethanol was added and incubated at room temperature for 2 h. Using 200 µL of sample and 30 µL of 5% sodium nitrite, the reaction was carried out at room temperature for 6 min in the dark, and 60 µL of 10% aluminum chloride was allowed to react in the dark for 5 min, at room temperature. Further, 200 µL of 1 N sodium hydroxide solution was added, centrifuged at 8000× g for 30 s, and the absorbance of the supernatant was measured at 520 nm with a spectrophotometer and quercetin was used as a standard to make a standard curve. The results are expressed as quercetin equivalents (mg QE g−1).
2.8. Sensory Evaluation
The evaluation team consisted of 52 people (untrained, 33 women and 19 men, aged between 15 and 35). Overall, 54% (28/55) of the reviewers were general fermented milk consumers who purchased fermented milk more than once a week. The evaluation samples were commercial original yogurt, commercial cereal yogurt, homemade fermented dairy products, and Taiwan quinoa fermented dairy products. The samples were all packed in plastic cups and numbered with random numbers. The sensory evaluation form consists of two parts. The first part is the consumer acceptance test. The samples’ appearance, aroma, taste, flavor, aftertaste, and integrity were scored with a 9-point product scoring method, from 1 to 9, respectively: “I do not like it very much”, “I do not like it very much”, “I do not like it a little”, “I do not like it a little”, “I do not like it at all”, “I like it a little”, “I like it a little”, “I like it a lot”, and “Like”. The second part is the consumer preference test, which was scored according to the degree of preference.
2.9. GABA (γ-Aminobutyric Acid) Content
In this stage, 0.1 g lyophilized samples were extracted with 70% ethanol three times. The solvent extract was concentrated and resolved in 3 mL 0.1 M ammonium acetate and then used for derivatization by 2-HN (2-hydroxynaphthaldehyde) and HPLC analysis [19].
2.10. Analysis of Amino Acid Composition
This method was used to analyze amino acid composition to understand product nutritional value. We took 1 g of lyophilized samples and added 9 mL of ddwater for extraction for further analysis. The amino acids were derivatized by 9-fluorenylmethyloxycarbonyl chloride and o-phtalaldehyde. Additionally, they were then separated by Inertsil ODS-3 column (5 µM, 250 × 4.6 mm) and HPLC system (Hitachi, Tokyo, Japan) with a fluorescence detector (L-2480) and diode array detector (L-2455) [20,21].
2.11. Analysis of Phytic Acid
Anti-nutritional factors impeded nutrient absorption; therefore, this method was used to measure them. We took 1 g of lyophilized samples and added 9 mL of ddwater for extraction for further analysis. Then, 600 µL of sodium phytate or sample extraction was added with 200 µL of Wade reagent. After mixing, the absorbance of the solution was read at 500 nm. The peptide content was calculated on the standard curve constructed using sodium phytate standard [22].
2.12. Analysis of Saponin
Anti-nutritional factors impeded nutrient absorption. Therefore, this method was used to measure saponin content. We mixed 0.5 g of the lyophilized sample of the freeze-dried powder with 10 mL of petroleum ether. This was shaken at 100 rpm to extract the oil for 4 h, then a 90 mm filter paper was used to remove the supernatant and volatilize the petroleum ether, and 40 mg of the degreasing solid was taken with 5 mL 80% methanol and 100 rpm in the dark for 4 h. The supernatant was filtered through a 0.8 µm filter to obtain the saponin extract. We took 100 µL of saponin extract, 400 µL of 80% methanol, and 500 µL of 8% vanillin solution (dissolved in 99% ethanol) in an ice bath, mixed them, added 4 mL of 77% sulfuric acid, and placed the mixture in a water bath at 60 °C for 15 min. Then, we measure its absorbance at 550 nm, and used soybean saponin to dissolve in 80% methanol to make a standard curve with concentrations of 10, 5, 2.5, 1.25, 0.625, and 0 mg/mL. Results are expressed in mg soy saponin/g extract [23].
2.13. Cell Experiment
FL83B hepatocyte cell culture medium contained 90% F12K culture medium and 10% Fetal bovine serum (FBS) (v/v). The culture was in a 37 °C, 5% CO2 incubator for about 3–4 days. After removing the medium, 1 mL of trypsin was added to detach cells in the incubator for 10 min, and 0.5 mL of FBS was added to stop the reaction. Dilute the cell fluid to an appropriate multiple, and follow the protocol of the CCK-8 kit and Mouse glucose transporter two immunoassay kit for follow-up experiments.
2.14. Glucose Utilization Test
Determination of glucose utilization was a simple method to assess whether a sample had the potential to be used as a future dietary supplement for the regulation of blood glucose in the clinic. The method was modified by Liu et al. [24]. Briefly, after removing the supernatant and washing with PBS, F12K medium was added and incubated for 24 h in a 37 °C, 5% CO2 incubator. After removing the supernatant and washing with PBS, KRB buffer containing 67 mM glucose was added and incubated for 24 h in an incubator at 37 °C, with 5% CO2. The supernatant was removed, and the residual was washed with PBS, then 500 µL of each concentration of samples diluted with a culture medium were added. After incubating for 30 min in a 37 °C, 5% CO2 incubator, the supernatant was removed and washed with PBS, then insulin (5 µg/mL) was added after incubating for 30 min in a 37 °C, 5% CO2 incubator, and the culture medium was collected for analysis. Then, 10 µL of culture solution was taken and reacted with 200 µL o-toluidine reagent at 100 °C for 8 min. After cooling in a water bath for 4 min, the absorbance was measured at a wavelength of 450 nm.
2.15. Statistical Analysis
All analytical experiments were performed at least thrice, and three samples were analyzed for each test. Data were collected and analyzed using a one-way analysis of variance and Duncan’s test. Differences were considered significant at p < 0.05. All statistical analyses were performed using SPSS Statistics software (version 12.0, St. Armonk, NY, USA).
3. Results and Discussion
3.1. Optimal Lactobacillus Screening of Fermented Taiwan Djulis
According to our previous research, the Taiwan djulis substrate was maintained at 30 °C, inoculated with ten different strains of lactic acid bacteria, and fermented at 37 °C for 24 h. The optimal Lactobacillus screening of fermented Taiwan djulis Lactiplantibacillus plantarum BCRC 11697 displayed the best antioxidant capacity [25] and total phenolic content post fermentation. In addition, total phenolic content before fermentation was 22 mg of gallic acid/g, while after fermentation it was 28 mg of gallic acid/g (Figure S1), which was significantly higher than those of other lactic acid bacteria. Earlier studies used mold and quinoa for solid-state fermentation to increase the total phenolic content by 2.46 mg/g [26], where the total phenolic content increased by 6 mg/g after fermentation in this experiment.
Studies have demonstrated a significant correlation between antioxidant capacity (ABTS) and total phenolic content, R = 0.966, while the correlation between DPPH and total phenolics is R = 0.939 [27], thus suggesting that Lactiplantibacillus plantarum BCRC 11697 is effective for the fermentation process. Lactiplantibacillus plantarum is a lactic acid bacteria commonly found in plant-derived fermented foods and is often used as a bacterial model for the study of phenolic compound metabolism [28]; it is capable of degrading phenolic compounds in food, resulting in aroma compounds and compounds with increased antioxidant activity [29]. Moreover, TanBLp (also known as tannases) is present in Lactiplantibacillus plantarum, which is capable of hydrolyzing gallic acids and protocatechuic acid ester bonds [28], and feruloyl esterases are involved in enzymes that release phenolic compounds from plant cell walls and increase antioxidant activity [30].
Different lactic acid bacteria have different antioxidant capacities, which may affect their ability to utilize Taiwan djulis as the substrate. Lactiplantibacillus plantarum grows in plants, fruits, and vegetables and can also be found in fermented dairy products and pickled vegetables. A previous study found that fermentation of Lactiplantibacillus plantarum with grape pomace produced the best antioxidant capacity, similar to the results of this experiment [31].
3.2. Optimization of Conditions for Fermenting Taiwan Djulis
When making fermented djulis dairy products, the aforementioned probiotic Lactiplantibacillus plantarum BCRC 11697, with the best antioxidant activity, was used in combination with the traditional yogurt fermentation strains Lactobacillus delbrueckii subsp. bulgaricus BCRC 10696 and Streptococcus salivarius subsp. thermophilus BCRC 14085, and the fermentation temperature and strain inoculum were determined. Three fermentation conditions, including the addition of Taiwan djulis, were optimized with the fermentation time of 18 h, and the most appropriate fermentation conditions were selected based on the scavenging ability of DPPH free radicals. DPPH is one of the few stable and commercially available organic nitrogen free radicals whose scavenging ability is also one of the most widely used methods for determining antioxidant activity. Its drug preparation and analysis operations are relatively simple, and the experimental results are highly reproducible. Hence, the free radical scavenging ability of DPPH was used as an indicator in this study [32].
The first optimized conditions were the fermentation temperature at 37, 40, and 45 °C, respectively. The DPPH free radical scavenging abilities were 634.83 ± 20.44, 674.56 ± 18.35, and 673.54 ± 32.34 µg/g, respectively, and there was no significant difference among the three (Figure S2a). Therefore, the fermentation temperature of 40 °C with relatively high DPPH free radical scavenging ability was used. The biochemical reactions required in the biological growth process are all affected by temperature. The suitable growth temperature ranges of Lactiplantibacillus plantarum, Lactobacillus delbrueckii subsp. bulgaricus, and Streptococcus salivarius subsp. thermophilus have been reported to be 30–40, 43–46, and 35–42 °C, respectively [33,34], which are similar to the optimum fermentation temperature of this experiment.
Further, the inoculum amounts were optimized to be 106, 107, and 108 CFU/mL, with their DPPH free radical scavenging abilities as 653.81 ± 20.76, 673.54 ± 32.34, and 675.29 ± 28.96 µg/g, respectively, and no significant difference was observed among the three (Figure S2b). Although the sample inoculated with 108 CFU/mL has a relatively high DPPH free radical scavenging ability, due to the considerable accumulation of organic acids, the flavor was peracid. Therefore, 107 CFU/mL was selected as the bacterial inoculum amount [33].
Finally, the conditions for the addition of Taiwan djulis were optimized, which were 0, 2.5, 5, and 7.5%, respectively, and their DPPH radical scavenging abilities were 331.89 ± 21.44, 621.46 ± 4.86, 673.54 ± 32.34, and 752.35 ± 29.29 µg/g, respectively (Figure S2c). Thus, Taiwan djulis fermented dairy products containing 7.5% have the highest DPPH free radical scavenging ability. Taiwan djulis is rich in polyphenols and flavonoids, such as betanin, and it is known that polyphenols and flavonoids have antioxidant capacity. Therefore, it can be speculated that Taiwan djulis fermented dairy products containing 7.5% would have the highest free DPPH, hence the base scavenging ability [35,36,37].
3.3. Antioxidant Capacity of Fermented Taiwan Djulis
Common antioxidant capacity measurement methods include DPPH and ABTS+ free radical scavenging ability. This method uses potassium persulfate to oxidize ABTS to generate ABTS+ free radicals. Therefore, 12–16 h of fermentation duration is necessary. It is mixed with potassium persulfate to generate ABTS+ free radicals. The DPPH free radical scavenging ability is more complicated in drug preparation, where the free radicals formed are less stable, affecting its reproducibility; therefore, ABTS+ free radicals, which are soluble in water and organic solvents, can determine the antioxidant capacity of hydrophilic and lipophilic compounds or samples are suitable. Therefore, this method is selected to determine the antioxidant capacity of samples in this experiment [32].
The results of the antioxidant capacity analysis of Taiwan djulis fermented products are shown in Table 1. The trend of ABTS+ free radical scavenging ability is the same as that of DPPH. The ABTS+ free radical scavenging abilities for 0, 5, and 7.5% Taiwan djulis fermented dairy products were 396.08 ± 1.67, 759.78 ± 23.44 and 771.52 ± 3.79 µg g−1, respectively. Thus, the 7.5% Taiwan djulis fermented dairy products had the highest ABTS+ free radical scavenging ability. It is speculated that the higher polyphenols and flavonoid content in 7.5% Taiwan djulis used to make the fermented dairy products provides the best ABTS+ free radical scavenging ability.

Table 1.
Comparison of antioxidant capacity, phenolic compounds, isoflavones, GABA and anti-nutritional compounds in fermented dairy products and Taiwan djulis fermented dairy products.
3.4. Evaluation of Functional Components in Fermented Taiwan Djulis
3.4.1. Content of Free Phenol, Bound Phenol, Total Phenol, and Total Flavonoids in Fermented Taiwan Djulis
The high antioxidant capacity of Taiwan djulis is related to its richness in phenolic compounds, which may exist in free form or in the bound form attached to the cell wall structure [38], Therefore, in this experiment, the contents of free phenol, bound phenol and total phenol were determined to ascertain their relationship with the antioxidant capacity of Taiwan djulis fermented dairy products.
The measurement of free phenol, bound phenol and total phenol of the samples showed (Table 1) that the matrix ratio and the total phenol had an increasing trend. There is a significant difference in Taiwan djulis to fermented dairy products, where the free phenol content increased by 2.5 times (fermented dairy product, FDP: 69.17 ± 2.03 mg GAE g−1; djulis fermented dairy product extract, DFDP: 169.90 ± 14.59 mg GAE g−1) and the fraction bound to phenol shown significantly decreased 13.65% (92.78 ± 0.88 mg GAE g−1). During the fermentation process, the bound phenolic compounds are converted due to bond breaking between the cell wall by the action of enzymes such as decarboxylase, esterase, hydrolase, and reductase, and by the metabolic action of the microorganisms. In the free form, phenolic compounds are more bioavailable, and the released free aglycones have the potential to enhance antioxidant activity [39]. Similar to previous reports on the fermentation of sourdough with lactic acid bacteria, the addition of Taiwan djulis also increased the content of phenolic compounds [40].
The seeds of djulis are rich in phenolic compounds, especially flavonoids, which promote health benefits and prevent diseases. The content of total flavonoids increased to five times that of the untreated group, and the results were in line with expectations (FDP: 0.636 ± 0.094 mg QE g−1; DFDP: 3.050 ± 0.026 mg QE g−1).
3.4.2. Content of Isoflavones in Fermented Taiwan Djulis
Isoflavones are found in various plants and are widely found in different foods. Isoflavone aglycone has high fat solubility, so it is easier to pass through the intestinal villi, increasing its bioavailability. However, isoflavone aglycone is less abundant in natural foods and more abundant in fermented foods because β-glucosidase produced by microorganisms decomposes isoflavones to produce isoflavone aglycones [41,42].
The isoflavone content analysis showed that the contents of daidzein, daidzin and genistin in Taiwan djulis fermented dairy products were higher than those in fermented dairy products without Taiwan djulis, but there was no significant difference in genistein content (Table 1). The literature points out that Lactobacillus can convert daidzin and genistin into daidzein and genistein, but the content of isoflavones in quinoa seeds is affected by its ecotype, growth climate, soil, sunlight and post-harvest storage conditions. Isoflavones have been shown to be converted into physiologically active substances through lactic acid bacteria. Daidzein and genistein are further converted into dihydrodaidzein and dihydrogenistein, which may be the possible reason for the lack of increase in the content of genistein.
3.4.3. GABA Content in Fermented Taiwan Djulis
GABA is widely distributed in nature. It is the primary inhibitory neurotransmitter in the mammalian brain and spinal cord. It is also essential in various physiological functions, such as regulating blood pressure, preventing chronic alcohol-related diseases, and inhibiting cancer cell proliferation [43,44,45,46]. In plants, GABA is a metabolic end product, which is irreversibly generated by the α-decarboxylation reaction of L-glutamic acid catalyzed by glutamic acid decarboxylase (GAD). GAD and GABA are produced from bacteria in higher organisms. The results showed that the GABA content of the lyophilized powders from 0 to 7.5% Taiwan djulis fermented dairy products were 6.52 ± 0.91 mg/g and 3.07 ± 0.94 mg/g, respectively (Table 1), it was similarly studied by Yu et al. (2020) [47], GABA-rich fermented milk had a beneficial effect on insomnia and could regulation of gut microbiota [47].
3.4.4. Content of Antinutritional Factor (Phytic Acid and Saponin) in Fermented Taiwan Djulis
Phytic acid, in which phosphorus is stored in plants, is often regarded as an anti-nutritional factor because it chelates divalent minerals such as calcium, iron, magnesium, zinc, copper, proteins, and starches, thereby reducing their bioavailability [48]. The phytic acid content in Taiwan djulis fermented dairy products was about 2.96 ± 1.39 µg/g (Table 1). The phytic acid is not only present in the outer layer of quinoa seeds, but also evenly distributed in the endosperm. Therefore, adding Taiwan djulis to fermented dairy products will increase the phytic acid content of the samples. Although the addition will also increase the phytate content of fermented dairy products, its content can be significantly reduced through soaking, germination and fermentation. Its degradation is better through processing methods that promote the activation of phytase, such as fermentation and cooking [46,49].
Djulis has a natural bitter layer called saponin, usually present in the outer layer, not only in the seeds but also in other parts, that protects it from birds and insects and adverse environmental conditions. As shown in Table 1, the saponin content in Taiwan djulis fermented dairy products is about 0.29 ± 0.27 µg g−1, and that in quinoa seeds is about 1.26% [50], showing that the saponin content tends to decrease after cleaning, heating and fermentation.
3.4.5. Component Analysis of Fermented Taiwan Djulis in Comparison with Commercially Available Fermented Milk
The weights of fermented dairy products and Taiwan djulis fermented dairy products were reduced by 87.58% and 83.33%, respectively, after freeze-drying. The general composition analysis was carried out with the freeze-dried powders.
The general components of the lyophilized powder of fermented dairy products were: moisture (3.04%); ash (5.74%); crude protein (13.75%); and crude fat (26.73%); while crude fiber was not detected. The general components of Taiwan djulis fermented dairy lyophilized powder were: moisture (4.48%); ash (4.57%); crude protein (11.94%); (crude fat 17.79%); and crude fiber (0.28%) (Table 1). In addition to the moisture and crude fiber content, the proportion between the two is higher in fermented dairy products.
According to the Taiwan Food Nutrients Database, the moisture content of high-fat fresh milk is about 85.8%, which is similar to the weight loss ratio after freeze-drying in the experiment. The crude fat and ash content of fermented dairy products are also the same as those in the database of full-fat milk. Milk powder data is similar, and volume dropped significantly.
3.4.6. Analysis of Amino Acid Content in Fermented Taiwan Djulis
The total amino acid content in fermented dairy products and Taiwan djulis fermented dairy products were similar (Table 2), 132.58 and 132.49 mg/g, respectively, but the distribution of each amino acid in the two was different. The content of cysteine, lysine, histidine, arginine, alanine, phenylalanine, isoleucine, leucin, and proline in the product was higher in the fermented products than that in Taiwan djulis fermented dairy products. In contrast, histidine in Taiwan djulis fermented dairy products is not found. In addition, threonine content is about 10 times higher than that of fermented dairy products. Both histidine and threonine cannot be synthesized by the human body and are essential amino acids, thus showing that the percentage of essential amino acids in Taiwan djulis fermented milk can be increased by fermentation.
Table 2.
Free amino acid content in fermented dairy products and Taiwan djulis fermented dairy products.
The initial pH value of Taiwan djulis fermented dairy products was 4.05, and the pH value dropped to 3.81 after being stored at 4 °C for 7 d. The pH value decreased, and then showed a gradual trend. The initial titratable acid of Taiwan djulis fermented dairy products was 0.77%, and the titratable acid increased to 0.91% after 7 d of storage at 4 °C. After 14 and 21 d of continuous storage, the titratable acid values were 0.93 and 0.98, respectively, which increased with time in the first 7 d, and showed a significant increasing trend, before becoming a plateau.
The initial total bacterial count of Taiwan djulis fermented dairy products was 1.8 × 109 CFU/mL, and the total bacterial count decreased to 1.5 × 109 CFU/mL after being stored at 4 °C for 7 days, and pH value decreased from 4.1 to 3.7 during storage period, respectively. The overall total bacterial counts showed a downward trend with time, especially in days 7–14, which may be attributed to the titratable acidity, pH value and low temperature.
3.5. Sensory Evaluation of Fermented Taiwan Djulis
The results of consumer sensory evaluation are shown in Figure 1. According to the results of the consumer acceptance test, the appearance, aroma, taste, flavor, aftertaste, and overall results of the commercially available yogurt are all relatively high, in the range of “slightly like” and “somewhat like”, while DFDP has the lowest score in each result, except for the appearance, which is “neither like nor dislike” all other indicators are “slightly disliked”. The consumer preference test results are consistent with the overall consumer acceptance test scores. The preference from high to low is as follows: COFDF, FDP, CFDP, and DFDP. This result shows that consumers’ acceptance of FDP is still higher than that of CFDP. It is speculated that the lower acceptance of DFDP is that compared with CFDP, in the DFDP the proportion of Taiwan djulis added in the product is as high as 7.5%. It can be seen from the appearance that the added amount is more significant than that of commercial grain yogurt. Furthermore, the grain of CFDP is added after yogurt is made, not before fermentation, while Taiwan djulis is added during the fermentation process, which increases the acidity of the fermented milk. Thus, the taste of Taiwan djulis is unacceptable to consumers compared with oatmeal, leading to a lower acceptance of DFDP.
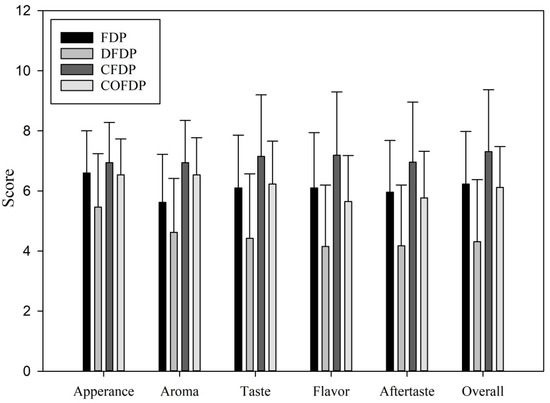
Figure 1.
Sensory evaluation results of fermented Taiwan djulis and other similar fermented dairy products. FDP: Fermented dairy products; DFDP: Taiwan djulis fermented dairy products; CFDP: Cereal fermented dairy products; COFDF: Commercial original fermented dairy products. The number of evaluations was 52, and the results were expressed as mean ± SD. Different letters indicate significant differences from each other.
3.6. Analysis of the Glucose Utilization-Promoting Activity of Fermented Taiwan Djulis in a Cellular Model
3.6.1. Cell Viability
The fermented milk products and Taiwan djulis fermented milk products were subjected to ethanol extraction, and the ethanol was entirely removed by concentrating under reduced pressure. It was then re-dissolved in DMSO to make the sample concentrations 25, 50, 100, and 200 µg/mL to test the viability in FL83B hepatocyte cell line. The extract from both FDP and Taiwan DFDP groups showed no significant cytotoxicity at a 200 µg/mL concentration, and the cell viability was higher than 95% (Figure S3a); Taiwan DFDP group at the concentration of 200 µg /mL, there was no significant cytotoxicity, and the cell viability was higher than 95% (Figure S3b). Based on these findings, groups with sample concentrations of 100 and 200 µg/mL were selected for subsequent glucose utilization experiments. Furthermore, the cell viability was investigated in response to guava extract on the FL83B hepatocyte cell line, which was not significantly cytotoxic even when the sample concentration was 200 µg/mL, and was consistent with the results of this experiment [45].
3.6.2. Effects of Ethanol Extracts from Fermented Dairy Products and Taiwan Djulis Fermented Dairy Products on Glucose Utilization in Mice FL83B Hepatocyte Cells
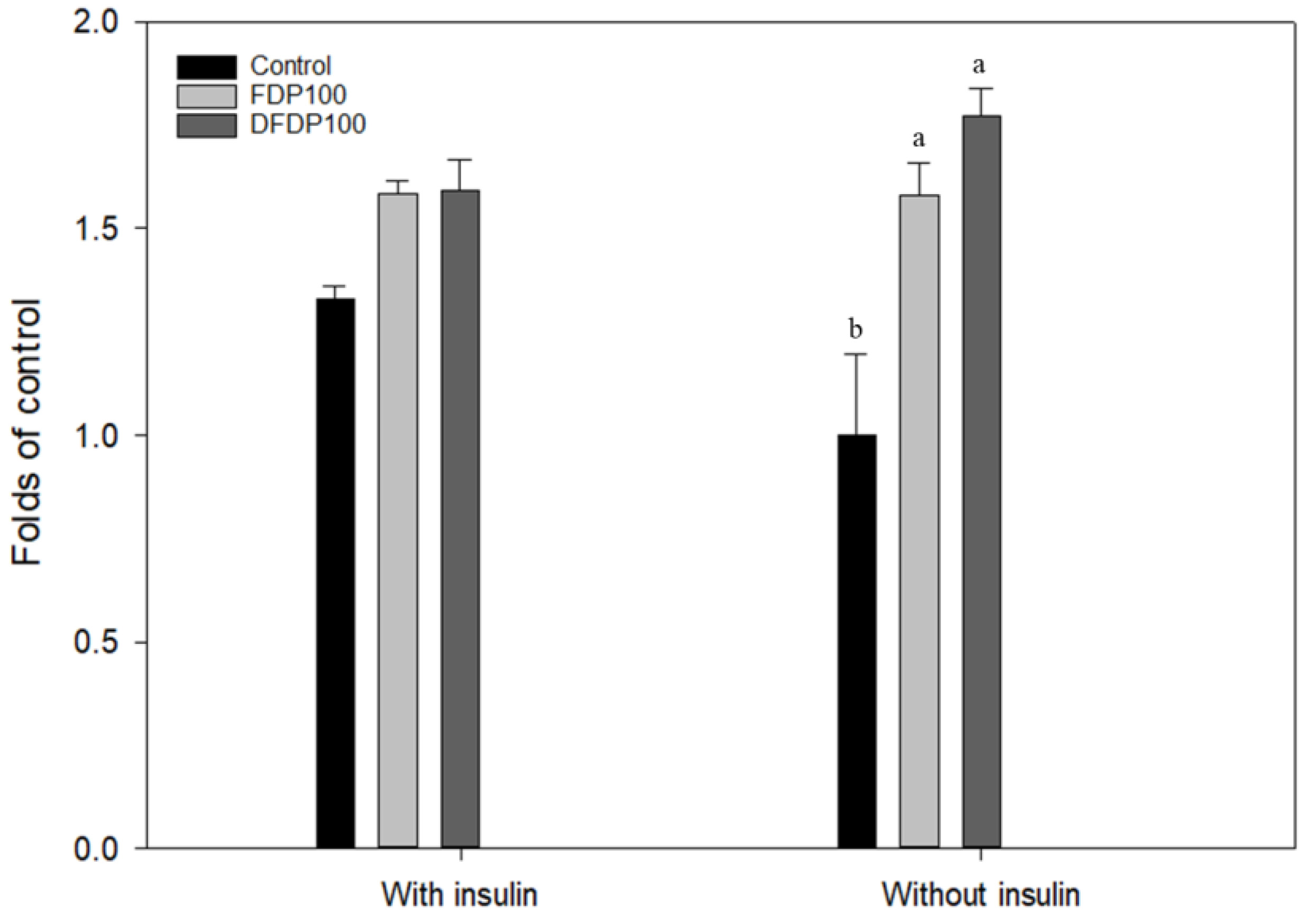
Ethanol extract (100 µg/mL) from fermented dairy products and Taiwan djulis fermented dairy products was selected and added to FL83B hepatocyte cell line to explore their glucose utilization. In the presence of 5 µg/mL insulin, the glucose utilization rate of the FL83B mouse liver cell line was significantly increased by 33% compared to that without insulin, suggesting that it was insulin sensitive (Figure 2). The ethanol extracts from both the fermented products had no significant difference in glucose utilization compared with the control group in an insulin-free environment. The glucose utilization rate can be increased to 1.6 times that of the control group, which is significantly different. However, there were no significant differences between DFDP and FDP groups on glucose utilization. Thus, both the ethanol extracts can potentially regulate the glucose utilization rate of FL83B hepatocyte cells and reduce glucose concentration, thereby reducing the phenomenon of insulin resistance, so follow-up experiments were continued at this concentration.
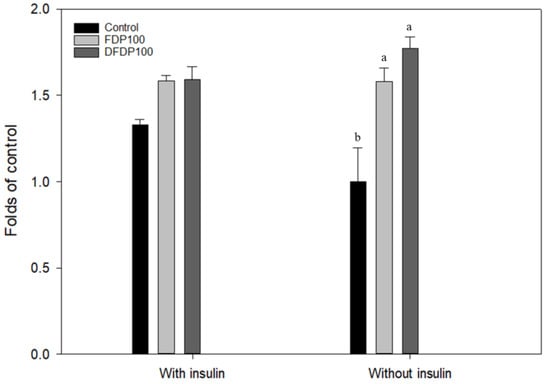
Figure 2.
Effects of ethanol extracts from fermented dairy products and Taiwan djulis fermented dairy products on glucose utilization in the liver cells of FL83B cell. Control: FL83B cell without treatment; FDP: Fermented dairy products ethanol extraction; DFDP: Taiwan djulis fermented dairy products ethanol extraction. The experiment was performed in triplicate and expressed as mean ± SD. Different letters indicate significant differences from each other.
3.6.3. Effects of Ethanol Extracts from Fermented Dairy Products and Taiwan Djulis Fermented Dairy Products on the Expression of Glucose Transporters in Mice FL83B Hepatocyte Cells
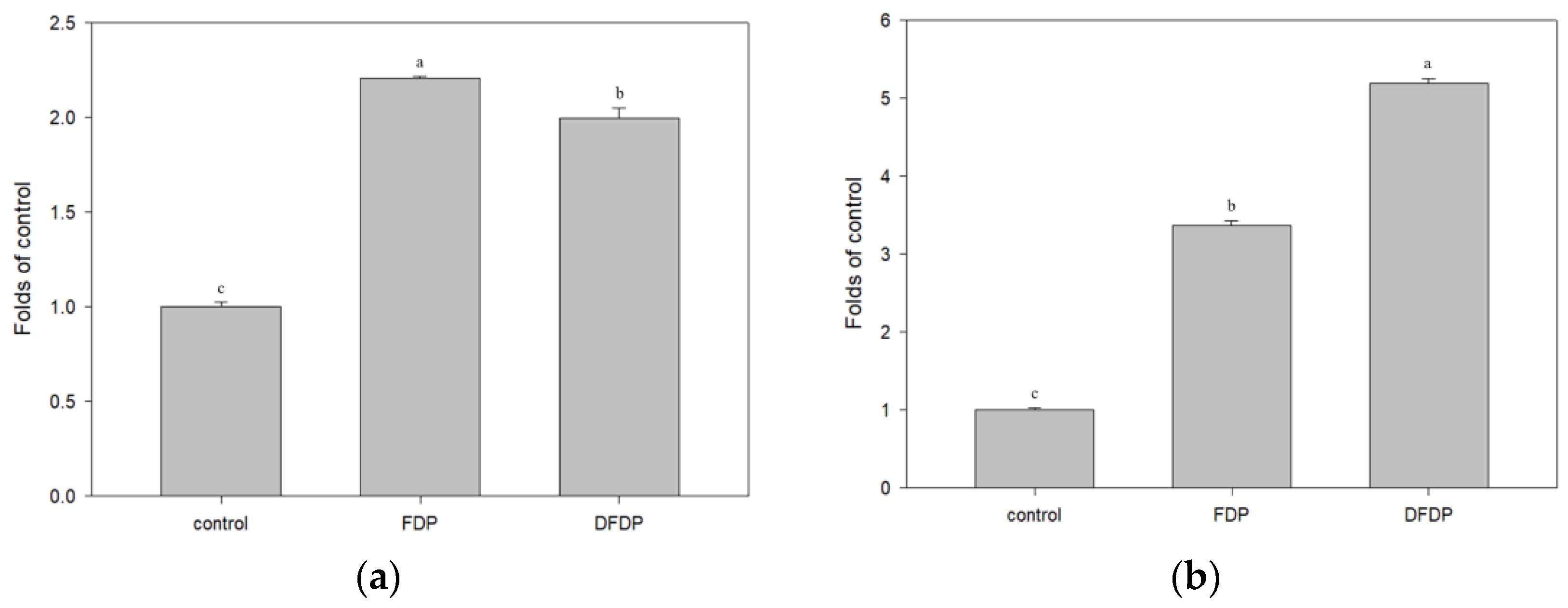
Ethanol extract (100 µg/mL) from fermented dairy products and Taiwan djulis fermented dairy products were added to the FL83B hepatocyte cell line to investigate the expression of glucose transporters. The addition of ethanol extract of both the fermented products in the presence of 5 µg/mL insulin led to a two-fold increase in the expression of glucose transporters, which is more than 10 times higher than that of the control group (Figure 3a). In the FL83B hepatocyte cell line, in an insulin-free environment, the addition of 100 µg/mL ethanol extract from fermented dairy products and Taiwan djulis fermented dairy products led to a 3.3 fold and 5.2 fold increase in the expression of glucose transporters, respectively, which was significantly different from the control group (Figure 3b). This indicated that Taiwan djulis fermented dairy products indeed have an effect of increasing the expression of glucose transporters in the FL83B hepatocyte cell line.
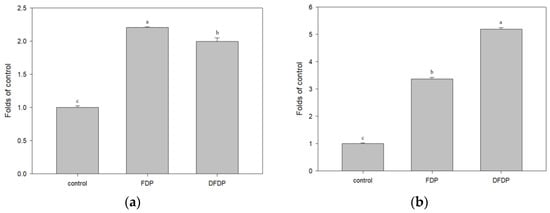
Figure 3.
The expression of glucose transporters in hepatocyte cells FL83B post-addition of ethanol extracts from fermented dairy products and Taiwan djulis fermented dairy products (a) in the presence of 5 µg/mL insulin and (b) in an insulin-free environment. The experiments were performed in triplicate and expressed as mean ± SD. Different letters indicate significant differences from each other.
4. Conclusions
This work highlights appropriate processing technology to add probiotics to Taiwan djulis, and combines Taiwan’s high-quality milk sources to develop functional Taiwan djulis fermented dairy products. The results demonstrated that 7.5% djulis, an inoculum of 107 CFU/mL, and fermenting at 40 °C for 18 h were the optimal fermentation conditions with Lactobacillus delbrueckii subsp. bulgaricus BCRC 10696 and Streptococcus salivarius subsp. thermophilus BCRC 14085. These conditions led to a high antioxidant capacity and phenolic content. Additionally, culture of hepatocyte cells, FL83B with fermented products, especially 100 µg/mL ethanol extract, can effectively improve glucose utilization and glucose transporter expression. These results indicated that the fermented dairy products have the potential to regulate glucose utilization and can be developed commercially to achieve high-value-added local agricultural products.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation8090423/s1, Table S1. Strain selection by DPPH, ABTS+ free radical scavenging capacity and content of phenolic compounds. Figure S1. Changes in antioxidant capacity (DPPH) of Taiwan djulis fermented with different lactic acid bacteria for 24 h (sample concentration 5 mg/mL). The experiments were performed in triplicate and expressed as mean ± standard deviation (SD). Figure S2. (a) DPPH free radical scavenging ability of samples at different fermentation tempera-tures (b) DPPH free radical scavenging ability of samples with different inoculation amounts (c) DPPH free radical scavenging ability with different proportions of Taiwan quinoa samples. The experiments were performed in triplicates and expressed as mean ± SD. Figure S3. Effects of ethanol extracts from fermented dairy products (a) and ethanol extracts from fermented dairy products of Taiwan quinoa (b) on the survival of FL83B mouse liver cell lines. The experiments were performed in triplicates and expressed as mean ± SD. Different letters indicate significant differences from each other.
Author Contributions
C.-Y.H. and C.-C.H. contributed equally to the manuscript writing and experimental execution; Y.-C.H. assisted with the cell and microbial experiment; K.-C.C. and C.-W.H. participated in manuscript writing and experimental design; C.-H.K. conducted the determination of amino acid contents; M.-H.C. provided experimental advice and manuscript writing. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Pingtung Agricultural Biotechnology Park Council of Agriculture, Executive Yuan, Taiwan (111AS-11.1.1-PT-f1) and (110AS-14.1.1-ST-aH).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available in the supplementary material of this article.
Acknowledgments
Thanks to the committee of Pingtung Agricultural Biotechnology Park Council of Agriculture, Executive Yuan for providing financial support and suggestions on the selection of Taiwan quinoa ingredients.
Conflicts of Interest
The authors do not declare any conflict of interest.
References
- Chu, C.-C.; Chen, S.-Y.; Chyau, C.-C.; Wang, S.-C.; Chu, H.-L.; Duh, P.-D. Djulis (Chenopodium formosanum) and Its Bioactive Compounds Protect Human Lung Epithelial A549 Cells from Oxidative Injury Induced by Particulate Matter via Nrf2 Signaling Pathway. Molecules 2021, 27, 253. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.-C.; Chen, S.-Y.; Chyau, C.-C.; Fu, Z.-H.; Liu, C.-C.; Duh, P.-D. Protective Effect of Djulis (Chenopodium formosanum) and Its Bioactive Compounds against Carbon Tetrachloride-Induced Liver Injury, in Vivo. J. Funct. Foods 2016, 26, 585–597. [Google Scholar] [CrossRef]
- International Year of Quinoa, 2013: Resolution/. Available online: https://www.fao.org/quinoa-2013/mobile/home/en/ (accessed on 17 July 2022).
- Tsai, P.-J.; Sheu, C.-H.; Wu, P.-H.; Sun, Y.-F. Thermal and PH Stability of Betacyanin Pigment of Djulis (Chenopodium formosanum) in Taiwan and Their Relation to Antioxidant Activity. J. Agric. Food Chem. 2010, 58, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Jiao, J.; Zhang, W.; Xu, J.; Wan, Z.; Zhang, W.; Gao, X.; Qin, L. Dietary Fiber Prevents Obesity-Related Liver Lipotoxicity by Modulating Sterol-Regulatory Element Binding Protein Pathway in C57BL/6J Mice Fed a High-Fat/Cholesterol Diet. Sci. Rep. 2015, 5, 15256. [Google Scholar] [CrossRef]
- Yao, Y.; Suo, T.; Andersson, R.; Cao, Y.; Wang, C.; Lu, J.; Chui, E. Dietary Fibre for the Prevention of Recurrent Colorectal Adenomas and Carcinomas. Cochrane Database Syst. Rev. 2017, 2017, CD003430. [Google Scholar] [CrossRef]
- Chen, S.H.; Chu, C.-C.; Lin, Y.-C.; Duh, P.-D. Djulis (Chenopodium Formosanum) and Its Bioactive Compounds for Management of Hyperlipidemia and Hyperglycemia in High-Fat Diet-Fed Mice. J. Food Nutr. Res 2019, 7, 452–457. [Google Scholar] [CrossRef]
- Chyau, C.-C.; Chu, C.-C.; Chen, S.-Y.; Duh, P.-D. The Inhibitory Effects of Djulis (Chenopodium formosanum) and Its Bioactive Compounds on Adipogenesis in 3T3-L1 Adipocytes. Molecules 2018, 23, 1780. [Google Scholar] [CrossRef]
- Nafees, S.; Rashid, S.; Ali, N.; Hasan, S.K.; Sultana, S. Rutin Ameliorates Cyclophosphamide Induced Oxidative Stress and Inflammation in Wistar Rats: Role of NFκB/MAPK Pathway. Chem. Biol. Interact. 2015, 231, 98–107. [Google Scholar] [CrossRef]
- Sikder, K.; Kesh, S.B.; Das, N.; Manna, K.; Dey, S. The High Antioxidative Power of Quercetin (Aglycone Flavonoid) and Its Glycone (Rutin) Avert High Cholesterol Diet Induced Hepatotoxicity and Inflammation in Swiss Albino Mice. Food Funct. 2014, 5, 1294–1303. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Nam, S.-Y.; Hong, S.-W.; Kim, M.-J.; Jeong, H.-J.; Kim, H.-M. Protective Effects of Rutin through Regulation of Vascular Endothelial Growth Factor in Allergic Rhinitis. Am. J. Rhinol. Allergy 2015, 29, e87–e94. [Google Scholar] [CrossRef]
- Hafez, M.M.; Al-Harbi, N.O.; Al-Hoshani, A.R.; Al-Hosaini, K.A.; AlShrari, S.D.; AlRejaie, S.S.; Sayed-Ahmed, M.M.; Al-Shabanah, O.A. Hepato-Protective Effect of Rutin via IL-6/STAT3 Pathway in CCl4-Induced Hepatotoxicity in Rats. Biol. Res. 2015, 48, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of Gaba (γ-Aminobutyric Acid) by Microorganisms: A Review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef]
- Rashmi, D.; Zanan, R.; John, S.; Khandagale, K.; Nadaf, A. Chapter 13-γ-Aminobutyric Acid (GABA): Biosynthesis, Role, Commercial Production, and Applications. Stud. Nat. Prod. Chem. 2018, 57, 413–452. [Google Scholar]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Hu, G.; Zheng, Y.; Liu, Z.; Xiao, Y.; Deng, Y.; Zhao, Y. Effects of High Hydrostatic Pressure, Ultraviolet Light-C, and Far-Infrared Treatments on the Digestibility, Antioxidant and Antihypertensive Activity of α-Casein. Food Chem. 2017, 221, 1860–1866. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Y.; Chen, Y.-W.; Hou, C.-Y. Antioxidant and Antibacterial Activity of Seven Predominant Terpenoids. Int. J. Food Prop. 2019, 22, 230–238. [Google Scholar] [CrossRef]
- Shih, M.-K.; Lai, Y.-H.; Lin, C.-M.; Chen, Y.-W.; Hou, Z.-T.; Hou, C.-Y. A Novel Application of Terpene Compound α-Pinene for Alternative Use of Sulfur Dioxide-Free White Wine. Int. J. Food Prop. 2020, 23, 520–532. [Google Scholar] [CrossRef]
- Hur, J.; Nguyen, T.T.H.; Park, N.; Kim, J.; Kim, D. Characterization of quinoa (Chenopodium quinoa) fermented by Rhizopus oligosporus and its bioactive properties. AMB Express 2018, 8, 143. [Google Scholar] [CrossRef]
- Wang, Y.H.; Kuo, C.H.; Lee, C.L.; Kuo, W.C.; Tsai, M.L.; Sun, P.P. Enzyme-assisted aqueous extraction of cobia liver oil and protein hydrolysates with antioxidant activity. Catalysts 2020, 10, 1323. [Google Scholar] [CrossRef]
- Gnanou, J.; Srinivas, S.; Kurpad, A. Automated derivatization with o-phthalaldehyde for the estimation of amino acids in plasma using reversed-phase high performance liquid chromatography. Indian J. Biochem. Biophys. 2004, 41, 322–325. [Google Scholar] [CrossRef]
- Gao, Y.; Shang, C.; Maroof, M.S.; Biyashev, R.M.; Grabau, E.A.; Kwanyuen, P.; Burton, W.J.; Buss, G.R. A modified colorimetric method for phytic acid analysis in soybean. Crop Sci. 2007, 47, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Bondoc, K.G.V.; Lee, H.; Cruz, L.J.; Lebrilla, C.B.; Juinio-Meñez, M.A. Chemical fingerprinting and phylogenetic mapping of saponin congeners from three tropical holothurian sea cucumbers. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2013, 166, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Wang, Y.C.; Hsieh, C.C.; Lu, H.C.; Chiang, W.D. Guava (Psidium guajava Linn.) leaf extract promotes glucose uptake and glycogen accumulation by modulating the insulin signaling pathway in high-glucose-induced insulin-resistant mouse FL83B cells. Process Biochem. 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Kuo, H.-C.; Kwong, H.K.; Chen, H.-Y.; Hsu, H.-Y.; Yu, S.-H.; Hsieh, C.-W.; Lin, H.-W.; Chu, Y.-L.; Cheng, K.-C. Enhanced Antioxidant Activity of Chenopodium formosanum Koidz. by Lactic Acid Bacteria: Optimization of Fermentation Conditions. PLoS ONE 2021, 16, e0249250. [Google Scholar] [CrossRef]
- Xu, L.-N.; Guo, S.; Zhang, S. Effects of Solid-State Fermentation with Three Higher Fungi on the Total Phenol Contents and Antioxidant Properties of Diverse Cereal Grains. FEMS Microbiol. Lett. 2018, 365, fny163. [Google Scholar] [CrossRef]
- Dudonné, S.; Vitrac, X.; Coutière, P.; Woillez, M.; Mérillon, J.-M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- María, E.-T.; María, L.J.; Inés, R.; Laura, S.; de las Rivas, B.; Rosario, M. A Lactobacillus Plantarum Esterase Active on a Broad Range of Phenolic Esters. Appl. Environ. Microbiol. 2015, 81, 3235–3242. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food Phenolics and Lactic Acid Bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef]
- Faulds, C.B. What Can Feruloyl Esterases Do for Us? Phytochem. Rev. 2010, 9, 121–132. [Google Scholar] [CrossRef]
- Campanella, D.; Rizzello, C.G.; Fasciano, C.; Gambacorta, G.; Pinto, D.; Marzani, B.; Scarano, N.; DeAngelis, M.; Gobbetti, M. Exploitation of Grape Marc as Functional Substrate for Lactic Acid Bacteria and Bifidobacteria Growth and Enhanced Antioxidant Activity. Food Microbiol. 2017, 65, 25–35. [Google Scholar] [CrossRef]
- Shah, P.; Modi, H.A. Comparative Study of DPPH, ABTS and FRAP Assays for Determination of Antioxidant Activity. Int. J. Res. Appl. Sci. Eng. Technol. 2015, 3, 636–641. [Google Scholar]
- Matejekov, Z.; Liptkov, D.; Spodniakov, S.; Valk, u. Characterization of the Growth of in Milk in Dependence on Temperature. Acta Chim. Slovaca 2016, 9, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Aryana, K.J.; Olson, D.W. A 100-Year Review: Yogurt and Other Cultured Dairy Products. J. Dairy Sci. 2017, 100, 9987–10013. [Google Scholar] [CrossRef] [PubMed]
- Ravipati, A.S.; Zhang, L.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Bartlett, J.; Smith, P.T.; Shanmugam, K.; Münch, G.; Wu, M.J.; et al. Antioxidant and Anti-Inflammatory Activities of Selected Chinese Medicinal Plants and Their Relation with Antioxidant Content. BMC Complement. Altern. Med. 2012, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-W.; Cheng, M.-C.; Chen, B.-Y.; Wang, C.-Y. Effects of High Pressure Extraction on the Extraction Yield, Phenolic Compounds, Antioxidant and Anti-Tyrosinase Activity of Djulis Hull. J. Food Sci. Technol. 2019, 56, 4016–4024. [Google Scholar] [CrossRef]
- Zhang, Q.; Xing, B.; Sun, M.; Zhou, B.; Ren, G.; Qin, P. Changes in Bio-Accessibility, Polyphenol Profile and Antioxidants of Quinoa and Djulis Sprouts during in Vitro Simulated Gastrointestinal Digestion. Food Sci. Nutr. 2020, 8, 4232–4241. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Melini, F.; Melini, V. Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa. Fermentation 2021, 7, 20. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, H.; Hao, S.; Pan, D.; Wang, G.; Yu, W. Evaluation of Phenolic Composition and Antioxidant Properties of Different Varieties of Chinese Citrus. Food Chem. 2021, 364, 130413. [Google Scholar] [CrossRef]
- Hsiao, Y.-H.; Ho, C.-T.; Pan, M.-H. Bioavailability and Health Benefits of Major Isoflavone Aglycones and Their Metabolites. J. Funct. Foods 2020, 74, 104164. [Google Scholar] [CrossRef]
- Shang, X.; Dou, Y.; Zhang, Y.; Tan, J.-N.; Liu, X.; Zhang, Z. Tailor-Made Natural Deep Eutectic Solvents for Green Extraction of Isoflavones from Chickpea (Cicer Arietinum L.) Sprouts. Ind. Crops Prod. 2019, 140, 111724. [Google Scholar] [CrossRef]
- Oh, S.-H.; Moon, Y.-J.; Oh, C.-H. γ-Aminobutyric Acid (GABA) Content of Selected Uncooked Foods. Prev. Nutr. Food Sci. 2003, 8, 75–78. [Google Scholar] [CrossRef]
- Oh, C.-H.; Oh, S.-H. Effects of Germinated Brown Rice Extracts with Enhanced Levels of GABA on Cancer Cell Proliferation and Apoptosis. J. Med. Food 2004, 7, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, A.; Sato, K.; Park, E.Y.; Nakamura, Y.; Ohtsuki, K. Control of Amylase and Protease Activities in a Phytase Preparation by Ampholyte-Free Preparative Isoelectric Focusing for Unrefined Cereal-Containing Bread. J. Funct. Foods 2012, 4, 513–519. [Google Scholar] [CrossRef]
- Oh, S.-H.; Soh, J.-R.; Cha, Y.-S. Germinated Brown Rice Extract Shows a Nutraceutical Effect in the Recovery of Chronic Alcohol-Related Symptoms. J. Med. Food 2003, 6, 115–121. [Google Scholar] [CrossRef]
- Yu, L.; Han, X.; Cen, S.; Duan, H.; Feng, S.; Xue, Y.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; et al. Beneficial effect of GABA-rich fermented milk on insomnia involving regulation of gut microbiota. Microbiol. Res. 2020, 233, 126409. [Google Scholar] [CrossRef]
- Mohan, V.R.; Tresina, P.S.; Daffodil, E.D. Antinutritional Factors in Legume Seeds: Characteristics and Determination. Agric. Sci. 2016, 211–220. [Google Scholar] [CrossRef]
- Khattab, R.Y.; Arntfield, S.D. Nutritional Quality of Legume Seeds as Affected by Some Physical Treatments 2. Antinutritional Factors. LWT Food Sci. Technol. 2009, 42, 1113–1118. [Google Scholar] [CrossRef]
- Lim, J.G.; Park, H.-M.; Yoon, K.S. Analysis of Saponin Composition and Comparison of the Antioxidant Activity of Various Parts of the Quinoa Plant (Chenopodium Quinoa Willd.). Food Sci. Nutr. 2020, 8, 694–702. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).