Molecular Characterization and Positive Impact of Brassinosteroids and Chitosan on Solidago canadensis cv. Tara Characteristics

 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Planting Material
2.2. System of Lighting
2.3. Experiments Treatments
2.4. Morphological Data
2.5. Flowering Data
2.6. Pigment Content
2.7. Nitrogen Determination (%)
2.8. Phosphorus Determination (%)
2.9. Potassium Determination (%)
2.10. Bioactive Compounds
2.10.1. Preparation of Solidago canadensis Plant Extracts
2.10.2. Determination of Total Phenolic Content
2.10.3. Determination of Antioxidant Activity
2.10.4. Identification of Phenolic Compounds of Solidago Extracts Using HPLC
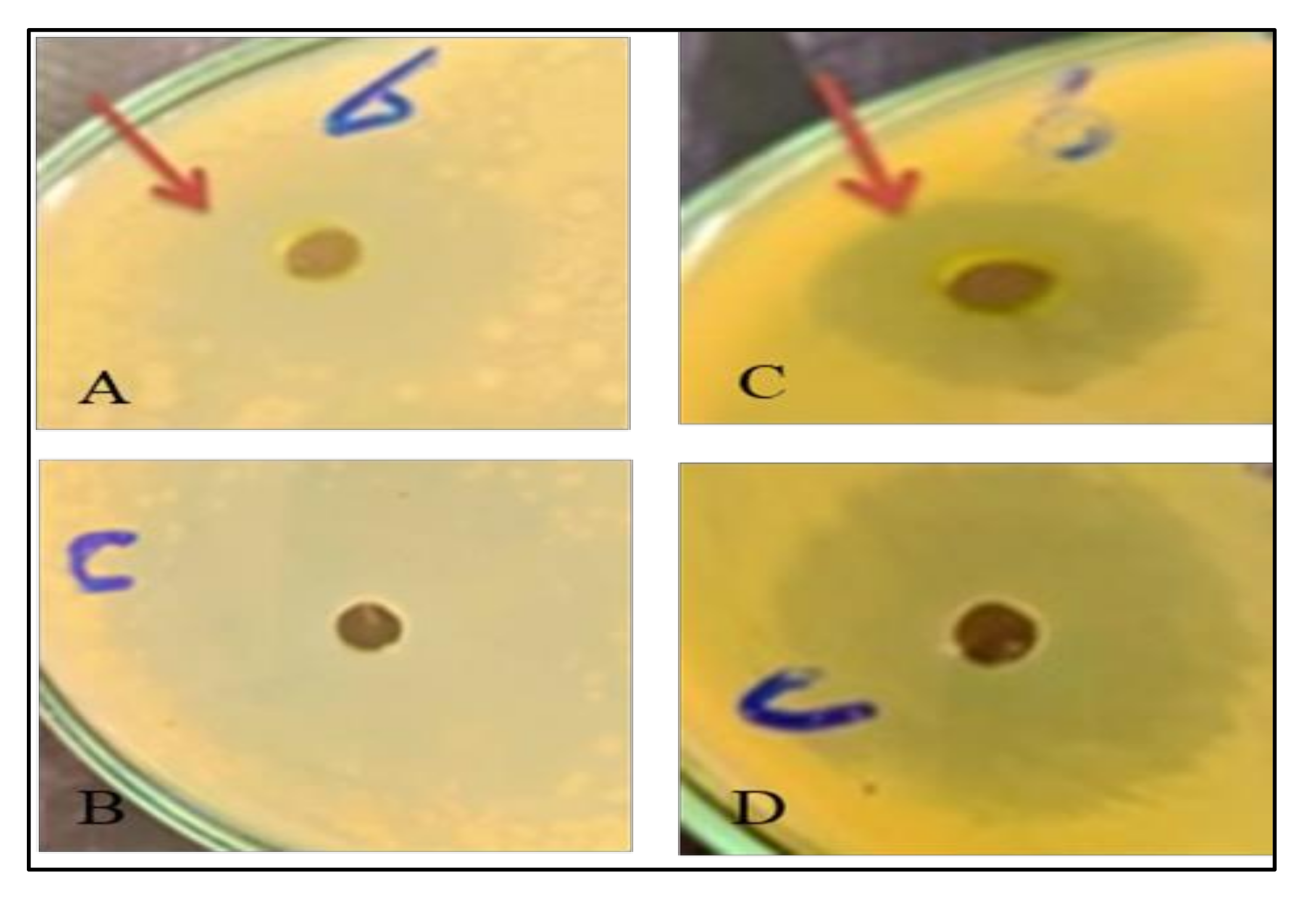
2.11. Antibacterial Activity
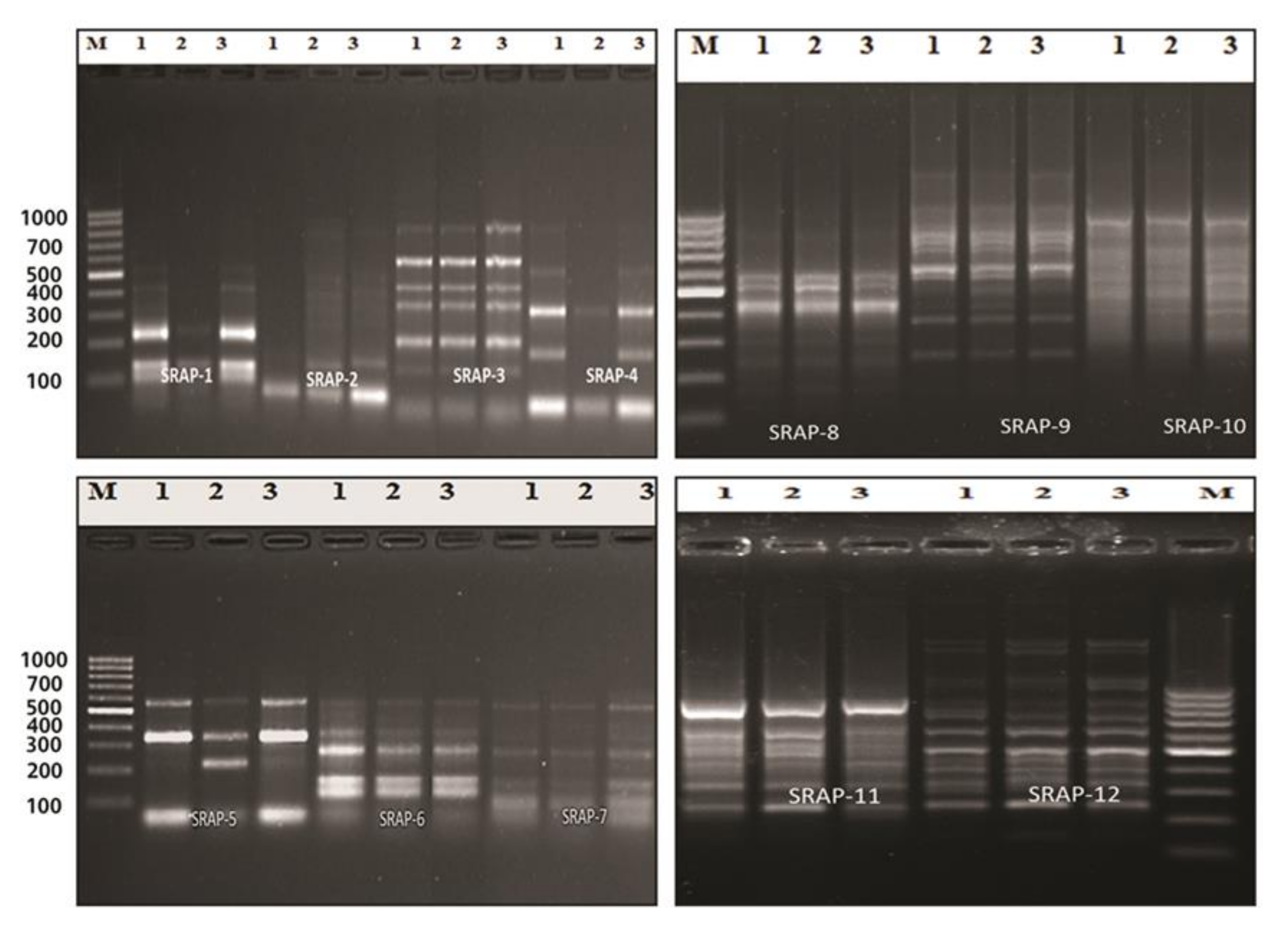
2.12. Sequence-Related Amplified Polymorphism (SRAP)
2.12.1. SRAP-PCR Reactions
2.12.2. Data Analysis
2.13. Statistical Analysis
3. Results and Discussion
3.1. Morphological Characteristics
3.2. Flowering Parameters
3.3. Vase Life
3.4. Chemical Composition
3.4.1. Pigment Content
3.4.2. Content of Macronutrients
3.4.3. Total Phenolic Content
3.4.4. Antioxidant Activity
3.4.5. Identification and Quantification of Individual Polyphenolic Compounds of Solidago canadensis L cv. Tara Extracts
3.5. Antibacterial Activities
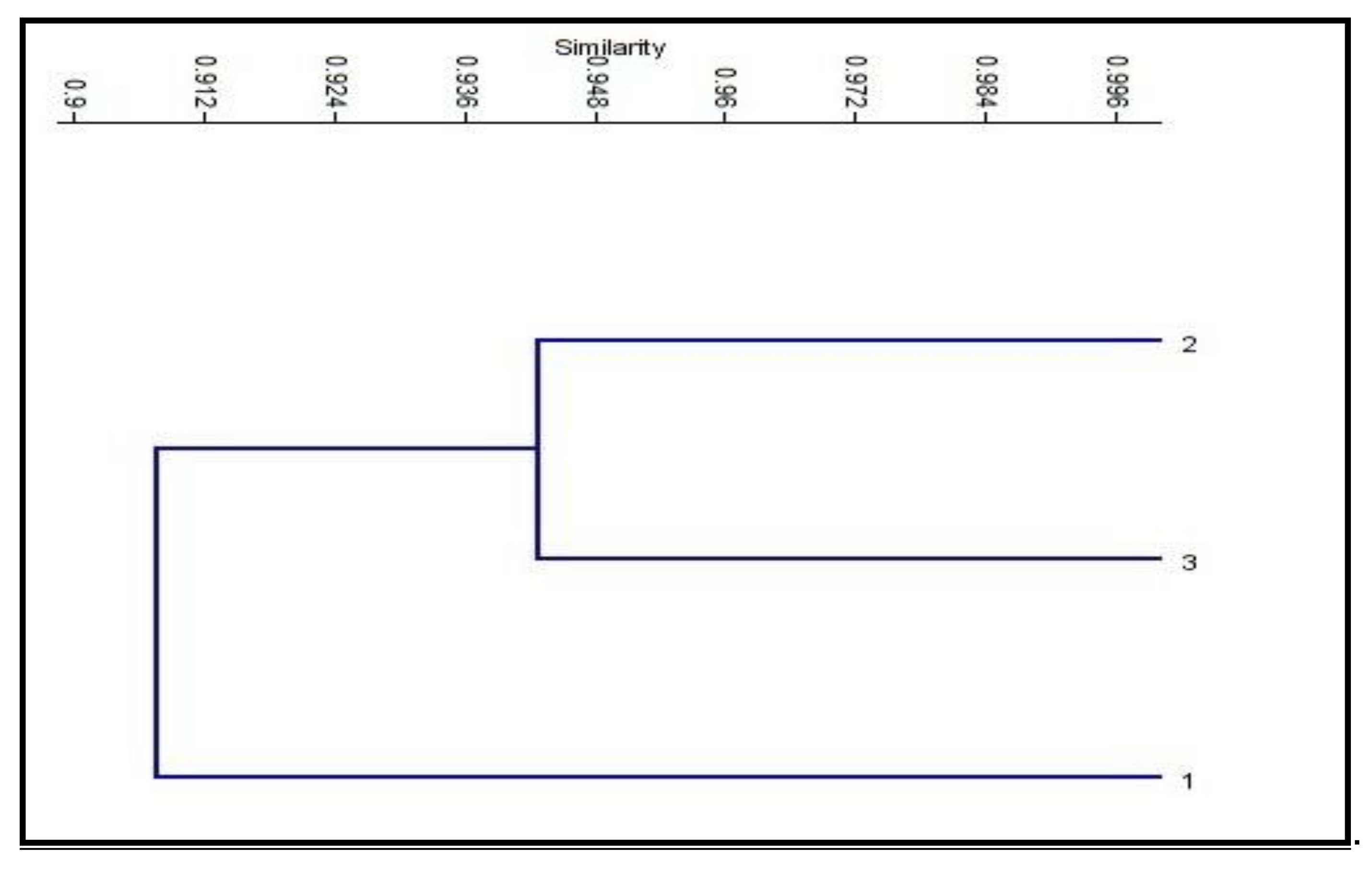
3.6. SRAP-PCR Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Walck, J.L.; Baskin, J.M.; Baskin, C.C. Why is Solidago shortii narrowly endemic and S. altissima geographically widespread? A comprehensive comparative study of biological traits. J. Biogeogr. 2002, 28, 1221–1237. [Google Scholar] [CrossRef]
- Osman, A.R.; Sewedan, E. Effect of planting density and gibberellic acid on quantitative and qualitative characteristics of Solidago canadensis “Tara” in Egypt. Asian J. Crop. Sci. 2014, 6, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Bader, G. Solidago: Constituents, pharmacology, clinical aspects and cultivation. Z. Phytother. 1999, 20, 196–400. [Google Scholar]
- Kołodziej, B. Antibacterial and antimutagenic activity of extracts aboveground parts of three Solidago species: Solidago virgaurea L., Solidago canadensis L. and Solidago gigantea Ait. J. Med. Plants Res. 2011, 5, 6770–6779. [Google Scholar] [CrossRef]
- Ravichandiran, V.; Deepa, N. In-vitro anti-cancer activity of Solidago canadensis L. Int. J. Res. Pharm. Sci. 2012, 3, 158–161. [Google Scholar]
- Bradette-Hébert, M.-E.; Legault, J.; Lavoie, S.; Pichette, A. A new labdane diterpene from the flowers of Solidago canadensis. Chem. Pharm. Bull. 2008, 56, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Khangoli, S. Potential of growth regulators on height and flowering control in ornamental plants. In Proceedings of the 1st National Scientific and Practical Symposium on Flowers and Ornamental Plants of Iran, Jahad-e-Keshavarzi, Iran, 16–18 October 2001. (In Persian). [Google Scholar]
- Fathima, S.A.; Johnson, M.; Lingakumar, K. Effect of crude brassinosteroid extract on growth and biochemical changes of Gosssypium hirsutum L. and Vigna mungo L. J. Stress Physiol. Biochem. 2011, 7, 324–334. [Google Scholar]
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146, dev151894. [Google Scholar] [CrossRef] [Green Version]
- Malerba, M.; Cerana, R. Recent advances of chitosan applications in plants. Polymers 2018, 10, 118. [Google Scholar] [CrossRef] [Green Version]
- Anonim. Fitosan. Jakarta: Laboratorium BahanIndustri-Pusat Aplikasi Teknologi Isotop & Radiasi. 2016.
- Panwar, P.; Nath, M.; Yadav, V.K.; Gupta, A.K. Comparative evaluation of genetic diversity using RAPD, SSR and cytochrome P450 gene based markers with respect to calcium content in finger millet (Eleusine coracana L. Gaertn.). J. Genet. 2010, 89, 121–133. [Google Scholar] [CrossRef]
- Ansari, S.A.; Narayanan, C.; Wali, S.A.; Kumar, R. ISSR markers for analysis of molecular diversity and genetic structure of Indian teak (Tectona grandis L.f.) populations. Ann. For. Res. 2012, 55, 11–23. [Google Scholar]
- Wang, Y.; Reighard, G.L.; Layne, D.R.; Abbott, A.G.; Huang, H. Inheritance of AFLP markers and their use for genetic diversity analysis in wild and domesticated pawpaw [Asimina triloba (L.) Dunal]. J. Am. Soc. Hortic. Sci. 2005, 130, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Quiros, C.F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica. Theor. Appl. Genet. 2001, 103, 455–461. [Google Scholar] [CrossRef]
- Highsun, E. Significant Variations in Seed, Variety and Crop Performance, in Results and in Crop Outcomes May Occur Depending upon Geographic Location, Climate, Soil Type, Soil Conditions, Cultural and Management Practices and Other Growth and Development Factors. Solidago Gold Princess 2008, 1–2. Available online: www.highsun.com.au (accessed on 7 December 2020).
- Wallace, D.H.; Munger, H.M. Studies of the physiological basis for yield differences. I. Growth analysis of six dry bean varieties 1. Crop. Sci. 1965, 5, 343–348. [Google Scholar] [CrossRef]
- Rapeseed. Determination of chlorophyll content. Spectrometric method. BSI Br. Stand. 2013, 215. [Google Scholar]
- Cottenie, A.; Kiekens, L.; Verloo, M.; Velghe, G.; Camerlynck, R. Chemical Analysis of Soils and Plants; State University: Ghent, Belgium, 1982; pp. 100–129. [Google Scholar]
- Snell, F.D.; Snell, C.T. Colorimetric Methods of Analysis, II; Von Nostrand: New York, NY, USA, 1949; p. 804. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of analysis for soils. Plants Waters 1961, 169–176. [Google Scholar]
- Lafka, T.-I.; Sinanoglou, V.; Lazos, E.S. On the extraction and antioxidant activity of phenolic compounds from winery wastes. Food Chem. 2007, 104, 1206–1214. [Google Scholar] [CrossRef]
- Matthäus, B. Antioxidant activity of extracts obtained from residues of different oilseeds. J. Agric. Food Chem. 2002, 50, 3444–3452. [Google Scholar] [CrossRef]
- Ilhan, S.; Savaroglu, F.; Çolak, F.; Iscen, C.; Erdemgil, F. Antimicrobial activity of Palustriella commutata Ochyra Extraxts. Turk. J. Biol. 2006, 30, 149–152. [Google Scholar]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy the Principles and Practice of Numerical Classification; Freeman WH: San Francisco, CA, USA, 1973; p. 573. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Soliman, D.M.; Badawy, E.-S.M.; Ahmed, A.H.H.; Habba, E.E.-A. Effect of mycorrhizal fungi (AMF), Brassinosteroids and sodium silicate on vegetative growth, flower production and Pb concentration of Zinnia (Zinnia elegans) plant under Pb stress. J. Environ. Sci. Technol. 2017, 10, 157–174. [Google Scholar] [CrossRef]
- Leubner-Metzger, G. Brassinosteroids promote seed germination. In Brassinosteroids; Hayat, S., Ahmad, A., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 2003; pp. 119–128. [Google Scholar]
- Castorina, G.; Consonni, G. The Role of Brassinosteroids in controlling plant height in Poaceae: A genetic perspective. Int. J. Mol. Sci. 2020, 21, 1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasmita, E.R.; Suryawati, A.; Irawati, E.B. Effectiveness of giving NPK FERTILIZER and Chitosan for the growth of Sunan Candlenut plant on marginal land. Ser. II-For. Wood Ind. Agric. Food Eng. 2020, 3, 49–60. [Google Scholar] [CrossRef]
- Irawati, E.B.; Sasmita, E.R.; Suryawati, A. Application of chitosan for vegetative growth of kemirisunan plant in marginal land. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Purwokerto, Indonesia, 2018; Volume 250, p. 012089. [Google Scholar]
- Salachna, P.; Zawadzińska, A. Effect of Chitosan on plant growth, flowering and corms yield of potted freesia. J. Eco. Eng. 2014, 15, 2014. [Google Scholar]
- Clouse, S.D. Brassinosteroid signal transduction: From receptor kinase activation to transcriptional networks regulating plant development. Plant Cell 2011, 23, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Mondal, M.M.A.; Malek, M.A.; Puteh, A.B.; Ismail, M.R.; Ashrafuzzaman, M.; Naher, L. Effect of foliar application of chitosan on growth and yield in okra. Aust. J. Crop Sci. 2012, 6, 918–921. [Google Scholar]
- Li, K.; Lu, X.Y.; Peng, L.S. Effects of carboxymethyl chitosan on key enzymes activities of nitrogen metabolism and grain protein contents in rice. J. Hunan. Agric. Univ. 2001, 27, 421–424. [Google Scholar]
- Pipattanawong, N.; Fujishige, N.; Yamane, K.; Ogata, R. Effects of Brassinosteroid on vegetative and reproductive growth in two day-neutral strawberries. J. Jpn. Soc. Hortic. Sci. 1996, 65, 651–654. [Google Scholar] [CrossRef]
- Rao, S.S.R.; Vardhini, B.V.; Sujatha, E.; Anuradha, S. Brassinosteroids-A new class of phytohormones. Curr. Sci. 2002, 82, 1239–1245. [Google Scholar]
- Utsunomiya, N.; Kinai, H. Effect of chitosan-oligosaccharides soil conditioner on the growth of passion fruit. J. Jpn. Soc. Hort. Sci. 1994, 64, 176–177. (In Japanese) [Google Scholar]
- Ohta, K.; Morishita, S.; Suda, K.; Kobayashi, N.; Hosoki, T. Effects of Chitosan soil mixture treatment in the seedling stage on the growth and flowering of several ornamental plants. J. Jpn. Soc. Hortic. Sci. 2004, 73, 66–68. [Google Scholar] [CrossRef]
- Mandava, N.B.; Sasse, J.M.; Yopp, J.H. Brassinolide, a growth-promoting steroidal lactone. II. Activity in selected gibberellin and cytokinin bioassays. Physiol. Plant. 1981, 53, 453–461. [Google Scholar] [CrossRef]
- Sağlam-Çağ, S. The Effect of Epibrassinolide on senescence in wheat leaves. Biotechnol. Biotechnol. Equip. 2007, 21, 63–65. [Google Scholar] [CrossRef] [Green Version]
- Solgi, M. The application of new environmentally friendly compounds on postharvest characteristics of cut carnation (Dianthus caryophyllus L.). Braz. J. Bot. 2018, 41, 515–522. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef]
- Broek, L.A.M.V.D.; Knoop, R.J.; Kappen, F.H.; Boeriu, C.G. Chitosan films and blends for packaging material. Carbohydr. Polym. 2015, 116, 237–242. [Google Scholar] [CrossRef]
- Arteca, R.N. Methodology for the Extraction, Purification, and Determination of Plant Growth Substances. In Plant Growth Substances: Principles and Applications; Springer: Boston, MA, USA, 1996; pp. 28–44. [Google Scholar]
- He, Y.J.; Xu, R.J.; Zhao, Y.J. Enhancement of senescence by epibrassinolide in leaves of mungbean seedling. Acta Physiol. Sin. 1996, 22, 58–62. [Google Scholar]
- Siddiqu, M.W. Eco-Friendly Technology for Postharvest Produce Quality, 1st ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 269–296. [Google Scholar]
- Zhang, M.; Zhai, Z.; Tian, X.; Duan, L.; Li, Z. Brassinolide alleviated the adverse effect of water deficits on photosynthesis and the antioxidant of soybean (Glycine max L.). Plant Growth Regul. 2008, 56, 257–264. [Google Scholar] [CrossRef]
- Sadak, M.S.; El-Awadi, M.E.-S.; Dawood, M.G.; El-Rorkiek, K.G.A. Physiological Role of Brassinosteroids and cauliflower extract on quinoa plant grown under sandy soil. Asian J. Appl. Sci. 2020, 13, 68–75. [Google Scholar] [CrossRef]
- Yu, J.Q.; Huang, L.F.; Hu, W.H.; Zhou, Y.H.; Mao, W.H.; Ye, S.F.; Nogués, S. A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp. Bot. 2004, 55, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, H.M.M.; Hasaneen, M.N.A.; Omer, A.M. Nano chitosan-NPK fertilizer enhances the growth and productivity of wheat plants grown in sandy soil. Span. J. Agric. Res. 2016, 14, e0902. [Google Scholar] [CrossRef]
- El-Tanahy, A.M.M.; Mahmoud, A.R.; Abde-Mouty, M.M.; Ali, A.H. Effect of chitosan doses and nitrogen sources on the growth, yield and seed quality of cowpea. Aust. J. Basic Appl. Sci. 2012, 6, 115–121. [Google Scholar]
- Raghu, K.; Rao, R. Effect of brassinosteroids on antioxidants content and radical scavenging activity of Tinospora cordifolia (Willd.) Miers ex Hook. F & Thoms. J. Med. Plants 2016, 4, 117–121. [Google Scholar]
- Bautista-Baños, S.; Hernández-Lauzardo, A.; Valle, M.V.-D.; Hernández-López, M.; Barka, E.A.; Bosquez-Molina, E.; Wilson, C. Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop. Prot. 2006, 25, 108–118. [Google Scholar] [CrossRef]
- Benhamou, N. Elicitor-induced plant defence pathways. Trends Plant Sci. 1996, 1, 233–240. [Google Scholar] [CrossRef]
- Liu, J.; Tian, S.; Meng, X.; Xu, Y. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest Biol. Technol. 2007, 44, 300–306. [Google Scholar] [CrossRef]
- Salimgandomi, S.; Shabrangi, A. The effect of Chitosan on antioxidant activity and some secondary metabolites of Mentha piperita L. J. Pharm. Health Sci. 2016, 4, 135–142. [Google Scholar]
- Wang, S.Y.; Lin, H.-S. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar] [CrossRef]
- Zilic, S.; Šukalović, V.H.-T.; Dodig, D.; Maksimović, V.; Maksimović, M.; Basić, Z. Antioxidant activity of small grain cereals caused by phenolics and lipid soluble antioxidants. J. Cereal Sci. 2011, 54, 417–424. [Google Scholar] [CrossRef]
- Ghasemnezhad, M.; Shiri, M.A. Effect of chitosan coatings on some quality indices of apricot (Prunus armeniaca L.) during cold storage. Casp. J. Environ. Sci. 2010, 8, 25–33. [Google Scholar]
- Mao, S.; Sun, W.; Kissel, T. Chitosan-based formulations for delivery of DNA and siRNA. Adv. Drug Deliv. Rev. 2010, 62, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Apáti, P.; Kéry, Á.; Houghton, P.J.; Steventon, G.B.; Kite, G. In-vitro effect of flavonoids fromSolidago canadensisextract on glutathione S-transferase. J. Pharm. Pharmacol. 2006, 58, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, M.J.; Ferreira, I.C.; Froufe, H.J.C.; Abreu, R.M.V.; Martins, A.; Pintado, M.M.E. Antimicrobial activity of phenolic compounds identified in wild mushrooms, SAR analysis and docking studies. J. Appl. Microbiol. 2013, 115, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Bajko, E.; Kalinowska, M.; Borowski, P.; Siergiejczyk, L.; Lewandowski, W. 5-O-Caffeoylquinic acid: A spectroscopic study and biological screening for antimicrobial activity. LWT Food Sci. Technol. 2016, 65, 471–479. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Abdollahi, M.; Rahimi, R. Role of dietary polyphenols in the management of peptic ulcer. World J. Gastroenterol. 2015, 21, 6499–6517. [Google Scholar] [CrossRef]
- Liu, S.; Shao, X.; Wei, Y.; Li, Y.; Xu, F.; Wang, H. Solidago canadensis L. Essential oil vapor effectively inhibits botrytis cinerea growth and preserves postharvest quality of strawberry as a food model system. Front. Microbiol. 2016, 7, 1179. [Google Scholar] [CrossRef]
- Hayat, S.H.A.M.S.U.L.; Mori, M.; Fariduddin, Q.A.Z.I.; Bajguz, A.N.D.R.Z.E.J.; Ahmad, A. Physiological role of brassinosteroids: An update. Indian J. Plant Physiol. 2010, 15, 99–109. [Google Scholar]
- Yu, M.; Ma, B.; Luo, X.; Zheng, L.; Xu, X.; Yang, Z. Molecular diversity of Auricularia polytricha revealed by inter-simple sequence repeat and sequence-related amplified polymorphism markers. Curr. Microbiol. 2008, 56, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, S.; Zhao, H.; Wang, H. Genetic linkage map of EST-SSR and SRAP markers in the endangered Chinese endemic herb Dendrobium (Orchidaceae). Genet. Mol. Res. 2012, 11, 4654–4667. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, S.S.; Al-Faifi, S.A.; Migdadi, H.M.; Khan, M.A.; El-Harty, E.H.; Ammar, M.H. Molecular diversity assessment using sequence related amplified polymorphism (SRAP) markers in Vicia faba L. Int. J. Mol. Sci. 2012, 13, 16457–16471. [Google Scholar] [CrossRef]
- Que, Y.-X.; Xu, L.; Lin, J.; Luo, J.; Xu, J.; Zheng, J.; Chen, R. cDNA-SRAP and its application in differential gene expression analysis: A case study in Erianthus arundinaceum. J. Biomed. Biotechnol. 2012, 2012, 390107. [Google Scholar] [CrossRef] [Green Version]
- El-Nashar, Y.; Ammar, M. Mutagenic influences of colchicine on phenological and molecular diversity of Calendula officinalis L. Genet. Mol. Res. 2016, 15, 15027745. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chen, J.; Wang, J.; Wang, X.; Lu, Y.; Ning, Y.; Li, Y. Intra-population genetic diversity of Buchloe dactyloides (Nutt.) Engelm (buffalograss) determined using morphological traits and sequence-related amplified polymorphism markers. 3 Biotech 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zagorcheva, T.; Stanev, S.; Rusanov, K.; Atanassov, I. SRAP markers for genetic diversity assessment of lavender (Lavandula angustifolia mill.) varieties and breeding lines. Biotechnol. Biotechnol. Equip. 2019, 34, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Malerba, M.; Cerana, R. Chitosan effects on plant systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | EC dS/m * | Soluble Cations Meq/L | Soluble Anions (Meq/L) * | Soil Texture | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ca | Mg | Na | K | CO3 HCO3 | Cl | SO4 | |||
| 7.88 | 0.492 | 1 | 0.6 | 3.1 | 0.2 | - | 3 | 1.9 | Sandy |
| loam | |||||||||
| Primer | Forward Primer | Reverse Primer |
|---|---|---|
| SRAP-1 | ME1-5′-TGAGTCCAAACCGGATA-3′ | EM1-5′-GACTGCGTACGAATTAAT-3 |
| SRAP-2 | ME1-5′-TGAGTCCAAACCGGATA-3′ | EM2-5′-GACTGCGTACGAATTTGC-3′ |
| SRAP-3 | ME1-5′-TGAGTCCAAACCGGATA-3′ | EM3-5′-GACTGCGTACGAATTGAC-3′ |
| SRAP-4 | ME1-5′-TGAGTCCAAACCGGATA-3′ | EM4-5′-GACTGCGTACGAATTTGA-3′ |
| SRAP-5 | ME2-5′-TGAGTCCAAACCGGAGC-3′ | EM1-5′-GACTGCGTACGAATTAAT-3 |
| SRAP-6 | ME2-5′-TGAGTCCAAACCGGAGC-3′ | EM2-5′-GACTGCGTACGAATTTGC-3′ |
| SRAP-7 | ME2-5′-TGAGTCCAAACCGGAGC-3′ | EM3-5′-GACTGCGTACGAATTGAC-3′ |
| SRAP-8 | ME3-5′-TGAGTCCAAACCGGAAT-3′ | EM1-5′-GACTGCGTACGAATTAAT-3 |
| SRAP-9 | ME3-5′-TGAGTCCAAACCGGAAT-3′ | EM2-5′-GACTGCGTACGAATTTGC-3′ |
| SRAP-10 | ME3-5′-TGAGTCCAAACCGGAAT-3′ | EM3-5′-GACTGCGTACGAATTGAC-3′ |
| SRAP-11 | ME4-5′-TGAGTCCAAACCGGACC-3′ | EM1-5′-GACTGCGTACGAATTAAT-3 |
| SRAP-12 | ME4-5′-TGAGTCCAAACCGGACC-3′ | EM2-5′-GACTGCGTACGAATTTGC-3′ |
| Characters | First Season | ||||||
|---|---|---|---|---|---|---|---|
| Treatments | Plant Height (cm)/Plant | No. of Leaves/Shoot | Stem Diameter (cm)/Plant | Total Leaf Area/Plant (cm2) | Herb F.W (g) | Herb D.W (g) | |
| Control | 53.50 e | 46.00 c | 0.40 d | 271.66 f | 4.25 f | 1.88 d | |
| BRs 10−3 M | 68.67 b,c | 52.00 b,c | 0.50 b,c | 444.20 d | 6.16 e | 2.96 c | |
| BRs 10−6 M | 70.39 b | 55.00 a,b | 0.56 a,b | 505.76 b | 10.33 b,c | 4.86 a,b | |
| BRs 10−8 M | 76.16 a | 58.00 a | 0.60 a | 690.43 a | 11.30 a | 5.56 a | |
| CHT100 mg/L | 60.15 d | 48.00 c | 0.42 d | 339.15 e | 8.62 d | 4.02 b,c | |
| CHT 150 mg/L | 64.50 c | 51.00 b,c | 0.45 c,d | 444.48 d | 9.78 c | 4.59 a,b | |
| CHT 200 mg/L | 72.07 a,b | 55.00 a,b | 0.57 a | 523.35 c | 11.02 a,b | 4.92 a,b | |
| Character | Second Season | ||||||
| Treatments | Plant Height (cm)/Plant | No. of Leaves/Shoot | Stem Diameter (cm)/Plant | Total Leaf Area/Plant (cm2) | Herb F.W (g) | Herb D.W (g) | |
| Control | 48.01 e | 43.00 b | 0.39 c | 152.83 g | 3.37 e | 1.70 d | |
| BRs 10−3 M | 65.05 b,c | 50.00 b | 0.50 b | 352.13 d | 5.18 d | 2.70 c,d | |
| BRs 10−6 M | 66.48 a,b | 53.00 a,b | 0.54 a,b | 503.59 b | 9.37 a,b | 4.67 a,b | |
| BRs 10−8 M | 72.03 a | 64.00 a | 0.58 a | 586.58 a | 10.89 a | 5.41 a | |
| CHT100 mg/L | 59.15 c | 44.00 b | 0.40 c | 207.06 f | 6.88 c | 3.60 b,c | |
| CHT 150 mg/L | 55.77 d | 46.00 b | 0.40 c | 274.29 e | 8.62 b | 4.35 a,b | |
| CHT 200 mg/L | 69.03 a,b | 53.00 a,b | 0.54 a,b | 381.2 c | 10.01 a,b | 5.01 a | |
| Characters | First Season | ||||||
|---|---|---|---|---|---|---|---|
| Treatments | No of Inflorescence /Plant | Length of Inflorescence Stalks (cm) | Days to Flower (Days) | F.W. Flowers /Plant (g) | D.W. Flowers /Plant (g) | Vase Life/Day | |
| Control | 19.00 d | 65.60 g | 115.72 a | 4.25 f | 1.88 d | 8.66 e | |
| BRs 10−3 M | 20.00 c,d | 74.50 f | 115.52 a | 6.16 e | 2.96 c | 10.03 d,e | |
| BRs 10−6 M | 28.00 b | 97.02 c | 110.25 c | 10.33 b,c | 4.86 a,b | 11.22 d | |
| BRs 10−8 M | 35.00 a | 112.02 a | 109.24 c,d | 11.30 a | 5.56 a | 13.03 c | |
| CHT100 mg/L | 23.00 c | 86.66 e | 113.12 b | 8.62 d | 4.02 b,c | 14.22 c | |
| CHT 150 mg/L | 27.00 b | 93.06 d | 111.22 b,c | 9.78 c | 4.59 a,b | 16.88 b | |
| CHT 200 mg/L | 29.00 b | 106.51 b | 107.18 d | 11.02 a,b | 4.92 a,b | 20.11 a | |
| Character | Second Season | ||||||
| Treatments | No of Inflorescence/Plant | Length of Inflorescence Stalks (cm) | Days to Flower (Days) | F.W Flowers /Plant (g) | D.W Flowers /Plant (g) | Vase Life/Day | |
| Control | 16.00 d | 55.04 g | 112.34 a | 3.37 e | 1.70 d | 7.88 d | |
| BRs 10−3 M | 18.00 c,d | 63.35 f | 110.25 b | 5.18 d | 2.70 c,d | 8.91 d | |
| BRs 10−6 M | 26.00 b | 86.05 c | 106.39 c,d | 9.37 a,b | 4.67 a,b | 10.14 c,d | |
| BRs 10−8 M | 32.00 a | 105.09 a | 105.29 d | 10.89 a | 5.41 a | 12.71 b,c | |
| CHT100 mg/L | 20.00 c | 78.20 d | 112.25 a | 6.88 c | 3.60 b,c | 13.34 b,c | |
| CHT 150 mg/L | 25.00 b | 75.94 e | 108.12 c | 8.62 b | 4.35 a,b | 15.54 b | |
| CHT 200 mg/L | 27.00 b | 93.55 b | 104.44 d | 10.01 a,b | 5.01 a | 19.02 a | |
| Treatments | Chemical Composition | |||||||
|---|---|---|---|---|---|---|---|---|
| Chlorophyll a mg/g F.W. | Chlorophyll b mg/g F.W. | Carotenoids mg/g F.W. | N % | P% | K% | Total Phenolic (mg GAE/g Tissue) | Antioxidant Activity | |
| Control | 0.65 c | 0.23 e | 0.30 e | 2.00 g | 0.20 c | 1.96 d | 45.76 e | 14.40 e |
| BRs 10−3 M | 0.82 b,c | 0.32 b | 0.46 c | 2.67 d | 0.22 b,c | 2.54 b | 63.11 c | 19.32 c |
| BRs 10−6 M | 0.92 b | 0.34 b | 0.52 b | 2.88 c | 0.24 b | 2.61 b | 66.68 b | 27.30 b |
| BRs 10−8 M | 1.35 a | 0.45 a | 0.68 a | 3.29 b | 0.27 a | 2.77 a | 71.47 a | 29.82 b |
| CHT100 mg/L | 0.70 c | 0.30 b | 0.31 e | 2.09 f | 0.20 c | 2.01 d | 55.77 d | 16.11 d,e |
| CHT 150 mg/L | 0.74 c | 0.30 b | 0.41 d | 2.29 e | 0.21 c | 2.15 c | 61.01 c | 18.79 c,d |
| CHT 200 mg/L | 1.37 a | 0.47 a | 0.70 a | 3.44 a | 0.29 a | 2.88 a | 72.92 a | 38.72 a |
| Polyphenolic Compound | Polyphenolic Compounds Content of Solidago canadensis cv Tara Extracts (μg/g) | ||
|---|---|---|---|
| Control * | BRs * 10−8 M | CHT *200 mg/L | |
| Gallic acid | 5.50 | 134.92 | 178.13 |
| Chlorogenic acid | 24.58 | 207.26 | 276.20 |
| Catechin | ND * | ND * | ND * |
| Methyl gallate | 4.54 | 15.27 | 31.27 |
| Caffeic acid | ND * | 9.90 | 15.57 |
| Syringic acid | 4.21 | 34.84 | 50.25 |
| Pyro catechol | 8.36 | 32.26 | 73.46 |
| Rutin | 24.71 | 170.05 | 364.10 |
| Ellagic acid | ND * | ND * | 4.82 |
| Coumaric acid | ND * | 5.23 | 5.30 |
| Vanillin | 3.20 | 27.25 | 46.52 |
| Ferulic acid | ND * | ND * | ND * |
| Naringenin | 8.25 | 67.52 | 133.42 |
| Taxifolin | ND * | 23.00 | 32.03 |
| Cinnamic acid | 1.16 | 9.21 | 12.47 |
| Kaempferol | 19.92 | 87.08 | 211.36 |
| Combination Primer | Total Bands | Polymorphic Bands | Unique Band | Bands of MW * | Polymorphism% | PIC * Value |
|---|---|---|---|---|---|---|
| ME1*EM1 | 7 | 4 | 3 | 70–520 | 57 | 0.7 |
| ME1*EM2 | 8 | 7 | 1 | 80–800 | 88 | 0.7 |
| ME1*EM3 | 7 | 0 | 0 | 70–800 | 0 | 0.1 |
| ME1*EM4 | 5 | 2 | 0 | 70–800 | 40 | 0.9 |
| ME2*EM1 | 5 | 1 | 1 | 80–600 | 10 | 0.9 |
| ME2*EM2 | 7 | 0 | 0 | 90–600 | 0 | 0.1 |
| ME2*EM3 | 6 | 1 | 0 | 100–600 | 17 | 0.9 |
| ME3*EM1 | 10 | 0 | 0 | 160–1000 | 0 | 1.0 |
| ME3*EM2 | 12 | 0 | 0 | 150–1500 | 0 | 1.0 |
| ME3*EM3 | 13 | 1 | 1 | 250–1000 | 8 | 0.9 |
| ME4*EM1 | 10 | 1 | 2 | 200–900 | 30 | 0.9 |
| ME4*EM2 | 15 | 2 | 1 | 150–1600 | 20 | 0.9 |
| Total | 115 | 19 | 9 | - | ||
| Average | 9.6 | 1.6 | - | - | 23 | 0.8 |
| Control | Control | BRs10−8 M | CHT 200 mg/L |
|---|---|---|---|
| 100 | |||
| BRs at 10−8 M | 89 | 100 | |
| CHT 200 mg/L | 93 | 94 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Sayed, I.M.; Salim, R.G.; El-Haggar, E.F.; El-Ziat, R.A.; Soliman, D.M. Molecular Characterization and Positive Impact of Brassinosteroids and Chitosan on Solidago canadensis cv. Tara Characteristics. Horticulturae 2020, 6, 100. https://doi.org/10.3390/horticulturae6040100
El-Sayed IM, Salim RG, El-Haggar EF, El-Ziat RA, Soliman DM. Molecular Characterization and Positive Impact of Brassinosteroids and Chitosan on Solidago canadensis cv. Tara Characteristics. Horticulturae. 2020; 6(4):100. https://doi.org/10.3390/horticulturae6040100
Chicago/Turabian StyleEl-Sayed, Iman M., Rasha G. Salim, Eman F. El-Haggar, Rasha A. El-Ziat, and Dina M. Soliman. 2020. "Molecular Characterization and Positive Impact of Brassinosteroids and Chitosan on Solidago canadensis cv. Tara Characteristics" Horticulturae 6, no. 4: 100. https://doi.org/10.3390/horticulturae6040100
APA StyleEl-Sayed, I. M., Salim, R. G., El-Haggar, E. F., El-Ziat, R. A., & Soliman, D. M. (2020). Molecular Characterization and Positive Impact of Brassinosteroids and Chitosan on Solidago canadensis cv. Tara Characteristics. Horticulturae, 6(4), 100. https://doi.org/10.3390/horticulturae6040100