
Biodiversity of Lactic Acid Bacteria in Traditional Fermented Foods in Yunnan Province, China, and Comparative Genomics of Lactobacillus plantarum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of LAB
2.2. Diversity Analysis
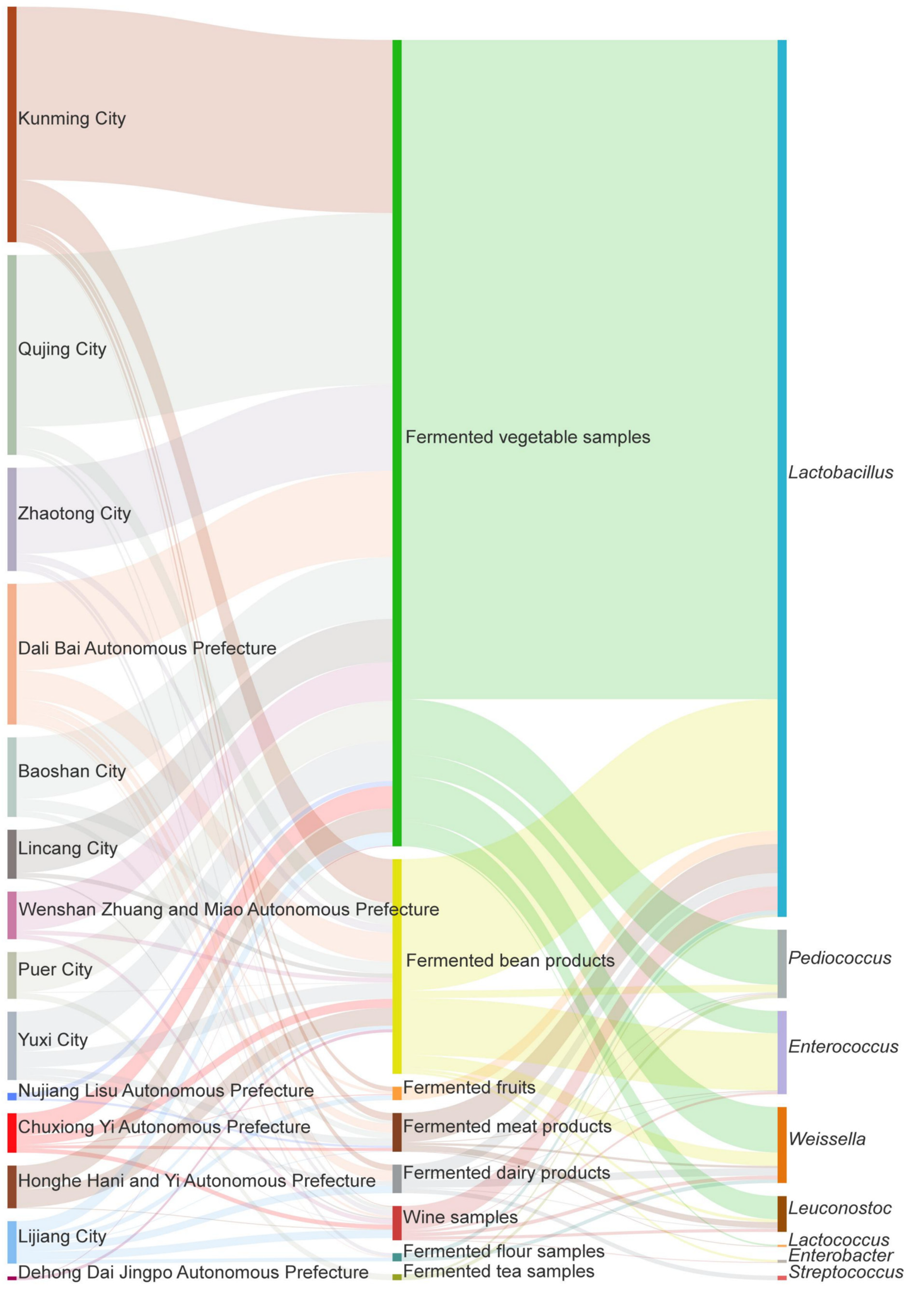
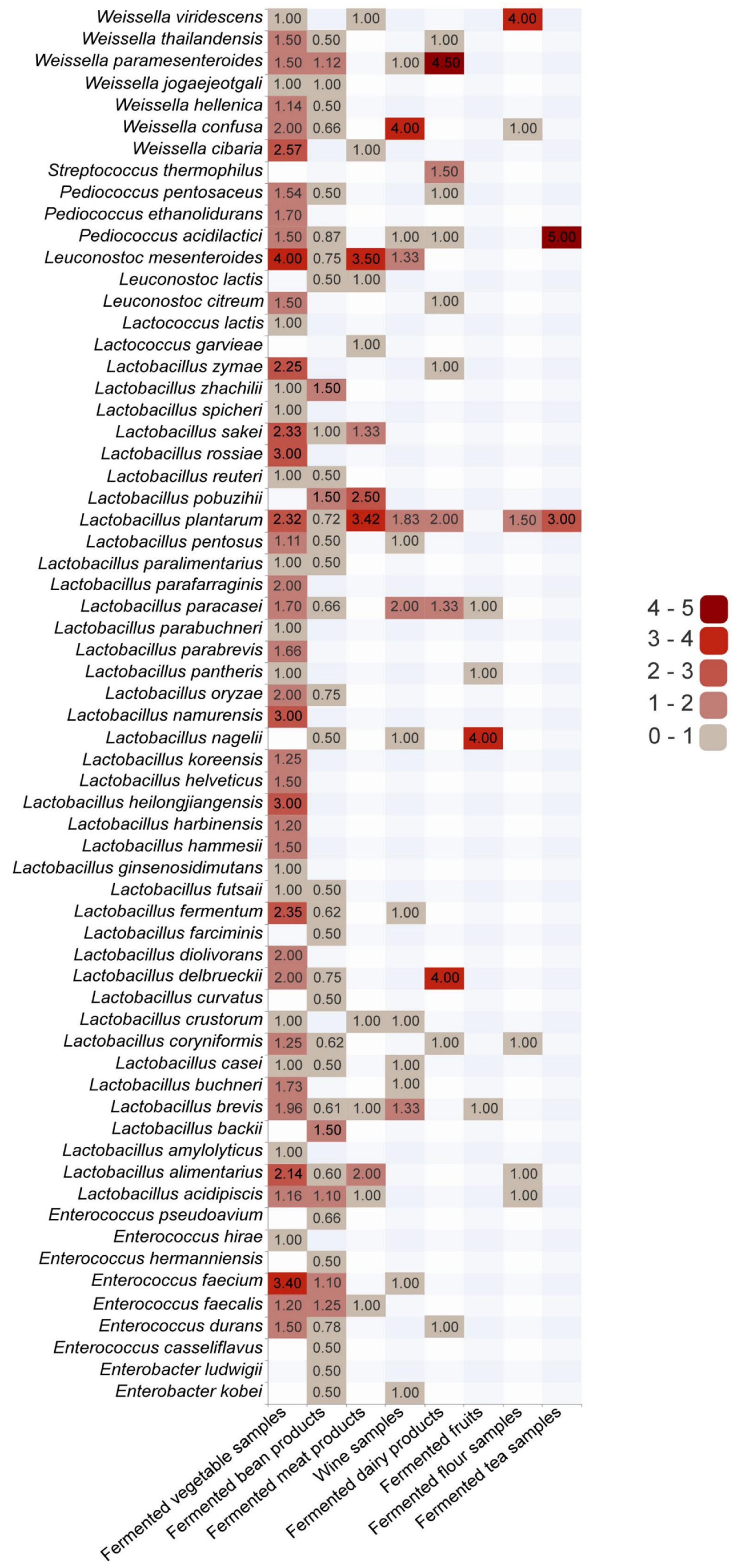
2.2.1. Distribution of LAB in Different Fermented Foods
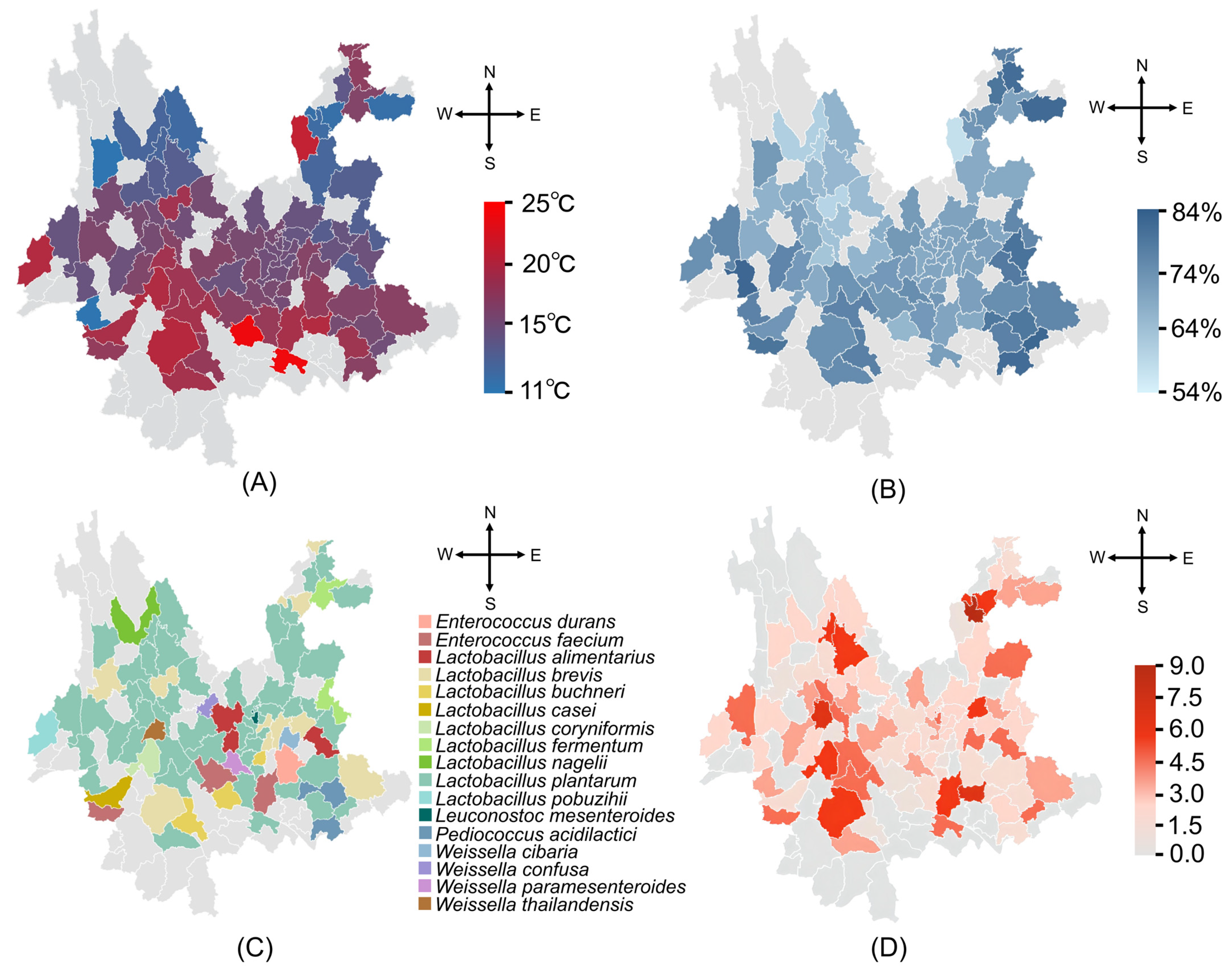
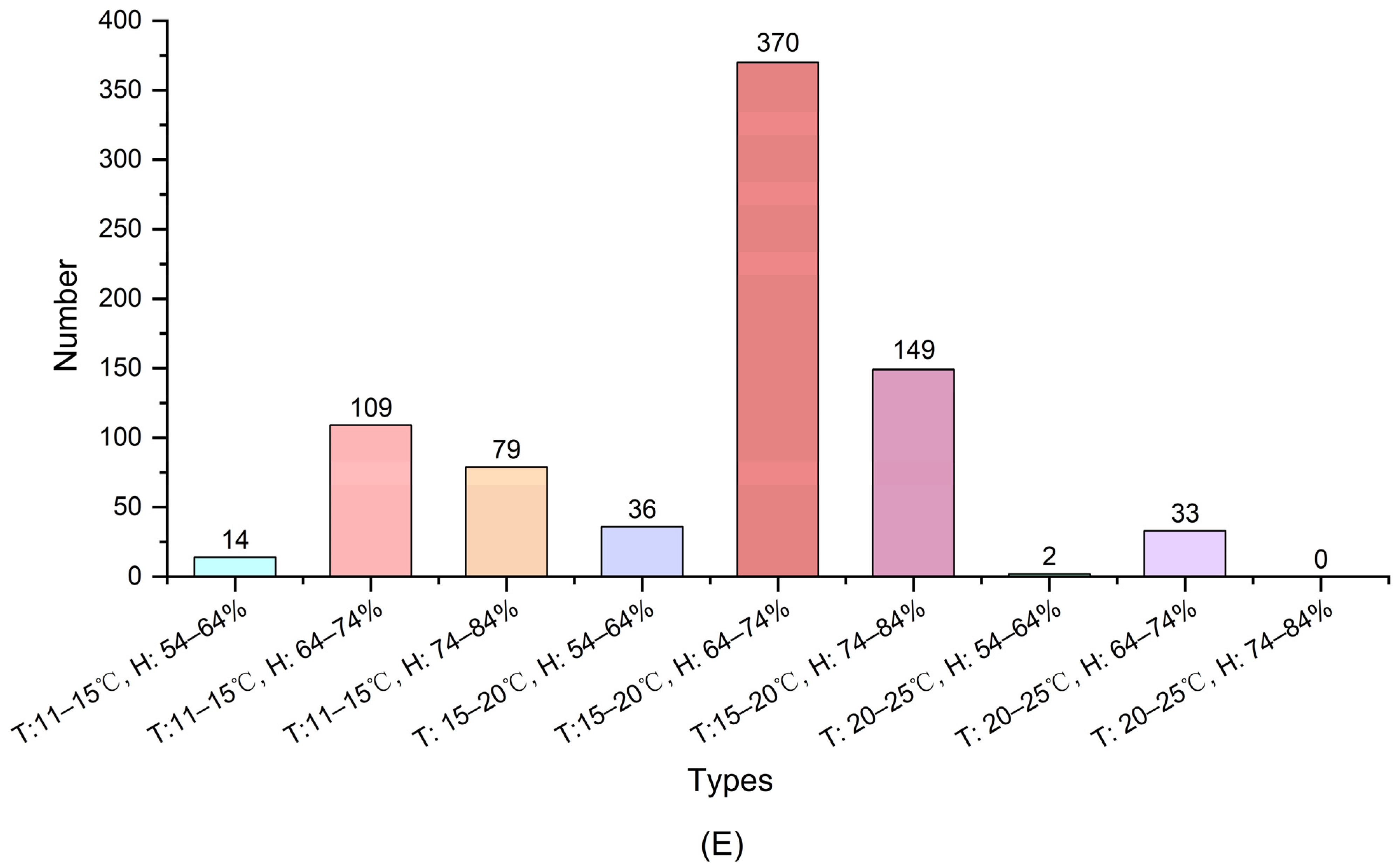
2.2.2. Distribution of LAB in Different Regions of Yunnan
2.3. Comparative Genomics Analysis
2.4. Statistical Analyses
3. Results
3.1. Diversity of Distribution of LAB
3.1.1. Species and Numbers of Culturable LAB
3.1.2. Dominant LAB in Different Types of Fermented Foods
3.1.3. Distribution of LAB in Different Regions of Yunnan
3.2. Genetic Diversity of L. plantarum
3.2.1. General Genomic Features
3.2.2. ANI Analyses of L. plantarum
3.2.3. Pan-Genome and Core Genes of L. plantarum
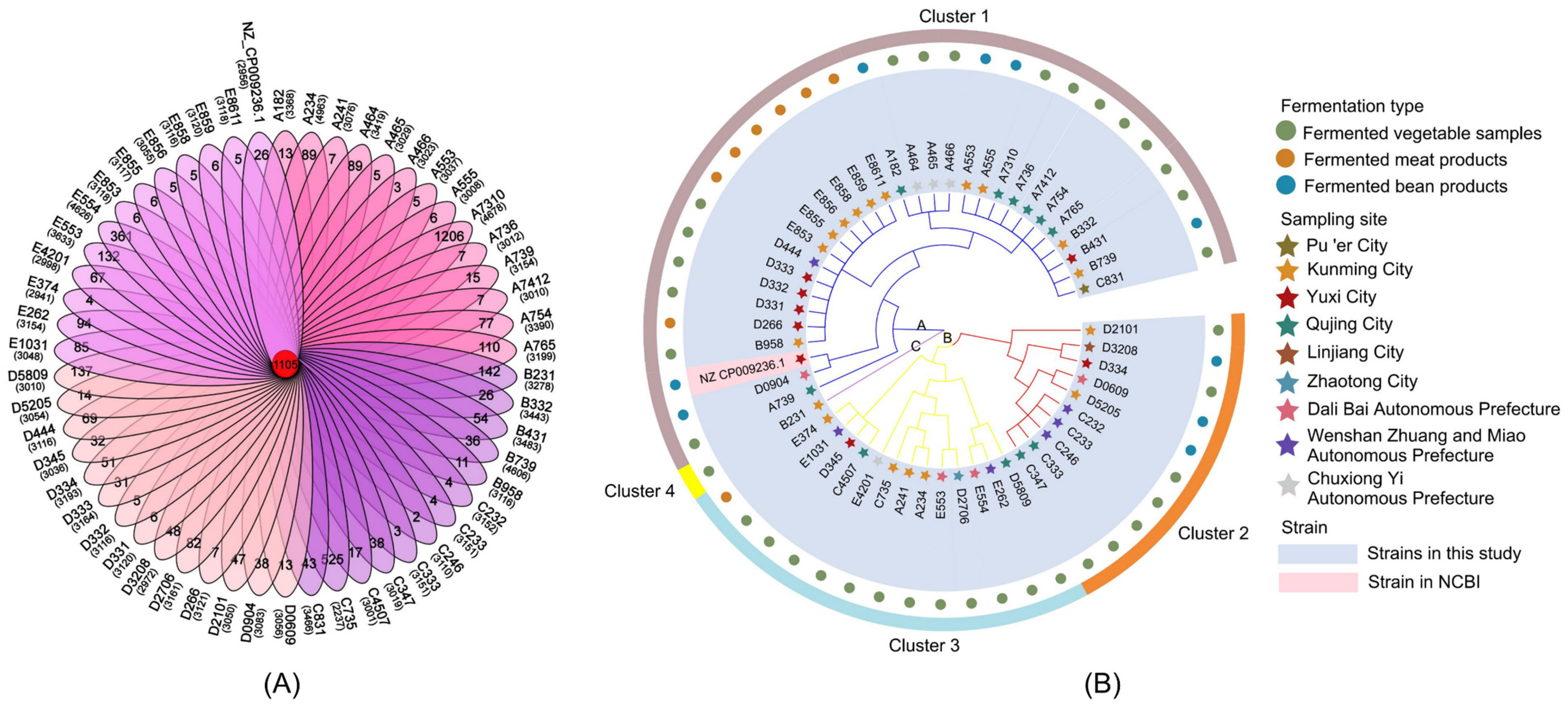
3.2.4. Homologous Genes and Phylogenetic Analyses
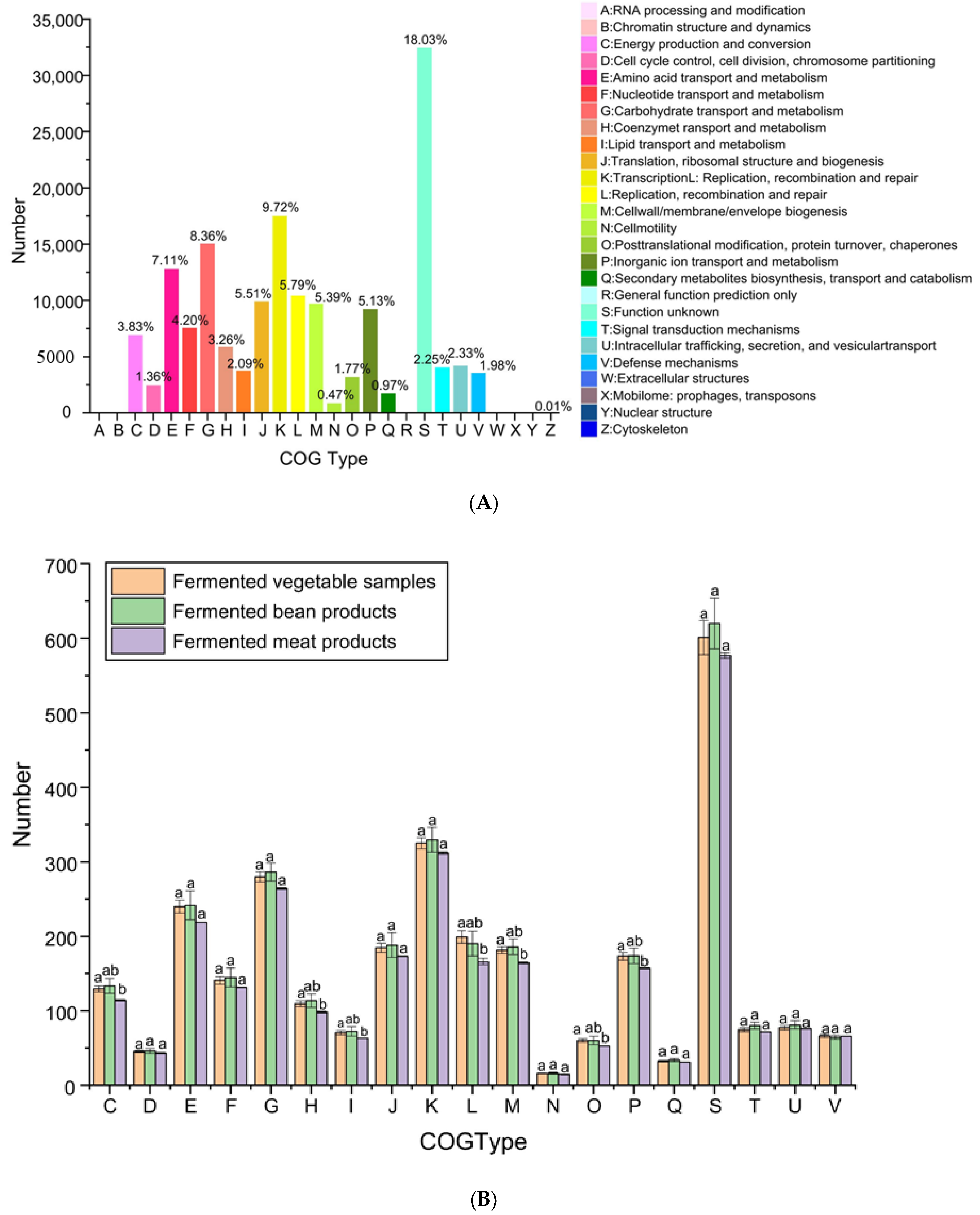
3.2.5. Functional Annotation Analysis of COG Database
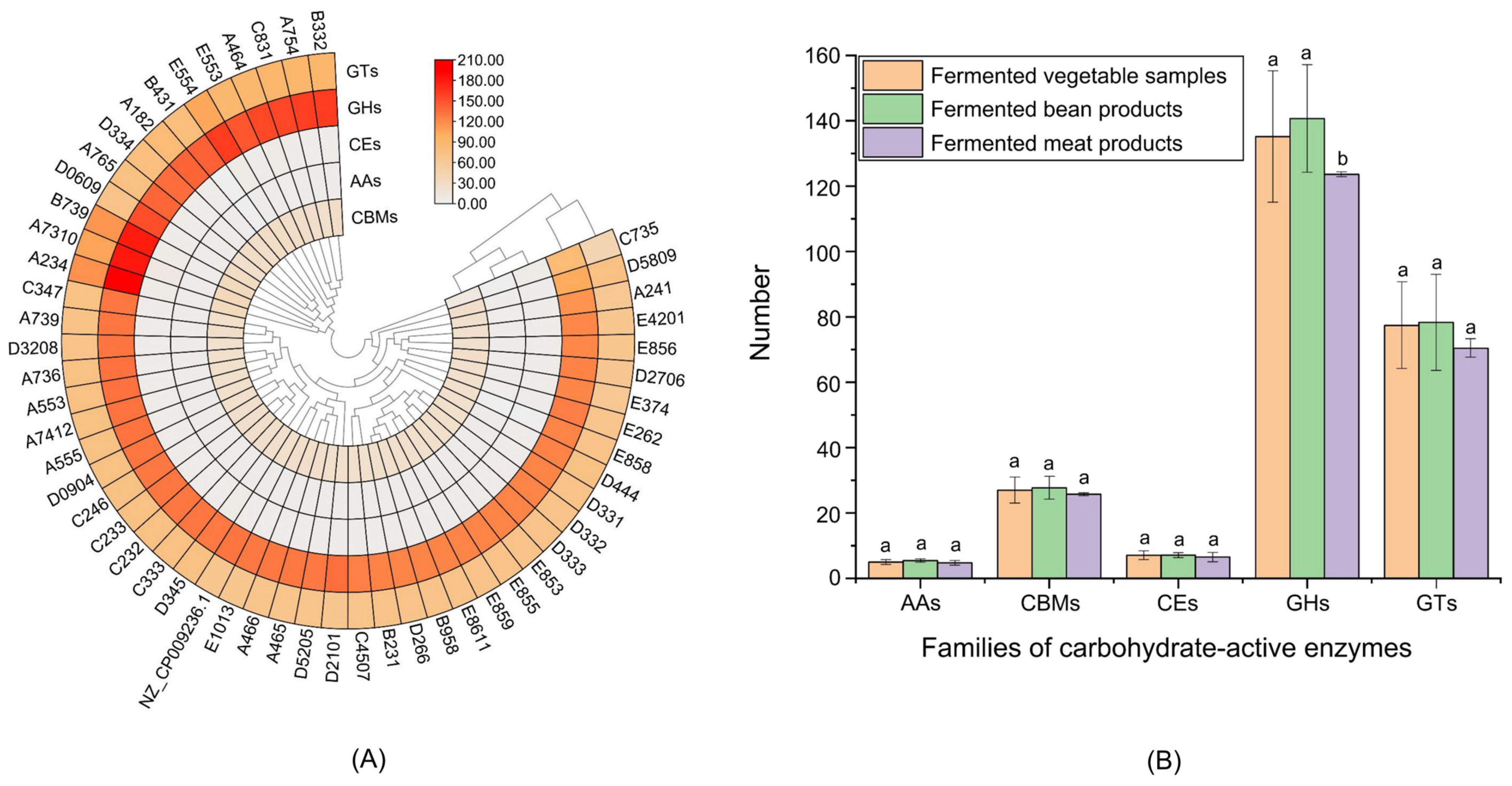
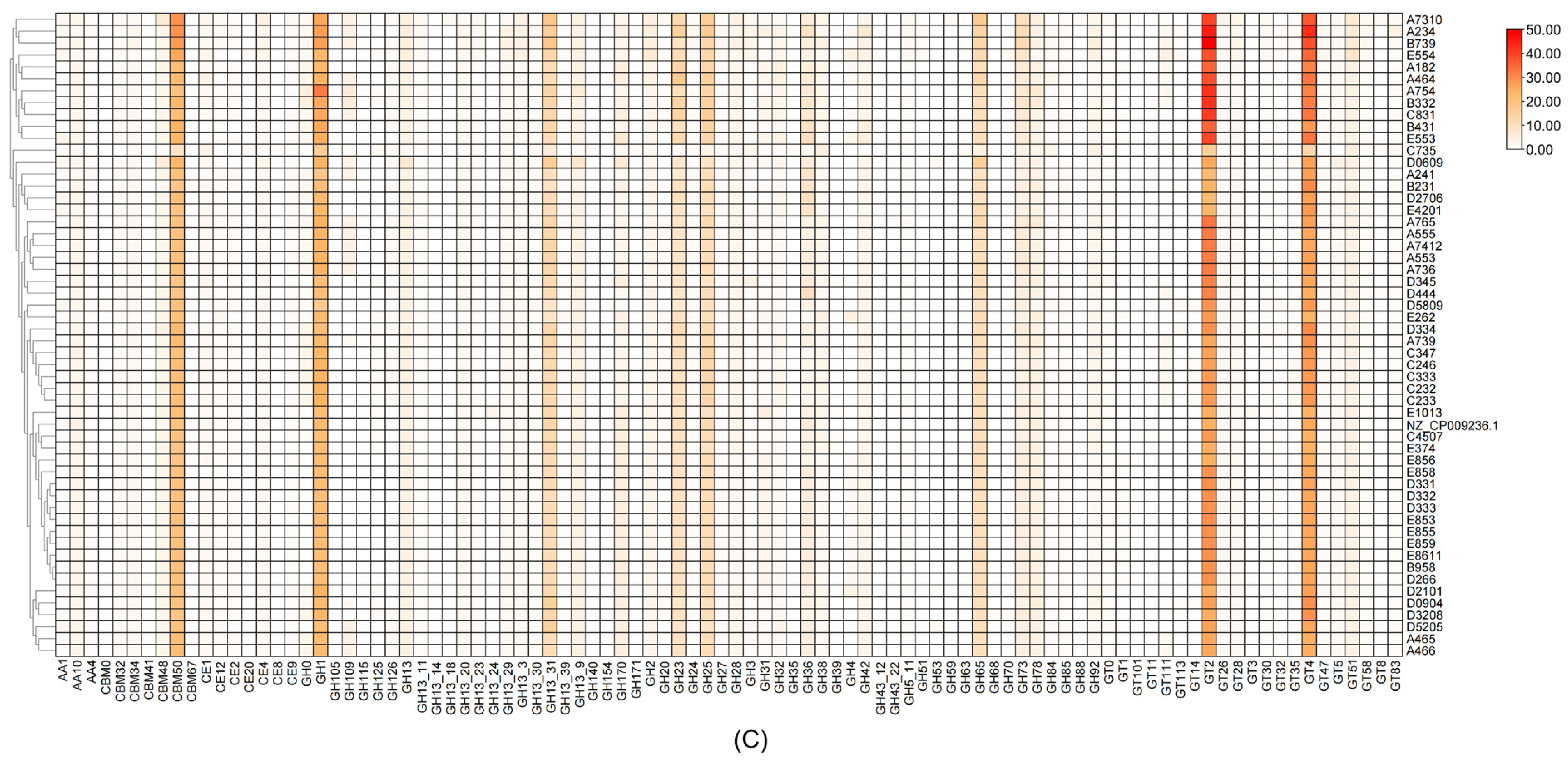
3.2.6. Carbohydrate Enzyme Analysis of L. plantarum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Souza, E.L.; de Oliveira, K.A.R.; de Oliveira, M.E.G. Influence of lactic acid bacteria metabolites on physical and chemical food properties. Curr. Opin. Food Sci. 2023, 49, 100981. [Google Scholar] [CrossRef]
- Jaffar, N.S.; Jawan, R.; Chong, K.P. The potential of lactic acid bacteria in mediating the control of plant diseases and plant growth stimulation in crop production—A mini review. Front. Plant Sci. 2023, 13, 1047945. [Google Scholar] [CrossRef] [PubMed]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the gut microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Barcenilla, C.; Ducic, M.; Lopez, M.; Prieto, M.; Alvarez-Ordonez, A. Application of lactic acid bacteria for the biopreservation of meat products: A systematic review. Meat Sci. 2022, 183, 108661. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Xu, R.; Zhang, S.; Wang, Y.; Hu, B.; Ao, X.; Li, Q.; Li, J.; Hu, K.; Yang, Y.; et al. Antifungal mechanisms and application of lactic acid bacteria in bakery products: A Review. Front. Microbiol. 2022, 13, 924398. [Google Scholar] [CrossRef]
- Reque, P.M.; Brandelli, A. Encapsulation of probiotics and nutraceuticals: Applications in functional food industry. Trends Food Sci. Technol. 2021, 114, 1–10. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Wen, R.; Chen, Q.; Kong, B. Role of lactic acid bacteria in flavor development in traditional Chinese fermented foods: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2741–2755. [Google Scholar] [CrossRef]
- Un-Nisa, A.; Khan, A.; Zakria, M.; Siraj, S.; Ullah, S.; Tipu, M.K.; Ikram, M.; Kim, M.O. Updates on the role of probiotics against different health issues: Focus on Lactobacillus. Int. J. Mol. Sci. 2023, 24, 142. [Google Scholar] [CrossRef]
- Ye, Z.; Shang, Z.; Zhang, S.; Li, M.; Zhang, X.; Ren, H.; Hu, X.; Yi, J. Dynamic analysis of flavor properties and microbial communities in Chinese pickled chili pepper (Capsicum frutescens L.): A typical industrial-scale natural fermentation process. Food Res. Int. 2022, 153, 110952. [Google Scholar] [CrossRef]
- Liu, C.-J.; Gong, F.-M.; Li, X.-R.; Li, H.-Y.; Zhang, Z.-H.; Feng, Y.; Nagano, H. Natural populations of lactic acid bacteria in douchi from Yunnan Province, China. J. Zhejiang Univ.-Sci. B 2012, 13, 298–306. [Google Scholar] [CrossRef]
- Liu, W.; Sun, Z.; Zhang, J.; Gao, W.; Wang, W.; Wu, L.; Sun, T.; Chen, W.; Liu, X.; Zhang, H. Analysis of microbial composition in acid whey for dairy fan making in Yunnan by conventional method and 16S rRNA sequencing. Curr. Microbiol. 2009, 59, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Cen, S.; Yu, L.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q. Niche-Specific adaptive evolution of Lactobacillus plantarum strains isolated from human feces and paocai. Front. Cell. Infect. Microbiol. 2021, 10, 615876. [Google Scholar] [CrossRef]
- Cen, S.; Yin, R.; Mao, B.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Comparative genomics shows niche-specific variations of Lactobacillus plantarum strains isolated from human, Drosophila melanogaster, vegetable and dairy sources. Food Biosci. 2020, 35, 100581. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, C.; Yu, L.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q. Phylogenetic and comparative genomic analysis of Lactobacillus fermentum strains and the key genes related to their intestinal anti-inflammatory effects. Engineering 2022, 17, 170–182. [Google Scholar] [CrossRef]
- Mendoza, R.M.; Kim, S.H.; Vasquez, R.; Hwang, I.-C.; Park, Y.-S.; Paik, H.-D.; Moon, G.-S.; Kang, D.-K. Bioinformatics and its role in the study of the evolution and probiotic potential of lactic acid bacteria. Food Sci. Biotechnol. 2022, 32, 389–412. [Google Scholar] [CrossRef]
- Wang, S.; Yang, B.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Chen, W. Comparative genomics analysis of Lactobacillus ruminis from different niches. Genes 2020, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Song, Y.; Ren, Y.; Qing, Y.; Liu, W.; Sun, Z. Genome-level comparisons provide insight into the phylogeny and metabolic diversity of species within the genus Lactococcus. BMC Microbiol. 2017, 17, 213. [Google Scholar] [CrossRef]
- Martino, M.E.; Bayjanov, J.R.; Caffrey, B.E.; Wels, M.; Joncour, P.; Hughes, S.; Gillet, B.; Kleerebezem, M.; van Hijum, S.A.F.T.; Leulier, F. Nomadic lifestyle of Lactobacillus plantarum revealed by comparative genomics of 54 strains isolated from different habitats. Environ. Microbiol. 2016, 18, 4974–4989. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, S.; Li, H.; Zhu, J.; Liu, Z.; Hu, X.; Yi, J. Assessments of probiotic potentials of Lactiplantibacillus plantarum strains isolated from Chinese traditional fermented food: Phenotypic and genomic analysis. Front. Microbiol. 2022, 13, 895132. [Google Scholar] [CrossRef]
- Mo, L.; Jin, H.; Pan, L.; Hou, Q.; Li, C.; Darima, I.; Zhang, H.; Yu, J. Biodiversity of lactic acid bacteria isolated from fermented milk products in Xinjiang, China. Food Biotechnol. 2019, 33, 174–192. [Google Scholar] [CrossRef]
- Jani, A.J. Amphibian microbiome linked to climate. Nat. Ecol. Evol. 2019, 3, 332–333. [Google Scholar] [CrossRef]
- Romero, F.; Cazzato, S.; Walder, F.; Vogelgsang, S.; Bender, S.F.; van der Heijden, M.G.A. Humidity and high temperature are important for predicting fungal disease outbreaks worldwide. New Phytol. 2022, 234, 1553–1556. [Google Scholar] [CrossRef]
- Zhou, W.; Li, W.; Chen, J.; Zhou, Y.; Wei, Z.; Gong, L. Microbial diversity in full-scale water supply systems through sequencing technology: A review. Rsc Adv. 2021, 11, 25484–25496. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; de Carvalho Neto, D.P.; Maske, B.L.; De Dea Lindner, J.; Vale, A.S.; Favero, G.R.; Viesser, J.; de Carvalho, J.C.; Goes-Neto, A.; Soccol, C.R. An updated review on bacterial community composition of traditional fermented milk products: What next-generation sequencing has revealed so far? Crit. Rev. Food Sci. Nutr. 2022, 62, 1870–1889. [Google Scholar] [CrossRef]
- Temmerman, R.; Huys, G.; Swings, J. Identification of lactic acid bacteria: Culture-dependent and culture-independent methods. Trends Food Sci. Technol. 2004, 15, 348–359. [Google Scholar] [CrossRef]
- Cuamatzin-Garcia, L.; Rodriguez-Rugarcia, P.; El-Kassis, E.G.; Galicia, G.; Meza-Jimenez Maria, d.L.; Banos-Lara Ma, d.R.; Zaragoza-Maldonado Diego, S.; Perez-Armendariz, B. Traditional fermented foods and beverages from around the world and their health benefits. Microorganisms 2022, 10, 1151. [Google Scholar] [CrossRef] [PubMed]
- Plessas, S. The rendering of traditional fermented foods in human diet: Distribution of health benefits and nutritional benefits. Fermentation 2022, 8, 751. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Liu, F.; Jin, Z.; Xia, X. Ecological succession and functional characteristics of lactic acid bacteria in traditional fermented foods. Crit. Rev. Food Sci. Nutr. 2022, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-N.; Han, Y.; Zhou, Z.-J. Lactic acid bacteria in traditional fermented Chinese foods. Food Res. Int. 2011, 44, 643–651. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Perez-Munoz, M.E.; Leulier, F.; Ganzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. Fems Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef]
- Farrell, J.; Rose, A. Temperature effects on microorganisms. Annu. Rev. Microbiol. 1967, 21, 101–120. [Google Scholar] [CrossRef]
- Daniel, R.M.; Peterson, M.E.; Danson, M.J.; Price, N.C.; Kelly, S.M.; Monk, C.R.; Weinberg, C.S.; Oudshoorn, M.L.; Lee, C.K. The molecular basis of the effect of temperature on enzyme activity. Biochem. J. 2010, 425, 353–360. [Google Scholar] [CrossRef]
- Hassan, N.; Anesio, A.M.; Rafiq, M.; Holtvoeth, J.; Bull, I.; Haleem, A.; Shah, A.A.; Hasan, F. Temperature driven membrane lipid adaptation in glacial psychrophilic bacteria. Front. Microbiol. 2020, 11, 824. [Google Scholar] [CrossRef]
- Bonatelli, M.L.; Lacerda-Junior, G.V.; dos Reis Junior, F.B.; Fernandes-Junior, P.I.; Melo, I.S.; Quecine, M.C. Beneficial plant-associated microorganisms from semiarid regions and seasonally dry environments: A review. Front. Microbiol. 2021, 11, 553223. [Google Scholar] [CrossRef] [PubMed]
- Inouye, M.; Phadtare, S. Cold shock response and adaptation at near-freezing temperature in microorganisms. Sci. STKE Signal Transduct. Knowl. Environ. 2004, 2004, pe26. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K. Mechanism of bacterial adaptation to low temperature. J. Biosci. 2006, 31, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-Q.; Jinadasa, K.B.S.N.; Gersberg, R.M.; Liu, Y.; Tan, S.K.; Ng, W.J. Application of constructed wetlands for wastewater treatment in tropical and subtropical regions (2000–2013). J. Environ. Sci. 2015, 30, 30–46. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhen, Y. Looking for life activity in ionic liquids. J. Mol. Liq. 2022, 349, 118185. [Google Scholar] [CrossRef]
- Frenkel-Pinter, M.; Rajaei, V.; Glass, J.B.; Hud, N.V.; Williams, L.D. Water and life: The medium is the message. J. Mol. Evol. 2021, 89, 2–11. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Z.; Hu, S.; Yu, J. On the molecular mechanism of GC content variation among eubacterial genomes. Biol. Direct 2012, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, J.; Song, Y.; Zhang, J.; Yu, Z.; Zhang, H.; Sun, Z. Comparative genomics of the herbivore gut symbiont Lactobacillus reuteri reveals genetic diversity and lifestyle adaptation. Front. Microbiol. 2018, 9, 1151. [Google Scholar] [CrossRef]
- Lo, W.-S.; Huang, Y.-Y.; Kuo, C.-H. Winding paths to simplicity: Genome evolution in facultative insect symbionts. Fems Microbiol. Rev. 2016, 40, 855–874. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.P.C.; Danchin, A. Base composition bias might result from competition for metabolic resources. Trends Genet. 2002, 18, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Beaz-Hidalgo, R.; Hossain, M.J.; Liles, M.R. Taxonomic affiliation of new genomes should be verified using average nucleotide identity and multilocus phylogenetic analysis. Genome Announc. 2014, 2, e00927-14. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zang, X.; Chen, Y.; Gao, Y.; Pei, Z.; Yang, B.; Zhang, H.; Narbad, A.; Tian, F.; Zhai, Q.; et al. Phenotype-genotype analysis of Latilactobacills curvatus from different niches: Carbohydrate metabolism, antibiotic resistance, bacteriocin, phage fragments and linkages with CRISPR-Cas systems. Food Res. Int. 2022, 160, 111640. [Google Scholar] [CrossRef]
- Wels, M.; Siezen, R.; van Hijum, S.; Kelly, W.J.; Bachmann, H. Comparative genome analysis of Lactococcus lactis indicates niche adaptation and resolves genotype/phenotype disparity. Front. Microbiol. 2019, 10, 4. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Zhang, D.; Liu, H.; Wang, S.; Wang, Y.; Ji, H. Complete genome sequencing and comparative genome characterization of Lactobacillus johnsonii ZLJ010, a potential probiotic with health-promoting properties. Front. Genet. 2019, 10, 812. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef]
- Brigandt, I. Homology in comparative, molecular, and evolutionary developmental biology: The radiation of a concept. J. Exp. Zool. Part B-Mol. Dev. Evol. 2003, 299B, 9–17. [Google Scholar] [CrossRef]
- Surve, S.; Shinde, D.B.; Kulkarni, R. Isolation, characterization and comparative genomics of potentially probiotic Lactiplantibacillus plantarum strains from Indian foods. Sci. Rep. 2022, 12, 1940. [Google Scholar] [CrossRef] [PubMed]
- Amin, K.; Tranchimand, S.; Benvegnu, T.; Abdel-Razzak, Z.; Chamieh, H. Glycoside hydrolases and glycosyltransferases from hyperthermophilic archaea: Insights on their characteristics and applications in biotechnology. Biomolecules 2021, 11, 1557. [Google Scholar] [CrossRef] [PubMed]
- Szymanska-Chargot, M.; Zdunek, A. Use of FT-IR spectra and PCA to the bulk characterization of cell wall residues of fruits and vegetables along a fraction process. Food Biophys. 2013, 8, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol.-Mysore 2012, 49, 255–266. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermented Vegetables (n = 37) | Fermented Bean Products (n = 9) | Fermented Meat Products (n = 8) | Average | |
|---|---|---|---|---|
| Content of GC (%) | 44.21 ± 0.27 a | 44.68 ± 0.34 a | 44.16 ± 0.05 a | 44.28 ± 0.20 |
| Genome size (Mb) | 3.63 ± 0.10 a | 3.63 ± 0.22 a | 3.37 ± 0.02 a | 3.59 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhu, J.; Xiao, Y.; Zhang, S.; Sun, Y.; Liu, Z.; Chu, C.; Hu, X.; Yi, J. Biodiversity of Lactic Acid Bacteria in Traditional Fermented Foods in Yunnan Province, China, and Comparative Genomics of Lactobacillus plantarum. Fermentation 2023, 9, 402. https://doi.org/10.3390/fermentation9040402
Li H, Zhu J, Xiao Y, Zhang S, Sun Y, Liu Z, Chu C, Hu X, Yi J. Biodiversity of Lactic Acid Bacteria in Traditional Fermented Foods in Yunnan Province, China, and Comparative Genomics of Lactobacillus plantarum. Fermentation. 2023; 9(4):402. https://doi.org/10.3390/fermentation9040402
Chicago/Turabian StyleLi, Hong, Jiang Zhu, Yue Xiao, Shiyao Zhang, Yuwei Sun, Zhijia Liu, Chuanqi Chu, Xiaosong Hu, and Junjie Yi. 2023. "Biodiversity of Lactic Acid Bacteria in Traditional Fermented Foods in Yunnan Province, China, and Comparative Genomics of Lactobacillus plantarum" Fermentation 9, no. 4: 402. https://doi.org/10.3390/fermentation9040402
APA StyleLi, H., Zhu, J., Xiao, Y., Zhang, S., Sun, Y., Liu, Z., Chu, C., Hu, X., & Yi, J. (2023). Biodiversity of Lactic Acid Bacteria in Traditional Fermented Foods in Yunnan Province, China, and Comparative Genomics of Lactobacillus plantarum. Fermentation, 9(4), 402. https://doi.org/10.3390/fermentation9040402