Abstract
Cd stress can cause severe damage to the chloroplast structure and plant function. The chloroplast stay-green gene (SGR2) is senescence-inducible and has several important functions such as regulating senescence, responding to abiotic stress, and regulating seed and fruit ripening. The SlSGR2 gene was cloned from the tomato “Hongluocheng” (full-length of 819 bp encoding 272 amino acids). Subcellular localization prediction and laser confocal observation revealed that the SlSGR2 protein was localized to the chloroplast of A. thaliana protoplasts. A qRT-PCR analysis suggested that the SlSGR2 gene was upregulated by different Cd concentrations and that the gene expression was significantly upregulated in tomato roots and leaves under 50 μM CdCl2 treatment. The pCAMBIA2300-SlSGR2 overexpression constructs were introduced into wild-type Arabidopsis plants via Agrobacterium-mediated transformation. Homozygous mutant lines of sgr2 were identified using the three-primer method. The growth of SlSGR2-overexpressed lines under Cd stress was significantly better than that of WT and sgr2. The chlorophyll, SOD, POD, and CAT contents of OE4-1 and OE4-2 were significantly higher than those of WT and sgr2 while the MDA content was significantly lower than other lines under cadmium stress. This study showed that the SlSGR2 gene is involved in Cd-induced plant senescence and is a negative regulator of plant chlorophyll degradation.
1. Introduction
Cadmium (Cd) is a highly toxic heavy metal that exists widely in the environment and causes abnormalities and mutations in humans, animals, and plants [1,2]. The serious consequences of Cd contamination have been well known since the outbreak of “Itai-Itai” disease in Japan in the 1960s [3,4]. The development of heavy industry will inevitably lead to a large amount of cadmium entering the soil, water, and atmospheric environment, of which soil intake is a potentially important source of exposure to environmental pollutants for children and adults [5]. According to the first national soil pollution survey in China from 2005 to 2013, the proportion of Cd in soil samples (7.0%) exceeded the limit established by the Ministry of Environmental Protection [6]. Nearly one-fifth (about 20 million hectares) of the arable land in China was reported to be contaminated with Cd, arsenic (As), and/or lead (Pb), resulting in about 12 million tons of grains being polluted and causing about RMB 200 billion in economic losses [7]. Cd, a nonessential trace metal element that has toxic effects on plant growth and human health, is classified as the sixth most poisonous substance by the Agency for Toxic Substances and Disease Registry (ATSDR). Cd-contaminated food threatens human health all over the world and the harm to the human body is chronic due to its long half-life [8,9]. Long-term exposure to Cd and its compounds impairs calcium accumulation and metabolism, leading to fractures, proteinuria, emphysema, osteomalacia, bronchiolitis, and cancer [10,11,12]. Dietary cadmium intake has exceeded the tolerable limit recommended by the FAO/WHO (25 mg/kg BW/month) in China and some other Asian countries [13,14,15]. Humans are mainly exposed to Cd through food; the main dietary sources are grains and vegetables [16,17]. Vegetables account for 10–40% of total dietary Cd intake depending on dietary habits and the Cd concentration in food [18]. A study found that the concentration of Cd in agricultural soils in China gradually increased from 1981 to 2016 and that the time trend could be divided into two stages: an upward trend from 1981 to 2001 and a slow upward trend from 2002 to 2016 [19]. Reports showed that cancer emerged in villages in China in the 1980s, grew rapidly in the 1990s, and continued to grow after 2000, which was consistent with the temporal trend of changes in Cd concentrations in agricultural soils in China.
Cd is mainly absorbed by and accumulated in plants through soil and can be transferred to other organisms through the food chain. Agricultural products grown in Cd-contaminated soil will be contaminated [20]. In terrestrial plants, the root is the most important organ for absorbing metals including Cd and various nonmetal nutrients, so the first organ affected by Cd poisoning is the root. Cd affects plant metabolism by interfering with the absorption, transportation, and utilization of various elements (Ca, Mg, P, and K), thereby altering plant growth and development [21]. Plants exhibit reduced root length or abnormal development due to direct inhibition of root metabolism by cadmium stress [22]. The most common features of Cd toxicity to plants also affect growth [23], including leaf curling, brown spots [24,25], chlorosis, necrosis, loss of turgor pressure, stomata closure, reduced seed germination rates, and organelle damage associated with senescence processes or plant death [26,27,28]. High concentrations of cadmium reduce photosynthesis due to its direct effects on the photosynthetic system, including thylakoids. Cadmium has effects on proteins, lipids, and the release of element components of thylakoid membranes, causing damage to photosynthesis and photosystem II (PS II) [29,30]. Cadmium damage to plant photosynthetic organelles leads to decreased chlorophyll and carotenoid levels [31]. There are two damage mechanisms for the destruction of photosynthesis by Cd. The first is that Cd2+ replaces the Mg2+ in the center of chlorophyll; [Cd2+]—Chl is very unstable so it easily fades [32]. Another mechanism of Cd-induced PSII reaction center inhibition is the substitution of Cd2+ for Ca2+ in the hydrolysis complex [33,34].
All terrestrial plants synthesize chlorophyll a and chlorophyll b. Chlorophyll (Chl) is the most abundant pigment on earth and is a key component for plants to absorb the light required for photosynthesis. Studies in Arabidopsis and rice have identified six plastid-localized chlorophyll-degrading metabolizing enzymes (CCEs): Non-yellowing coloring1 (NYC1) [35,36]; Non-yellowing coloring1-like (NOL) [37]; 7-hydroxymethyl chlorophyll a reductase (HCAR), which encodes two different Chl b reductases [38]; Pheophytin pheophorbide hydrolase (PPH) [39,40]; pheophorbide a oxygenase (PAO) [41,42]; and Red chlorophyll catabolite reductase (RCCR) [41]. In addition to these proteins, a protein called STAYGREEN (SGR), which plays an important role in recruiting CCE for LHCII to aid in the breakdown of chlorophyll, is also required [43]. The gene that controls the Mendelian green cotyledon trait was identified in [44] and was named STAYGREEN1 (SGR) or NONYELLOWING1 (NYE1) in Arabidopsis [45]. Different teams at the same time independently identified SGR1 homologs in Arabidopsis [46] and rice [47]; subsequent work isolated and characterized SGR1 homologs in various plant species including tomato (Solanum lycopersicum L.) [48], sweet pepper (Capsicum annuum L.) [48], tall fescue (Festuca arundinacea Schreb.) [49], alfalfa (Medicago truncatula Gaertn.) [50], and soybean (Glycine max L.) [51]. The physiological and molecular roles and functions of SGR homologues in different plants have been revealed successively since 2010. Arabidopsis SGR2-overexpressed lines display a stay-green phenotype and sgr2 mutants exhibit a leaf-yellowing phenotype under dark-induced and abiotic stress conditions [52]. However, AtSGR1-overexpressed plants showed leaf yellowing at an early stage and the sgr1-1/nye1-1 mutant exhibited a stay-green phenotype, which was opposite to the phenotypes of the SGR2-overexpressed and sgr2 mutants. Compared with the interaction between SGR1 and CCEs, the interaction between SGR2 and CCEs is much weaker. Leaf and seed embryos of the d1 d2 double mutant in soybean showed a stay-green phenotype [53,54]. Follow-up research found that D1 and D2 encode GmSGR1 and GmSGR2, respectively [51]. Unlike the AtSGR genes, d1 leaves showed a much weaker stay-green phenotype than d1-d2 leaves while d1 seeds turned yellow normally, suggesting that the two GmSGRs in soybean are functionally redundant. Except for senescence, residual Chl retained in mature seeds has severe negative effects on seed storability and longevity, so rapid degradation of Chl is required for seed maturation [55,56]. Studies have shown that Arabidopsis SGR1 and SGR2 play key roles in seed chlorosis and are functionally redundant [57]. During seed development, SGR1 and SGR2 also play important roles in balancing Chl degradation and biosynthesis of the main component tocopherol during Arabidopsis seed maturation [58]. The tomato SlSGR1 gene balances the synthesis of two major pigments in fruit ripening—carotenoids and chlorophyll [59]. Chl degradation is induced in response to several abiotic stresses [60]. Arabidopsis SGR1 and SGRL gene-overexpressed plants exhibited an early leaf-yellowing phenotype while sgr1/nye1-1 and sgrl mutants exhibited a stay-green phenotype under abiotic stress conditions, including salt stress, osmotic stress, and drought stress [52,60,61]. The SGR-silenced lines of perennial ryegrass lost the stay-green phenotype under heat stress and leaf senescence accelerated with increased chloroplast disruption [62]. Therefore, SGR1, SGRL, and CCEs also play a role in abiotic stress-induced leaf yellowing, but the mode of action of the SGR2 gene under heavy metal stress is rarely reported.
Tomato (Solanum lycopersicum L.) belongs to Solanaceae, which is an important fruit crop worldwide with increasing production in China, Japan, and Southeast Asia [63]. Tomato is a day-neutral plant; its fruit contains B vitamins and vitamin C and is rich in carotene, which has a high nutritional value. With the advancement of social industrialization, heavy metal Cd pollution has become one of the important factors of abiotic stress in crop growth. Accumulation and poisoning of crops will eventually lead to the occurrence of human diseases. The SGR2 gene not only participates in regulating the synthesis and degradation of plant chlorophyll, but also responds to the degradation of chlorophyll under stress. The purpose of this paper was to investigate the response of the tomato SlSGR2 gene in the process of chlorophyll degradation induced by cadmium stress.
2. Materials and Methods
2.1. Materials, Growth Conditions, and Stress Treatment
Tomato (Solanum lycopersicum L.) “Hongluocheng”, Arabidopsis thanliana ecotype Columbia (WT), and transgenic A. thanliana were grown in pots within a soil/vermiculite mixture. The plants were grown under a 16 h:8 h photoperiod at temperature of 23 °C during the light and the dark periods. For assays of cadmium stress, 8-week-old seedlings were transferred into Hoagland’s liquid with 0, 25 µM, 50 µM, or 100 µM of CdCl2. The growth conditions were the same as those of potted plants; the leaves of seedlings were collected on the 3rd, 5th, and 7th day after treatment. For the gene expression analysis, the materials were processed as above and collected at 0, 3, 6, 9, and 12 h. After that, the leaves and roots of tomato were snap-frozen in liquid nitrogen and then stored at −80 ℃. The WT, SlSGR2-OE, and sgr2 mutant were grown in a soil/vermiculite mixture and watered with or without 1 mM of CdCl2 at six weeks. After 14 days of CdCl2 treatment, fresh leaves were tested for physiological indicators and other leaves of the three lines were frozen in liquid nitrogen and stored at −80 °C for the gene expression analysis. The experimental treatments were repeated three times.
2.2. Ultrastructural Analysis
Samples of the 3rd and 4th leaves of tomato that were treated with CdCl2 after five days were cut to a 1 × 3 mm size near the main vein and fixed in 2.5% glutaraldehyde buffered with deionized water and post-fixed in 1% osmium tetroxide after rinsing. The samples were embedded in epoxy resin after gradient dehydration. Ultrathin sections (Leica EM UC6, Wetzlar, Germany; stained with uranyl acetate and lead for 10 min) were analyzed using a transmission electron microscope (HITACHI H-7650, Tokyo, Japan).
2.3. Subcellular Localization
According to Phytozome 12.0, the full-length primers (GFP-SlSGR2-QF: 5′-CGGGATCCATGGGAACTTTGACTACTTCTCTAGTG-3′ GFP-SlSGR2-QR: 5′-GCTCTAGATCAACTTTGCTGCTCTTGCAAG-3′) of the CDS region of senescence-inducible chloroplast stay-green protein 2 (Solyc08g080090.2, SlSGR2) were designed to amplify the gene sequence. The coding sequence of SlSGR2 without a stop codon was inserted into the p16318hGFP. The control vector and recombinant vector (p16318hGFP-SlSGR2) were transformed into prepared A. thaliana protoplasts and the location of the SlSGR2 protein was observed through a laser confocal microscope (Lecia SP5, Wetzlar, Germany).
2.4. Total RNA Extraction, cDNA Synthesis, and Isolation of SlSGR2 Gene
The total RNA of tomato and A. thanliana were extracted by using the total RNA extraction kit (TIANGEN, Beijing, China) according to the manufacturer’s instructions. The cDNA was obtained with a First Stand cDNA Synthesis Kit (Toyobo, Japan). The full length of the CDS region of the SlSGR2 gene (Solyc08g080090.2) was amplified using RT-PCR with specific primers (SlSGR2-QM1-F: 5′-GGGGTACCATGGGAACTTTGACTACTTCTCTAGTG-3′ and SlSGR2-QM2-R: 5′-GCTCTAGATCAACTTTGCTGCTCTTGCAAG-3′).
2.5. Bioinformatics Analysis
The open reading frame (ORF) was analyzed using NCBI-ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder, accessed on 20 March 2019) according to the full-length gene sequence of the CDS region of SlSGR2. The sequences of SGR proteins from other species were downloaded from the National Center for Biotechnology Information (NCBI) database (National Center for Biotechnology Information (nih.gov)). Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA, USA) was used to design the primers. The alignment of amino acid sequences of SGRs from tomato and other species was analyzed using DNAMAN 6.0 (Lynnon BioSoft, San Ramon, CA, USA) and Clustalx 1.81 (The Conway Institute UCD Dublin, Dublin, Ireland) software. The phylogenetic tree was created with MEGA 7.0. The basic physicochemical properties of SlSGR2 protein were predicted with ExPASy—ProtParam tool (https://web.expasy.org/protparam/, accessed on 20 March 2019). The secondary structure of SlSGR2 was predicted using SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, accessed on 20 March 2019) and SWISS-MODEL (https://swissmodel.expasy.org/interactive, accessed on 20 March 2019) for tertiary structure establishment. To explore the possible function of the SlSGR2 gene, the promoter region of about 2000 bp from the upstream of start codon in the coding region was obtained through the NCBI website. The plant promoter cis-acting element analysis database P1antCARE (http://bioinfomiatics.psb.ugent.be/webtoQls/pIantcare/html, accessed on 20 March 2019) was used to visualized the promoter region. Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2, accessed on 20 March 2019) was used to predict SlSGR2’s protein subcellular localization.
2.6. Expression Pattern Analysis of SlSGR2 Gene under Cadmium Treatment
Total RNA was extracted from leaves and roots of tomato. Samples were analyzed for gene expression levels using real-time quantitative PCR (qRT-PCR) on an Applied Biosystems 7500 Fast Real-Time PCR system (ABI 7500). Primer 5.0 was used to design the primers (QRT-SlSGR2-F: 5′-CTAAACTTACTTTGGCTATCTCCC-3′ and QRT-SlSGR2-R: 5′-TTTCTGAGTCTAGCAAATAAGTCTAAC-3′). β-actin was selected as the internal reference gene. PCR products were quantified using Ultra SYBR Mixture and ROX 1 (CWBIO, Beijing, China). Each group had three biological replicates and three technological replicates.
2.7. Overexpression Vector Construction and A. thanliana Transformation
The PCR product was cloned in pCAMBIA2300 in the restriction sites of KpnⅠ and XbaⅠ and the transformation was carried out using DH5α Competent cells (Biomed, Beijing, China) and sequenced (Songon Biotech, Shanghai, China). The recombinant plasmid (35 s: SlSGR2) was introduced into Agrobacterium tumefaciens GV3101 (Biomed, Beijing, China) via heat shock conversion and then transformed into Arabidopsis thanliana using the floral dip method. Transgenic Arabidopsis seeds were collected and planted in MS medium (pH = 5.8) containing antibiotics (50 mg/L kanamycin and 200 mg/L timentin). We selected the positive, continued planting, and identified T3 generation via PCR.
2.8. Mutant Identification
The mutant was purchased from The Arabidopsis Information Resource (TAIR) home page (https://www.arabidopsis.org, accessed on 20 March 2019). Primers (LP: CAGTGACCGTAACACACCATG; LBb1.3 (BP): ATTTTGCCGATTTCGGAAC; RP: TTCATAGCAAACACAAACTTTTAAGG) were searched in TAIR. The genomic DNA of the sgr2 mutant was extracted from plant leaves with a Plant DNA Isolation Kit (FOREGENE, Chengdu, China). The three-primer reaction [64] was used to identify the sgr2 mutant; DNA was used as the template; LP-BP, LP-RP, and RP-BP combinations were used as primers; and PCR amplification of each strain was respectively carried out.
2.9. Determination of Plant Physiological Indicators
Chlorophyll-a and chlorophyll-b were extracted in acetone (80%) and estimated from mature leaves following the standard method of Arnon [65]. The level of lipid peroxidation was determined by the content of malondialdehyde (MDA) in the leaf segment homogenates prepared in 10 mL of 10% trichloroacetic acid (TCA) containing 0.5% thiobarbituric acid (TBA) as described previously [66]. The antioxidant enzyme activity, including superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT), were measured using the Total Superoxide Dismutase (T-SOD) assay kit, Peroxidase assay kit, and Catalase (CAT) assay kit (NJJCBIO, Jiangsu, China), respectively. Samples were prepared according to the manufacturer’s instructions.
2.10. Statistical Analysis
All data in the study were obtained from the average of three repeats. For all experiments in this study, SPSS 25.0 (SPSS, Chicago, IL, USA) was used for the statistical analysis of the data. Differences at the 0.05 level in the data were analyzed using one-way ANOVA with Duncan’s test and Student’s t-test. Data shown here are the mean with standard error (SE).
3. Results
3.1. Phenotype of Tomato Leaves under Cadmium Stress
Grown tomato plants were treated with different concentrations of CdCl2. After 3 days of treatment, there was no difference in the tomato leaves under Cd stress. Compared with other leaves, the first and second leaves of tomato were slightly albino, which was a normal development phenomenon. After 5 days of treatment, the first leaves under the 25 µM and 50 µM CdCl2 treatments all turned yellow while the first true leaves under the 100 µM treatment fell off, wilted and curled, and were dark brown. All of the second leaves of both groups turned yellow; the yellowing degree of the third leaves gradually increased with the increase in the treatment concentration. Compared with the control, the fourth leaves of the treatment group were slightly chlorotic with a tendency to yellow. After 7 days treatment, the first (not shown) and the second leaves withered and completely fell off. The third leaves of all groups tuned yellow; the leaves with 50 µM and 100 µM CdCl2 had the highest degree of yellowing among them, while the fourth leaf of the CK group still stayed green but showed a yellowing trend. The 25 µM group basically showed the same results as the control group; The uppermost leaflet of the leaves began to turn yellow under the treatment of 50 µM and the yellowing degree of the leaflet at the same position increased when the concentration reached 100 µM (Figure 1A).

Figure 1.
Phenotype of Solanum lycopersicum L. leaves under cadmium treatment. (A) Phenotype of leaves yellowing. (B) Ultrastructure of tomato leaves. (a,b) Control groups; (c–h) cadmium treatment groups; (c,d) under 25µM CdCl2 treatment; (e,f) under 50µM CdCl2 treatment; (g,h) under 100 µM CdCl2 treatment. (a,c,e,g) Samples of the third leaves after treatment; (b,d,f,h) samples of the fourth leaves after treatment. Abbreviations: ch, chloroplasts; g, granum thylakoid; sg, starch grain. All bars = 500 nm.
3.2. Ultrastructure of Tomato Leaves
We observed the ultrastructure of chloroplasts to analyze the effects of CdCl2 on chloroplast development. As shown in Figure 1B, the chloroplasts of the third and fourth leaves of the tomato leaves in the control group had an elongated spindle shape and clung to the cell wall and the structure inside was complete and oval; stroma lamella were connected and the thylakoids were neatly stacked and contained a few large starch grains with clear veins (Figure 1B(a,b)). Under the 25 µM CdCl2 treatment, the chloroplast thylakoids of the third leaf were stacked neatly and connected to each other and also had an elongated spindle shape, but the veins were blurred compared with the control group; there were no significant changes in the chloroplasts of the fourth leaf under the same concentration treatment, which was basically the same results as those of the control group (Figure 1B(c,d)). When the Cd treatment concentration reached 50 µM, the chloroplasts of the third leaf no longer adhered to the cell wall and the structure changed significantly compared with the control group; the thylakoids were not stacked tightly, the stroma lamellae were expanded and most of them were fused with each other in a disordered arrangement and the starch grain disappeared; the number of grana lamellae in the chloroplasts of the fourth leaf were significantly reduced and they had a blurred structure (Figure 1B(e,f)). The structures of the chloroplasts of the third leaf were basically separated from the cell wall and had a shape close to spherical under the 100 µM CdCl2 treatment; the thylakoid arrangement was completely disordered, which made it difficult to distinguish the internal structure; plastid globules and many vacuoles of different sizes emerged and the grana lamellae were scattered and in a state on the verge of disintegration. While the fourth chloroplasts were separated from the cell wall, plastid globules were produced inside and only a very small amount of thylakoids were stacked in the grana lamellae, with the interstitium swelled, the structure disorganized, and veins blurred (Figure 1B(g,h)).
3.3. Analysis of SlSGR2 Sequence
The full-length of the cDNA sequence of the SlSGR2 gene was cloned from “Hongluocheng” (Solyc08g080090.2), which contained an open reading frame (ORF) with a length of 819 bp encoding 272 amino acids (Figure 2A). According to the prediction of ExPASy ProtParam, the molecular structure of SlSGR2 was C1363H2142N376O397S12, the relative molecular mass was 30.533 KD, the protein isoelectric point (pI) was 8.85, the total number of negatively charged residues was 25, the total number of positively charged residues was 31, and the average hydropathicity was −0.337. The fat coefficient was 82.35, the estimated half-life was 30 h, and the protein instability coefficient was 59.96, which made it an unstable protein. The protein was composed of 20 different amino acids (Figure 2B). SOPMA was used to predict the secondary structure of the SlSGR2 protein, which showed that in the secondary structure of SlSGR2, the α-helix accounted for 27.57%, the β-turn was 2.57%, the extended chain accounted for 14.71%, and the random coil accounted for the largest portion (55.15%). The prediction results showed that the tertiary structure of the SlSGR2 protein had different degrees of helices and folding (Figure 2C). The amino acid sequence of SlSGR2 was aligned with the homologous sequences from Arabidopsis thaliana L. (NP_001328989.1, AtNYE1/SGR1), A. thaliana L. (NP_192928.2, AtSGRL), Capsicum annuum L. (NP_001311847.1, CaSGR), Medicago truncatula Linn. (XP_003611177.1, MtSGR), Pisum sativum L. (A7VLV1.1, PsSGR), Oryza sativa L. (XP_015611682.1, OsSGR), Zea mays L. (NP_001105770.2, ZmSGR1), Z. mays L. (AAW82957.1, ZmSGR2), Glycine max Linn. (NP_001238357.1, GmSGR1), G. max Linn. (NP_001236690.1, GmSGR2), and Sorghum bicolor L. (AAW82958.1, SbSGR). Among them, StSGRL shared the highest identity with SlSGR2 of 95.59% followed by NtSGR at 81.99% (Figure 3). The phylogenetic tree of the SGRs was created using MEGA7.0 with neighbor-joining (NJ), which showed that SGRs existed in both the monocotyledonous and dicotyledonous plants and were divided into two subgroups. However, the SGR of pea did not share a common ancestor with other dicotyledonous plants but had a relatively close genetic distance with soybean, which is in the same family. The SlSGR2 belonged to the same branch as the potato SGRL and had a similar genetic distance to the MtSGR and AtSGRL (Figure 4). The subcellular localization prediction proved that SlSGR2 was a chloroplast-localized protein (Figure 5A). A 2 kb upstream sequence of the coding regions of the SlSGR2 gene was selected as the promoter region for cis-element analysis. Four categories of cis-acting elements were identified from the promoter of SlSGR2 gene, which included hormone-responsive elements, light-responsive elements, environmental-stress-related elements, and anaerobic-induction elements. The hormone-response element contained five: ABRE (abscisic acid responsiveness); ERE and W-box (ethylene-responsive element); P-box (gibberellin-responsive element); and TCA (salicylic acid responsiveness). There were five elements that might respond to environmental stress: MYB and MYC (drought-, salt-, and high-temperature-responsive elements); STRE (abiotic-stress-responsive element); TC-rich (defense and stress-responsive element); and WRE (low-temperature-responsive element). Five elements related to light response were found: AE-box, Box4, G-box, GT1-motif, and MRE. Moreover, an element ARE, which was responsive to anaerobic induction, was discovered (Figure 6).

Figure 2.
Sequence assays of SlSGR2: (A) nucleotide and gene sequences of SlSGR2 coding regions; (B) amino acid composition of SlSGR2; (C) tertiary structure of SlSGR2 protein.

Figure 3.
Homologous multiple alignment of amino acid sequences.

Figure 4.
Phylogenetic tree of SlSGR2 proteins.
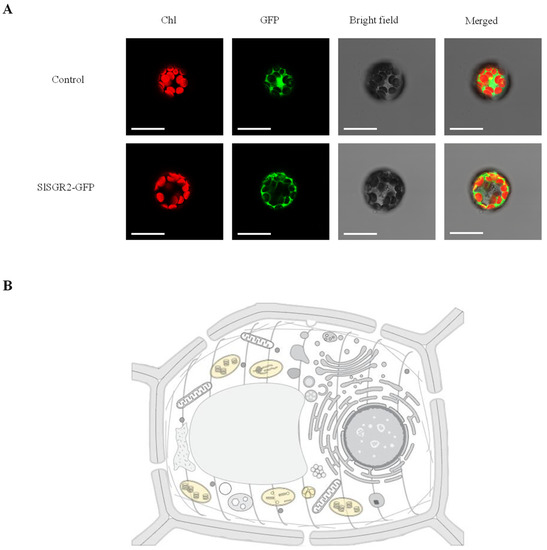
Figure 5.
Subcellular localization of analysis of SlSGR2. (A) Transient expression of SlSGR2-GFP fusion proteins in Arabidopsis mesophyll protoplasts. Scale bar = 10 μm. (B) Prediction of the SlSGR2 protein’s subcellular localization. The yellow part is the predicted location.
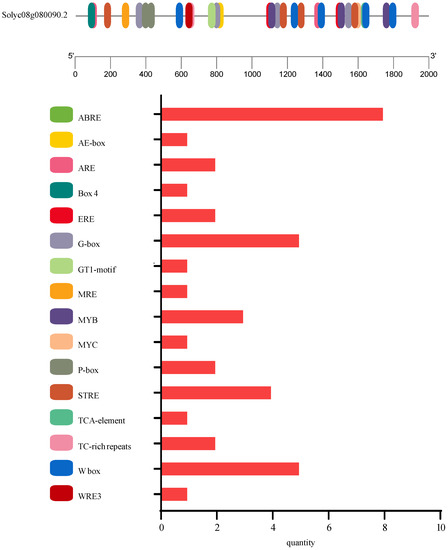
Figure 6.
Visualization of cis-acting elements in the promoter sequence of SlSGR2.
3.4. Subcellular Localization Analysis of SlSGR2
To detect the subcellular localization of SlSGR2, the empty vector (control) and the SlSGR2-GFP vector were transformed into Arabidopsis mesophyll protoplasts, respectively. The results indicated that the SlSGR2 was localized to the chloroplast (Figure 5B).
3.5. Expression Analysis of SlSGR2 under Cadmium Stress
As shown in Figure 7, the relative expression level of the SlSGR2 gene in leaves and roots were upregulated significantly under cadmium treatment. The relative expression level of SlSGR2 was obviously increased at 6 h under the 25 μM CdCl2 Cd treatment in leaves(7.5-fold of that at 0 h)then decreased at 9 h, then reached the highest level at 12 h (11.6-fold of that at 0 h) The expression level of SlSGR2 in roots exhibited an upward trend under the concentration after treatment, showing significant increases at 9 h and 12 h (4.9- and 15.6-fold that at 0 h, respectively) (Figure 7A,B). Under the 50 μM CdCl2 treatment, the relative expression level of SlSGR2 exhibited increases at 6 h, 9 h, and 12 h that were 12.5-, 19.7- and 10.45-fold higher, respectively; the relative expression level of that in roots was significantly increased at 3 h and 6 h (12.6- and 11.3-fold, respectively. The expression level was downregulated compared to previously, but was still higher than that at 0 h; the expression level was obviously increased at 12 h compared to 0 h (8-fold) (Figure 7C,D). Under the treatment of 100 μM CdCl2, the relative expression level of SlSGR2 was decreased at 3 h and 6 h (0.5- and 0.9-fold, respectively), then increased remarkably at 9 h and 12 h (9.5- and 8.4-fold), respectively. The relative expression level in roots under the 100 μM CdCl2 treatment were all elevated significantly at 3 h, 6 h, 9 h, and 12 h (5.8-, 7.1-, 9.2-, and 19.4-fold, respectively) (Figure 7E,F). In summary, these results showed that SlSGR2 was responsive to cadmium stress and that the leaves of tomato responded significantly to the 50 µM CdCl2 treatment.
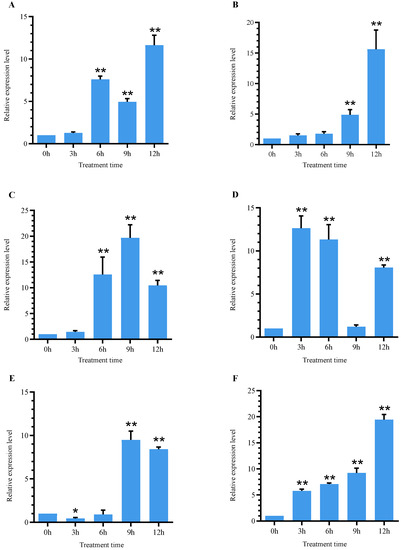
Figure 7.
Expression analysis of SlSGR2 in tomato leaves and roots under cadmium treatment: (A) 25 μM CdCl2 in leaf; (B) 25 μM CdCl2 in root; (C) 50μM CdCl2 in leaf; (D) 50 μM CdCl2 in root; (E) 100 μM CdCl2 in leaf; (F) 100 μM CdCl2 in root. For data statistical significance, Student’s t-test was performed; p < 0.05 was considered statistically significant (* <0.05 and ** <0.01).
3.6. Identification of Transgenic and Mutant of A. thaliana
To further study the function of SlSGR2 gene, transgenic A. thaliana plants were obtained through an Agrobacterium-mediated floral dip transformation and selected using kanamycin. The transgenic A. thaliana lines (OE2-1, OE2-2, OE2-3, OE2-4, OE4-1, OE4-2, OE4-3, OE4-4, OE4-5, and OE4-6) were screened on an MS medium containing kanamycin and timentin. The total leaf genomic DNA was extracted from the wild-type (WT) and homozygous T3 generation transgenic Arabidopsis for subsequent experiments. About 819 bp PCR products were observed in all the transgenic lines based on the PCR amplification. Real-time quantitative reverse transcription PCR of the T3 generation strains was further analyzed with an ABI-7500. The results showed that the relative expression levels of SlSGR2 in the transgenic lines (OE4-1 and OE4-2) of 31 and 35 were significantly higher than that in WT (Figure 8B). This was further confirmation that the SlSGR2 gene was introduced in the WT Arabidopsis and overexpressed. According to the results of the three-primer method, the homozygous plants were confirmed (Figure 8A).

Figure 8.
Phenotypic and physiological analysis of pCAMBIA2300-SlSGR2 transgenic lines under cadmium stress: (A) T-DNA insertion site; (B) expression level of SlSGR2 gene. Student’s t-test was performed; p < 0.05 was considered statistically significant (* <0.05 and ** <0.01); (C) six-week-old A. thaliana phenotype under 1 μM cadmium stress for 14 days; (D) chlorophyll content of A. thaliana; (E) MDA content of A. thaliana; (F) SOD activities of A. thaliana; (G) POD activities of A. thaliana; (H) CAT activities of A. thaliana. Lowercase letters indicate significant (p < 0.05) differences among treatments according to Duncan’s test.
3.7. Overexpression of SlSGR2 Negatively Regulates Chlorophyll Degradation
As shown in Figure 8C, there was no evident difference in phenotype among WT, OE4-1, OE4-2, and sgr2 that grew normally before treatment. After 14 days of cadmium treatment, the leaves of the WT plants turned white and stopped growing, and most plants died. The overexpression lines OE4-1 and OE4-2 were inhibited in growth and development compared with those before treatment, showing chlorotic or even bleached leaves, and the overall plant was short and slender; the sgr2 mutant plants had a similar phenotype to the WT with yellow or albino leaves, and most plants died. For analysis of the function of SlSGR2, we further measured the relevant physiological indicators. The chlorophyll content of the WT was 1.1 mg/g and the chlorophyll content of the sgr2 mutant was 1.3 mg/g after Cd treatment. The chlorophyll content of the overexpression lines OE4-1 and OE4-2 were 2.1 and 1.96 mg/g, respectively, which were significantly higher than those of the wild-type and mutant plants (Figure 8D). The malondialdehyde (MDA) content in the OE4-1 and OE4-2 lines was 22.7 and 24.2 μmol/g, respectively; the MDA content of the wild-type was 55.6 μmol/g; and the MDA content in sgr2 was 53.1 μmol/g, all of which were significantly higher than those of the overexpression strains (Figure 8E). Moreover, we examined the enzyme activities of antioxidant enzymes including SOD, POD, and CAT of the leaves from the transgenic plant lines and the WT plants under cadmium stresses. Under cadmium treatment, the SOD, POD, and CAT activity was significantly higher in the transgenic lines than the WT plants (Figure 8F–H).
4. Discussion
Cadmium is a highly toxic contaminant to plants and negatively affects plant growth [67]. The morphological characteristics of plants are one of the main indicators of metal toxicity in plants. In this experiment, tomato plants were treated with different concentrations of CdCl2 solution; we observed that the yellow phenotype of tomato leaves appeared on the fifth day after cadmium treatment. Among the treatments, the 50 μM CdCl2 treatment group showed the most significant effects compared with the control group. The first leaves of the tomato plant completely fell off on the seventh day after the treatment while the second leaves were dry and withered and completely curled. The second leaves of the 100 μM CdCl2 treatment group turned tawny; the third leaves under the cadmium treatment all turned yellow at 7 d. Cadmium has toxic effects on plants including general growth inhibition and abnormalities, reductions in shoot and root elongation, and leaf curling and yellowing. The symptoms of yellowing, dehydration, and browning of leaves observed in this experiment are common symptoms of toxicity in plants exposed to cadmium [68]. The chloroplast is an organelle that performs photosynthesis that is composed of two membranes and is ellipsoid-shaped. There are thylakoids stacked inside to form tightly arranged grana. The grana are interconnected tightly, indicating that the more complete the structure of the chloroplast, the better the biosynthetic function can be used for photosynthesis. According to the observation results of the submicroscopic structure of tomato leaves under different concentrations of Cd stress, we found that with the increase in CdCl2 concentration, the internal structure of the chloroplast gradually deteriorated. Leaves are important sites for plant photosynthesis and are sensitive to metal stress [26]. The chloroplasts of the clover can be severely damaged and on the verge of disintegration. Research has shown that Cd stress inhibits photosynthesis in plants [21]. Early studies also showed that cadmium stress severely disrupted the structural integrity of the chloroplast, mitochondria, and endoplasmic reticulum [69].
In this experiment, the CDS region of the SlSGR2 gene was cloned from tomato; the sequencing results showed that the full length was 819 bp encoding 272 amino acids. Analysis of its protein physicochemical properties showed that the SlSGR2 protein was composed of 20 amino acids; the most abundant ones were leucine (Leu) and serine (Ser). SlSGR is a weakly alkaline labile hydrophilic protein. The structure of a protein determines its function. The analysis of the secondary and tertiary structure of the SlSGR2 protein predicted that it would have a great number of α-helices and random coils. The SGR gene family is highly conserved in plants except algae [48]. The amino acid sequence homology analysis in this study showed that the SlSGR2 protein had similarities with many species; the highest similarity was for StSGRL. According to the phylogenetic tree, the SlSGR2 protein and the StSGRL protein belonged to the same branch in the monocot subfamily, showing a similarity of 91%. All of the SGR family proteins in higher plants are predicted to localize to the chloroplast [48], which suggests that they may function in plastids, most likely in chlorophyll degradation. Subsequent studies found that rice OsSGR [56], AtSGRL [61], AtNYE1/SGR1 [70], and AtSGR2 [52] of Arabidopsis thaliana were localized in the thylakoid membrane, while perennial ryegrass LpSGR were localized in the chloroplast [71]. The results of the SlSGR2 protein localization prediction analysis and subcellular localization showed that the SlSGR2 chloroplast gene was located in the chloroplast, which was consistent with the previous prediction. The core promoter is the smallest stretch of contiguous DNA sequence that enables the RNA polymerase II machinery to accurately initiate transcription [72]. Analysis of the 2000 bp promoter sequence upstream of the coding region of the SlSGR2 gene revealed that there were five cis-acting elements that responded to hormones, light, environmental stress, and anaerobic stress, which indicated that SlSGR2 may be involved in responding to abiotic stress.
Chlorophyll, which is the most important pigment involved in plant photosynthesis, regulates the distribution and utilization of nitrogen in plants. As much as 75% of the nitrogen present in mesophyll cells is in chloroplasts [73]. Moreover, chlorophyll degradation also contributes to fruit ripening and color formation [59] and plays a key role in the biosynthesis of Arabidopsis seed tocopherols [58]. There are two major factors in the decomposition of chlorophyll, one of which mainly occurs in senescent leaves of plants; the process is regulated by well-characterized chlorophyll catabolism genes. The second major factor is influenced by adversity stress and hormones; chlorophyll breakdown can be triggered in various tissues under different conditions such as after pathogen infection [74] and under abiotic stress [75]. A study found that SGR1 was involved in (Pseudomonas Syringae pv. tomato, Pst) DC3000 infection of leaf chlorosis [76]. Several studies in 2014 revealed that the Arabidopsis SGR1 and SGRL genes are involved in the regulation of chlorophyll degradation under salt stress, osmotic (mannitol) stress, and drought stress [52,60,61]. The expression level of the LeSGR1 gene was upregulated in tomato under drought stress [77]. The above results indicated that SGRs proteins were involved in inhibiting chlorophyll degradation under stress. In this experiment, a qRT-PCR assay was used to analyze the expression level of the SlSGR2 gene of tomato under different concentrations of Cd treatment. The results showed that the SlSGR2 gene responded to Cd stress and the expression of this gene was upregulated in leaves and roots after treatment. The expression level of the SlSGR2 gene in tomato roots was significantly upregulated after the 50 μM CdCl2 treatment at 3 h, which was faster than that in leaves, indicating that the toxicity of Cd to plants first affects the roots and then affects the leaves. However, the expression of the SlSGR2 gene was delayed in roots under the 25 µM CdCl2 treatment and in leaves under the 100 µM CdCl2 treatment, possibly due to the low or high concentration of Cd. In summary, we concluded that the SlSGR2 gene responded to cadmium stress and that 50 µM CdCl2 was the appropriate treatment concentration.
The STAY-GREEN (SGR) protein plays an important role in the regulation of plant chlorophyll degradation and senescence [52]. In addition to promoting natural senescence, stress also can promote plant leaf senescence [78].
Overexpression of the AtSGR2 gene in A. thaliana showed that the plants exhibited an evergreen phenotype and that the sgr2-1 mutant had earlier yellowed under natural senescence. The leaves of the SGR1 gene-overexpressing line turned yellow earlier than those of the wild-type, which was opposite to the function of the SGR2 gene [61]. The leaves of SGR2-overexpressed plants stayed green better than the wild-type under abiotic stress [52]. After 14 days of cadmium stress on the wild-type (WT), overexpressed, and mutant A. thaliana lines under the same conditions, we found that the growth states of two the overexpressed lines (OE4-1 and OE4-2) were significantly better than those of WT and sgr2, indicating that the overexpression of the SlSGR2 gene in Arabidopsis improved the tolerance of the plants to cadmium stress. The chlorophyll content of Arabidopsis after cadmium stress showed that the overexpression lines were more green-keeping and more tolerant to cadmium stress than the WT and sgr2 lines. Malondialdehyde (MDA) is an indicator that reflects the effect of free radicals on membrane lipids to produce peroxides after plant cells are injured [79]. The higher the content, the greater the damage. The results showed that the MDA contents of the WT and sgr2 were significantly higher in the transgenic lines than in the WT plants, indicating that the degree of damage was greater than that in the overexpressed lines. When plants are exposed to adverse conditions, they will produce excessive reactive oxygen species (ROS). High concentrations of ROS are very harmful to plants. When the ROS levels exceed cellular defenses, oxidative stress will occur, triggering the plants defense mechanisms and ultimately leading to plant death [80]. SOD, POD, and CAT are the enzymes that can scavenge ROS and protect the plant against oxidative damage [81]. Our results showed that SlSGR2 remarkably increased the activities of the SOD, POD, and CAT enzymes. Overall, the physiological analysis suggested that Arabidopsis plants that overexpressed SlSGR2 outperformed the wild-type plants in various indicators, which was related to the better cadmium tolerance of the transgenic Arabidopsis plants compared with the WT plants.
5. Conclusions
The phenotype observation of tomato leaves under cadmium stress showed that with the increase in treatment time, the degree of yellowing of tomato leaves gradually deepened; the higher the treatment concentration, the greater the degree of yellowing. The ultrastructural observation showed that the higher the concentration of cadmium treatment, the more serious the damage to the chloroplast structure. The STAY-GREEN (SGR) protein played an important role in recruiting CCE for LHCII to aid in the breakdown of chlorophyll. Here, the SlSGR2 gene, which contained 819 bp encoding 272 amino acids, was identified and characterized. The sequence alignment suggested that the SlSGR2 protein was highly conserved among different plant species. An analysis of the cis-acting elements indicated that SlSGR2 may be involved in responding to abiotic stress. The subcellular localization prediction and laser confocal analysis indicated that the SlSGR2 protein was localized to the chloroplast. A qRT-PCR analysis of the cadmium-treated tomato plants showed that the SlSGR2 gene responded to cadmium stress. The heterologous overexpression of SlSGR2 in Arabidopsis thaliana proved the role of SlSGR2 in regulating cadmium stress. These results suggested that the SlSGR2 gene may be involved in Cd-stress-induced plant senescence and may also be a negative regulator of plant chlorophyll degradation.
Author Contributions
Conceptualization and methodology, Y.Z. and J.M.; software, X.D.; validation, Y.Z. and L.Y.; formal analysis, J.M. and X.D.; investigation and resources, Y.Z. and J.M.; data curation, J.M. and X.D.; writing—original draft preparation, J.M and Y.Z.; writing—review and editing, Y.Z. and L.Y.; visualization, J.M and X.D.; supervision, project administration, and funding acquisition, Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by the Applied Technology Research and Development Project of Heilongjiang Province under grant number GA19B201-5.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hechmi, N.; Aissa, N.B.; Abdenaceur, H.; Jedidi, N. Evaluating the phytoremediation potential of Phragmites australis grown in pentachlorophenol and cadmium co-contaminated soils. Environ. Sci. Pollut. Res. Int. 2014, 21, 1304–1313. [Google Scholar] [CrossRef]
- Huang, H.; Yu, N.; Wang, L.; Gupta, D.K.; He, Z.; Wang, K.; Zhu, Z.; Yan, X.; Li, T.; Yang, X.E. The phytoremediation potential of bioenergy crop Ricinus communis for DDTs and cadmium co-contaminated soil. Bioresour. Technol. 2011, 102, 11034–11038. [Google Scholar] [CrossRef] [PubMed]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 402647. [Google Scholar] [CrossRef]
- Yamagata, N.; Shigematsu, I. Cadmium pollution in perspective. Bull. Inst. Public Health 1970, 19, 1–27. [Google Scholar]
- Wang, B.; Lin, C.; Xuan, Z.; Duan, X.; Wu, F. A soil ingestion pilot study for teenage children in China. Chemosphere 2018, 202, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Ma, Y.; Zhu, Y.G.; Zhong, T.; Mcgrath, S.P. Soil Contamination in China: Current Status and Mitigation Strategies. Environ. Sci. Technol. 2015, 49, 750. [Google Scholar] [CrossRef] [PubMed]
- Huaiman, C. Heavy Metal Pollution in Soil-Plant Systems. Pedosphere 1998, 8, 72–75. [Google Scholar]
- Rizwan, M.; Ali, S.; Abbas, T.; Zia-Ur-Rehman, M.; Hannan, F.; Keller, C.; Al-Wabel, M.I.; Ok, Y.S. Cadmium minimization in wheat: A critical review. Ecotoxicol. Environ. Saf. 2016, 130, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.W.; Zia-Ur-Rehman, M.; Zahir, Z.A.; Rinklebe, J.R.; Tack, F.M.G.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef]
- Hallenbeck, W. Human health effects of exposure to cadmium. Birkhäuser 1986, 50, 131–137. [Google Scholar] [CrossRef]
- Hongyan, L.; Weiqin, W.; Aibin, H.; Lixiao, N. Correlation of leaf and root senescence during ripening in dry seeded and transplanted rice. Rice Sci. 2018, 25, 279–285. [Google Scholar] [CrossRef]
- Kobayashi, E.; Suwazono, Y.; Dochi, M.; Honda, R.; Kido, T. Influence of consumption of cadmium-polluted rice or Jinzu River water on occurrence of renal tubular dysfunction and/or Itai-itai disease. Biol. Trace Elem. Res. 2009, 127, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Meharg, A.A.; Norton, G.J.; Deacon, C.; Williams, P.; Islam, M.R. Variation in rice cadmium related to human exposure. Environ. Sci. Technol. 2013, 47, 5613–5618. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, Y.; Mao, W.; Sui, H.; Yong, L.; Yang, D.; Jiang, D.; Zhang, L.; Gong, Y. Dietary cadmium exposure assessment among the Chinese population. PLoS ONE 2017, 12, e0177978. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.A.; Hc, A.; Pmk, B.; Fjz, A. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar]
- EFSA Panel. Cadmium in food—Scientific opinion of the Panel on Contaminants in the Food Chain. EFSA J. 2009, 7, 1–139. [Google Scholar] [CrossRef]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- Hongping, C.; Xinping, Y.; Peng, W.; Zixuan, W.; Ming, L.; Fang-Jie, Z. Dietary cadmium intake from rice and vegetables and potential health risk: A case study in Xiangtan, southern China. Sci. Total Environ. 2018, 639, 271–277. [Google Scholar]
- Shi, T.; Zhang, Y.; Gong, Y.; Haiying, J.; Xiao, W. Status of cadmium accumulation in agricultural soils across China (1975–2016): From temporal and spatial variations to risk assessment. Chemosphere 2019, 230, 136–143. [Google Scholar] [CrossRef]
- Ye, K.; Wu, R. Autumn snow cover variability over northern Eurasia and roles of atmospheric circulation. Adv. Atmos. Sci. 2017, 34, 847–858. [Google Scholar] [CrossRef]
- Andresen, E.; Küpper, H. Cadmium toxicity in plants. Met. Ions Life Sci. 2013, 11, 395–413. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C.; Schat, H. Mechanisms to cope with arsenic or cadmium excess in plants. Curr. Opin. Plant Biol. 2009, 12, 364–372. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 2027–2038. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Vaculík, M.; Martinka, M.; Lisková, D.; Staden, J.V. Cadmium induces hypodermal periderm formation in the roots of the monocotyledonous medicinal plant Merwilla plumbea. Ann. Bot. 2011, 107, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, Z. Physiological mechanism of hypertolerance of cadmium in Kentucky bluegrass and tall fescue: Chemical forms and tissue distribution. Environ. Exp. Bot. 2013, 96, 35–42. [Google Scholar] [CrossRef]
- Baldantoni, D.; Morra, L.; Zaccardelli, M.; Alfani, A. Cadmium accumulation in leaves of leafy vegetables. Ecotoxicol. Environ. Saf. 2016, 123, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J.; Mimura, T. Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ. 2016, 39, 1112–1126. [Google Scholar] [CrossRef]
- Carrier, P.; Baryla, A.; Havaux, M. Cadmium distribution and microlocalization in oilseed rape (Brassica napus) after long-term growth on cadmium-contaminated soil. Planta 2003, 216, 939–950. [Google Scholar] [CrossRef]
- Bačkor, M.; Váczi, P.; Barták, M.; Bud’ová, J.; Dzubaj, A. Uptake, photosynthetic characteristics and membrane lipid peroxidation levels in the lichen photobiont Trebouxia erici exposed to copper and cadmium. Bryologist 2007, 110, 100–107. [Google Scholar] [CrossRef]
- Yi, T.H.; Kao, C.H. Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul. 2004, 42, 227–238. [Google Scholar]
- Toppi, L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Küpper, H.; Küpper, F.; Spiller, M. Environmental relevance of heavy metal-substituted chlorophylls using the example of water plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef]
- Faller, P.; Kienzler, K.; Krieger-Liszkay, A. Mechanism of Cd2+ toxicity: Cd2+ inhibits photoactivation of Photosystem II by competitive binding to the essential Ca2+ site. Biochim. Biophys. Acta 2005, 1706, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Sigfridsson, K.G.; Bernát, G.; Mamedov, F.; Styring, S. Molecular interference of Cd(2+) with Photosystem II. Biochim. Biophys. Acta 2004, 1659, 19–31. [Google Scholar] [CrossRef]
- Horie, Y.; Ito, H.; Kusaba, M.; Tanaka, R.; Tanaka, A. Participation of chlorophyll b reductase in the initial step of the degradation of light-harvesting chlorophyll a/b-protein complexes in Arabidopsis. J. Biol. Chem. 2009, 284, 17449–17456. [Google Scholar] [CrossRef]
- Kusaba, M.; Ito, H.; Morita, R.; Iida, S.; Sato, Y.; Fujimoto, M.; Kawasaki, S.; Tanaka, R.; Hirochika, H.; Nishimura, M.; et al. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell 2007, 19, 1362–1375. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morita, R.; Katsuma, S.; Nishimura, M.; Tanaka, A.; Kusaba, M. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J. 2009, 57, 120–131. [Google Scholar] [CrossRef]
- Meguro, M.; Ito, H.; Takabayashi, A.; Tanaka, R.; Tanaka, A. Identification of the 7-hydroxymethyl chlorophyll a reductase of the chlorophyll cycle in Arabidopsis. Plant Cell 2011, 23, 3442–3453. [Google Scholar] [CrossRef]
- Morita, R.; Sato, Y.; Masuda, Y.; Nishimura, M.; Kusaba, M. Defect in non-yellow coloring 3, an alpha/beta hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J. 2009, 59, 940–952. [Google Scholar] [CrossRef]
- Schelbert, S.; Aubry, S.; Burla, B.; Agne, B.; Kessler, F.; Krupinska, K.; Hörtensteiner, S. Pheophytin pheophorbide hydrolase (pheophytinase) is involved in chlorophyll breakdown during leaf senescence in Arabidopsis. Plant Cell 2009, 21, 767–785. [Google Scholar] [CrossRef]
- Pruzinská, A.; Anders, I.; Aubry, S.; Schenk, N.; Tapernoux-Lüthi, E.; Müller, T.; Kräutler, B.; Hörtensteiner, S. In vivo participation of red chlorophyll catabolite reductase in chlorophyll breakdown. Plant Cell 2007, 19, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Hirashima, M.; Satoh, S.; Tanaka, A. The Arabidopsis-accelerated cell death gene ACD1 is involved in oxygenation of pheophorbide a: Inhibition of the pheophorbide a oxygenase activity does not lead to the “stay-green” phenotype in Arabidopsis. Plant Cell Physiol. 2003, 44, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Park, S.Y.; Paek, N.C. The Divergent Roles of STAYGREEN (SGR) Homologs in Chlorophyll Degradation. Mol. Cells 2015, 38, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Armstead, I.; Donnison, I.; Aubry, S.; Harper, J.; Hörtensteiner, S.; James, C.; Mani, J.; Moffet, M.; Ougham, H.; Roberts, L.; et al. Cross-species identification of Mendel’s I locus. Science 2007, 315, 73. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.W.; Lee, Y.J.; Koh, H.J.; Lee, B.M.; Nam, Y.W.; Paek, N.C. Isolation, characterization, and mapping of the stay green mutant in rice. Theor. Appl. Genet. 2002, 104, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; An, K.; Liao, Y.; Zhou, X.; Cao, Y.; Zhao, H.; Ge, X.; Kuai, B. Identification of a novel chloroplast protein AtNYE1 regulating chlorophyll degradation during leaf senescence in Arabidopsis. Plant Physiol. 2007, 144, 1429–1441. [Google Scholar] [CrossRef]
- Jiang, H.; Li, M.; Liang, N.; Yan, H.; Wei, Y.; Xu, X.; Liu, J.; Xu, Z.; Chen, F.; Wu, G. Molecular cloning and function analysis of the stay green gene in rice. Plant J. 2007, 52, 197–209. [Google Scholar] [CrossRef]
- Barry, C.S.; McQuinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-flesh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol. 2008, 147, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Guo, Y.; Kuai, B. Isolation and characterization of a chlorophyll degradation regulatory gene from tall fescue. Plant Cell Rep. 2011, 30, 1201–1207. [Google Scholar] [CrossRef]
- Zhou, C.; Han, L.; Pislariu, C.; Nakashima, J.; Fu, C.; Jiang, Q.; Quan, L.; Blancaflor, E.B.; Tang, Y.; Bouton, J.H.; et al. From model to crop: Functional analysis of a STAY-GREEN gene in the model legume Medicago truncatula and effective use of the gene for alfalfa improvement. Plant Physiol. 2011, 157, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Li, C.; Li, W.; Wang, Z.; Zhou, Z.; Shen, Y.; Wu, M.; Wu, Y.; Li, G.; Kong, L.A.; et al. Concerted evolution of D1 and D2 to regulate chlorophyll degradation in soybean. Plant J. 2014, 77, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Park, S.Y.; Kim, Y.S.; Wang, S.H.; Yoo, S.C.; Hörtensteiner, S.; Paek, N.C. Arabidopsis STAY-GREEN2 is a negative regulator of chlorophyll degradation during leaf senescence. Mol. Plant 2014, 7, 1288–1302. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.S.; Liu, V.; Thomson, W.W.; Platt, K.; Walling, L.L. The Impact of Chlorophyll-Retention Mutations, d1d2 and cyt-G1, during Embryogeny in Soybean. Plant Physiol. 1995, 107, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Guiamét, J.J.; Schwartz, E.; Pichersky, E.; Noodén, L.D. Characterization of Cytoplasmic and Nuclear Mutations Affecting Chlorophyll and Chlorophyll-Binding Proteins during Senescence in Soybean. Plant Physiol. 1991, 96, 227–231. [Google Scholar] [CrossRef]
- Clerkx, E.J.; Vries, H.B.; Ruys, G.J.; Groot, S.P.; Koornneef, M. Characterization of green seed, an enhancer of abi3-1 in Arabidopsis that affects seed longevity. Plant Physiol. 2003, 132, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Flanagan, A.M.; Spencer, M.S. Ethylene Production during Development of Mustard (Brassica juncea) and Canola (Brassica napus) Seed. Plant Physiol. 1994, 106, 601–606. [Google Scholar] [CrossRef]
- Delmas, F.; Sankaranarayanan, S.; Deb, S.; Widdup, E.; Bournonville, C.; Bollier, N.; Northey, J.G.; McCourt, P.; Samuel, M.A. ABI3 controls embryo degreening through Mendel’s I locus. Proc. Natl. Acad. Sci. USA 2013, 110, E3888–E3894. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, T.; Ren, G.; Hörtensteiner, S.; Zhou, Y.; Cahoon, E.B.; Zhang, C. Chlorophyll degradation: The tocopherol biosynthesis-related phytol hydrolase in Arabidopsis seeds is still missing. Plant Physiol. 2014, 166, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A STAY-GREEN protein SlSGR1 regulates lycopene and β-carotene accumulation by interacting directly with SlPSY1 during ripening processes in tomato. New Phytol. 2013, 198, 442–452. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Lee, S.H.; Kim, Y.S.; Park, O.K.; Hörtensteiner, S.; Paek, N.C. Delayed degradation of chlorophylls and photosynthetic proteins in Arabidopsis autophagy mutants during stress-induced leaf yellowing. J. Exp. Bot. 2014, 65, 3915–3925. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Kim, D.; Kim, Y.S.; Hörtensteiner, S.; Paek, N.C. Arabidopsis STAYGREEN-LIKE (SGRL) promotes abiotic stress-induced leaf yellowing during vegetative growth. FEBS Lett. 2014, 588, 3830–3837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, H.; Huang, X.; Xing, J.; Yao, J.; Yin, T.; Jiang, J.; Wang, P.; Xu, B. STAYGREEN-mediated chlorophyll a catabolism is critical for photosystem stability during heat-induced leaf senescence in perennial ryegrass. Plant Cell Environ. 2022, 45, 1412–1427. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J.; Kirk, D.D.; Walmsley, A.M. Tomato (Lycopersicum esculentum). Methods Mol. Biol. 2006, 343, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dongfang, L.; Xiaole, C.; Naixing, W. Determination of Malondialdehyde in Biological Samples by 2.5th Order Differential Voltammetry. Prog. Biochem. Biophys. 1997, 24, 361–363. [Google Scholar]
- Yazdi, M.; Kolahi, M.; Mohajel Kazemi, E.; Goldson Barnaby, A. Study of the contamination rate and change in growth features of lettuce (Lactuca sativa Linn.) in response to cadmium and a survey of its phytochelatin synthase gene. Ecotoxicol. Environ. Saf. 2019, 180, 295–308. [Google Scholar] [CrossRef]
- Tran, T.A.; Popova, L.P. Functions and toxicity of cadmium in plants: Recent advances and future prospects. Doga Turk. J. Bot. 2013, 37, 1–13. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Cadmium Disrupts Subcellular Organelles, Including Chloroplasts, Resulting in Melatonin Induction in Plants. Molecules 2017, 22, 1791. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Schelbert, S.; Park, S.Y.; Han, S.H.; Lee, B.D.; Andrès, C.B.; Kessler, F.; Hörtensteiner, S.; Paek, N.C. STAY-GREEN and chlorophyll catabolic enzymes interact at light-harvesting complex II for chlorophyll detoxification during leaf senescence in Arabidopsis. Plant Cell 2012, 24, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Yu, G.; Li, H.; Xie, Z.; Wen, W.; Zhang, J.; Huang, B. Knockdown of STAYGREEN in Perennial Ryegrass (Lolium perenne L.) Leads to Transcriptomic Alterations Related to Suppressed Leaf Senescence and Improved Forage Quality. Plant Cell Physiol. 2019, 60, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; Kadonaga, J.T. The RNA polymerase II core promoter: A key component in the regulation of gene expression. Genes Dev. 2002, 16, 2583–2592. [Google Scholar] [CrossRef]
- Peoples, M.B.; Dalling, M.J. The interplay between proteolysis and amino acid metabolism during senescence and nitrogen reallocation. In Senescence and Aging in Plants; Academic Press: San Diego, CA, USA, 1988; pp. 181–217. [Google Scholar]
- Mur, L.A.; Aubry, S.; Mondhe, M.; Kingston-Smith, A.; Gallagher, J.; Timms-Taravella, E.; James, C.; Papp, I.; Hörtensteiner, S.; Thomas, H.; et al. Accumulation of chlorophyll catabolites photosensitizes the hypersensitive response elicited by Pseudomonas syringae in Arabidopsis. New Phytol. 2010, 188, 161–174. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Mecey, C.; Hauck, P.; Trapp, M.; Pumplin, N.; Plovanich, A.; Yao, J.; He, S.Y. A critical role of STAYGREEN/Mendel’s I locus in controlling disease symptom development during Pseudomonas syringae pv tomato infection of Arabidopsis. Plant Physiol. 2011, 157, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.L.; Deng, L.; Yan, B.; Pan, Y.; Luo, M.; Chen, X.Q.; Hu, T.Z.; Chen, G.P. Silencing of the LeSGR1 gene in tomato inhibits chlorophyll degradation and exhibits a stay-green phenotype. Biol. Plant. 2011, 55, 27–34. [Google Scholar] [CrossRef]
- Pic, E.; de La Serve, B.T.; Tardieu, F.; Turc, O. Leaf senescence induced by mild water deficit follows the same sequence of macroscopic, biochemical, and molecular events as monocarpic senescence in pea. Plant Physiol. 2002, 128, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.Q.; Liu, Y.F.; Liu, P.; Lei, G.; He, S.J.; Ma, B.; Zhang, W.K.; Zhang, J.S.; Chen, S.Y. Receptor-like kinase OsSIK1 improves drought and salt stress tolerance in rice (Oryza sativa) plants. Plant J. 2010, 62, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Gu, C.; Zhang, T.; Zhang, Q.; Yu, Q.; Jiang, J.; Liu, G. Functional Study of BpPP2C1 Revealed Its Role in Salt Stress in Betula platyphylla. Front. Plant Sci. 2021, 11, 617635. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).