Abstract
Walnut (Juglans regia L.) plants typically flower after eight to ten years of juvenile growth. Precocious germplasm, also known as early-flowering or early-mature genotypes, have shortened juvenile phases of one to two years and are therefore crucial for enhancing breeding efficiency. However, such precocious germplasms are very limited. Here, we isolated and characterized the key flowering-time gene FLOWERING LOCUS C (FLC) in the precocious walnuts of the Xinjiang Uygur Autonomous Region. Sequence alignment showed that Juglans regia FLC (JrFLC)contained a conserved MINICHROMOSOME MAINTENANCE 1 (MCM1), AGAMOUS (AG), DEFICIENS (DEF), and SERUM RESPONSE FACTOR (SRF) (MADS)-box domain. Analysis of an FLC–green fluorescent protein (GFP) fusion protein revealed that JrFLC was localized to the nucleus. Gene expression analysis showed that JrFLC was specifically expressed during the bud dormancy stage of precocious walnut, and that expression levels gradually decreased as the ambient temperature warmed. Exogenous JrFLC overexpression in Arabidopsis thaliana delayed flowering and increased the total leaf number, suggesting a similar function of JrFLC as a floral repressor in walnut and in other plants. Together, these results showed that JrFLC played an important role in regulating the floral transition of Xinjiang precocious walnut. Further studies, including a detailed characterization of JrFLC, are expected to validate JrFLC as a strong target for genetic improvement in flowering time in walnut.
1. Introduction
Walnut, also known as Persian walnut (Juglans regia L.; 2n = 2x = 32), originate from Central Asia [1]. It is as a nut crop that is rich in a variety of microelements and polyphenols [2,3], but has additional value in medicine and other industrial fields [4,5,6]. Walnut trees have a relatively long juvenile stage (defined as the time from the vegetative to the reproductive phase), usually taking 8–10 years to undergo floral transition. In plants, the floral transition is regulated by several exogenous and endogenous stimuli to ensure the correct timing [7,8]. In the model plant Arabidopsis thaliana, flowering is controlled by various pathways including the photoperiod, vernalization, autonomous, and gibberellin pathways. The critical reproductive phase of flowering is regulated by a network of flowering genes. The most important and well-studied among them are FLOWERING LOCUS C (FLC) and FLOWERING LOCUS T (FT) [9,10]. Some walnut germplasms have the short-juvenile (precocious) trait, which is stably inherited and has been successfully applied to shorten the breeding cycle in selecting new varieties. However, the floral transition is poorly understood in walnut due to a genetic bottleneck that has limited the number of precocious walnut germplasm.
The Xinjiang Uygur Autonomous Region of China, which is near the region in Central Asia in which walnut originated, has a large population of previously unutilized wild-type walnut resources. These wild germplasms are naturally preserved in situ in Gongliu County, at the northern portion of the Tianshan Mountains. Southern Xinjiang contains ancient walnut trees that have grown for hundreds to thousands of years. The main cultivated varieties are Xinjiang precocious thin-skinned varieties, which are abundant in the Xinjiang region [11,12,13]. A previous study from our lab revealed the genetic factors underlying floral transition in early-flowering walnut using RNA sequencing and microRNA sequencing [12]. In this brief report, we isolated the key floral transition gene FLC and measured its dynamic expression levels at multiple growth stages. Furthermore, we confirmed its molecular function via transgenic expression in wild type (Col-0) of A. thaliana. The discovery of FLC-driven precocious germplasm will allow us to answer critical questions about the floral transition and could be applied in future breeding efforts to shorten juvenility in Xinjiang walnuts.
2. Materials and Methods
2.1. Walnut Germplasm
The precocious walnut varieties ‘Xinwen 179′ and ‘Xinwen 81′ were planted in the field in the Xinjiang Uygur Autonomous Region, China (40°32′27.74239″ N, 81°18′3.40166″ E, altitude 957.37 m). Floral buds were collected from each variety during the growth stage in 2021 and 2022, with at least three biological replicates per genotype. Samples were flash-frozen in liquid nitrogen, then stored at −80 °C prior to further analyses.
2.2. Walnut Morphological Analysis
Walnut buds were fixed in formaldehyde–alcohol–acetic acid (FAA), then dehydrated in an alcohol gradient. After dehydration, the samples were treated with a xylene gradient (2:1 alcohol:xylene for 20 min, 1:2 alcohol:xylene for 20 min, then 100% xylene for 10 min) to render the samples transparent. These samples were embedded in paraffin, sectioned with an RM2235 microtome (Leica, Wetzlar, Germany), and stained with fast green (1% w/w) and safranin (1% w/w). The samples were observed and photographed using an Olympus DP71 microscope (Olympus, Tokyo, Japan).
2.3. RNA Extraction
Total RNA was extracted from walnut buds with the RNA Easy Fast Plant Tissue Kit (TIANGEN, Beijing, China) as previously described [12]. RNA concentration and purity were detected with a NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA) and agarose gel electrophoresis. Only high-integrity RNA was used in further experiments.
2.4. Identification of a Putative FLC Protein in Xingjiang Walnut
In plants, FLC belongs to the MINICHROMOSOME MAINTENANCE 1 (MCM1), AGAMOUS (AG), DEFICIENS (DEF), and SERUM RESPONSE FACTOR (SRF) (MADS)-box gene family. FLC-like proteins with a MADS domain were identified in the walnut genome [14] via BLAST. Other candidates were identified with the NCBI Conserved Domain Search database (NCBI CDD) (https://www.ncbi.nlm.nih.gov/cdd/, accessed on 2 November 2022) and the MultAlin software [15] (http://multalin.toulouse.inra.fr/multalin/, accessed on 25 November 2022) based on the presence of a MADS-box domain and AtFLC protein sequence.
2.5. Subcellular Localization
To construct an FLC–green fluorescent protein (GFP) fusion protein, the full-length coding sequence (CDS) of FLC was inserted between the 35S promoter and the sequence encoding GFP in the pMDC85-GFP vector. The subcellular localization of FLC in Arabidopsis protoplasts was determined as previously described [16]. Briefly, protoplasts were isolated from 14-day-old Arabidopsis seedlings and the protoplasts were transformed with the JrFLC-GFP construct. GFP fluorescent signals were observed and images captured on an LSM 700 confocal laser scanning microscope (Zeiss, Oberkochen, Germany).
2.6. Quantitative Reverse Transcription (qRT)-PCR
qRT-PCR was performed with SYBR Green Master mix on a Light Cycle 96 Real-Time PCR system (Roche, Basel, Switzerland). Relative gene expression levels were calculated with the 2−ΔΔCt method [17], using our previously published internal reference gene [12]. Primers of qRT-PCR are shown in Table S1.
2.7. Plasmid Construction and Arabidopsis Transformation
To generate the 35Spro:JrFLC plasmid, the full-length JrFLC CDS without the stop codon was placed into the pK2GW7 vector using Gateway technology. The vector was inserted into Agrobacterium tumefaciens GV3101, and Arabidopsis plants were stably transformed with an Agrobacterium-mediated floral dip method [18]. Flowering time and total leaf number were measured in transgenic 35Spro:JrFLC lines. Downstream target gene expression was determined with qRT-PCR using previously published primers for Arabidopsis FT, SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1), and LEAFY (LFY) [19]. For measurement of flowering time and total leaf number, at least 10 Arabidopsis plants for each line were scored. Significant differences between the means of the indicated genotypes were calculated by a two-tailed Student’s t-test (Excel 2019, accessed on 15 November 2022).
3. Results and Discussion
3.1. JrFLC Isolation and Expression in Precocious Walnut
In plants, the critical reproductive phase (flowering) is regulated by a network of flowering genes, including FLC [9,10]. The precocious walnut varieties found in Xinjiang have very short juvenility, typically blooming and bearing fruit within 3 years; some even flower within a single year [12]. Based on the known essential roles of FLC homologs in plants, we hypothesized that the key regulatory gene FLC may contribute to the short juvenility of Xinjiang precocious walnuts, and that this locus could be used to shorten breeding cycles. Thus, we isolated FLC and explored FLC expression patterns at several growth stages of precocious walnut.
We first cloned the full-length CDS of FLC from the precocious variety ‘Xinwen 81’ (Table S1). Sequence alignment showed that JrFLC had moderate similarity with AtFLC, and we identified a conserved N-terminal MADS-box domain in these two proteins (Figure 1A). Next, we conducted a subcellular localization assay. Protoplasts isolated from the leaves of 14-day-old Arabidopsis seedlings were transformed with the JrFLC-GFP construct. An examination with a confocal laser scanning microscope showed that the JrFLC-GFP fusion protein was localized to the nucleus (Figure 1B), comparable to FLC proteins in Arabidopsis [20] and other species. The conserved MADS-box domain and consistent subcellular localization indicated that JrFLC may have a similar molecular function as previously characterized plant FLCs.
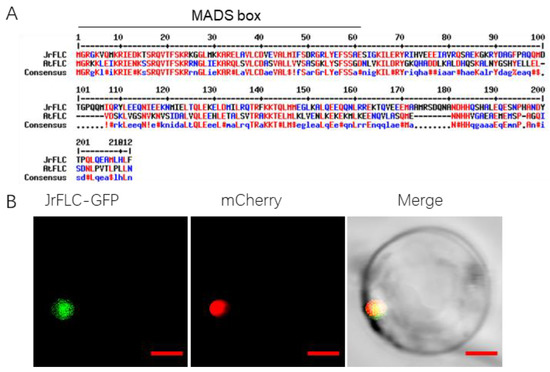
Figure 1.
(A) Sequence alignment of Juglans regia and Arabidopsis thaliana FLC proteins. The N-terminal regions of JrFLC and AtFLC showed a conserved MINICHROMOSOME MAINTENANCE 1 (MCM1), AGAMOUS (AG), DEFICIENS (DEF), and SERUM RESPONSE FACTOR (SRF) (MADS)-box domain. Sequence identity between JrFLC and AtFLC was 45.5%. (B) Subcellular localization of JrFLC. JrFLC tagged with green fluorescent protein (GFP) co-localized with a nuclear marker. Bars in (B) were 50 μm.
To understand the dynamics of JrFLC expression throughout precocious walnut development, we collected samples at several growth stages. We examined overall bud morphology during its formation in precocious walnut (Figure 2A–D) and further assessed internal bud morphology at two key developmental stages, the dormant bud stage and the floral bud stage (Figure 2E,F). In apple, FLC is subject to dynamic regulation during the growth stage. There is some evidence that it is accumulated during the bud dormancy stage due to epigenetic modifications [21]. We hypothesized that JrFLC may have comparable expression patterns in precocious walnut. This hypothesis was tested by analyzing JrFLC expression at multiple stages via qRT-PCR. JrFLC was most highly expressed in the dormant buds, and gradually decreased when ambient temperatures increased (Figure 2G and Figure S1). These results indicated that JrFLC expression was responsive to temperature signals, which may therefore have an impact on precocious walnut germplasm.
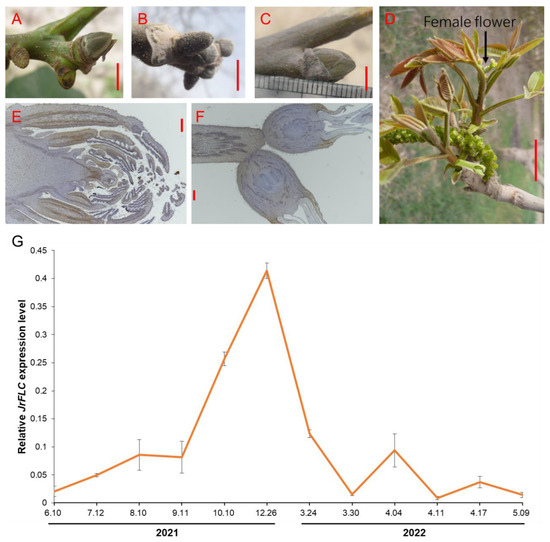
Figure 2.
FLC accumulation in precocious walnut variety ‘Xinwen 81’ during bud dormancy. (A–D) Representative images of buds at several growth stages of precocious walnut: (A) bud differentiation, (B) bud dormancy, (C) bud sprout, and (D) floral formation and leaf expansion. (E–F) Internal morphological analysis of (E) dormant buds and (F) floral buds. (G) Dynamic expression levels of JrFLC in ‘Xinwen 81’. Bars in (A–C) were 0.5 cm; bar in (D) was 1 cm; bars in (E,F) were 50 nm. For qRT-PCR assays, bars in (G) indicated the standard deviations.
3.2. JrFLC Overexpression Delayed Flowering in Arabidopsis
To analyze the molecular function of JrFLC, we created transgenic Arabidopsis plants overexpressing JrFLC (35Spro:JrFLC) (Figure 3A). We first verified the successful construction of the overexpression lines by quantifying FLC expression patterns in transgenic plants (Figure 3B). We then analyzed the phenotypes of the validated 35Spro:JrFLC plants. Compared to the wild-type Columbia-0 plants, JrFLC overexpressors showed delayed flowering and increased total leaf number (Figure 3C,D). Moreover, JrFLC overexpression reduced the expression levels of downstream floral transition genes, including AtFT, AtLFY, and AtSOC1 (Figure 3E). Together, these results indicated that JrFLC and AtFLC had similar functions as floral repressors.
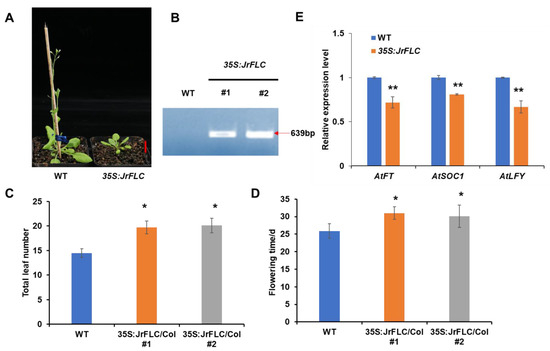
Figure 3.
Functional validation of Juglans regia FLC via overexpression in Arabidopsis. (A) Representative image of wild-type (WT) and JrFLC-overexpression (35Spro:JrFLC) plants of the same growth stage. (B) Validation of JrFLC overexpression in transgenic plants via quantitative reverse transcription PCR. (C,D) Quantification of (C) total leaf number and (D) flowering time in WT and 35Spro:JrFLC Arabidopsis plants. (E) Expression levels of flowering genes downstream of FLC (FT, SOC1, and LFY) in WT and 35Spro:JrFLC Arabidopsis plants. Bar in (A) was 2 cm. At least 10 Arabidopsis plants for each line were scored with flowering time and total leaf number; significant differences between the means of the indicated genotypes were revealed by a two-tailed Student’s t-test (* p < 0.05; ** p < 0.01). For qRT-PCR assays and measurement of flowering time, bars indicated the standard deviations.
Walnut trees are known to have originated in Central Asia [1,22,23]. In this region, previous reports have revealed that some germplasm display an early-flowering phenotype characterized by short juvenility; such plants may produce flowers within three years (or even within the same year) of germination [24,25]. In these early-flowering germplasm, orthologs of the Arabidopsis MADS-box genes AGAMOUS and APETALA 3 may be responsible for the early transition from the juvenile to the adult phase [24]. In walnut germplasm from the Xinjiang Uygur Autonomous Region, near Central Asia, we here revealed that another MADS-box gene, FLC, was involved in this transition. However, the mechanism by which MADS-box genes regulate the floral transition in early-flowering walnuts remains unclear.
Notably, with the improvement in sequence technologies, we have previously performed RNA sequencing and microRNA sequencing for analyzing transcriptome-wide expression of genes and miRNAs in the hybrids between precocious walnut ‘Xinwen 81’ and a local later-flowering walnut germplasm [12]. In a previous study, we identified a large amount of differentially expressed genes (DEGs), including these MADS-box genes [12]. Guided by this work with a strategy for the molecular identification of a MADS-box gene FLC, our further study will isolate more MADS-box genes related to the regulation of floral transition in Xinjiang precocious walnuts.
In addition to MADS-box genes, flowering time in plants is also regulated by epigenetic mechanisms [26], including DNA methylation, non-coding RNAs, and histone modifications. These modifications control growth and development by regulating gene expression. In Brassicaceae species, the transcription factor FLC represses flowering until a plant is exposed to extended cold conditions (i.e., winter) in a process known as vernalization [10,27]. The prolonged chilling of winter leads to higher histone H3 lysine 27 trimethylation (H3K27me3) and lower histone H3 lysine 4 trimethylation (H3K4me3) levels at the FLC locus, stably repressing FLC transcription even after plants are returned to warmer temperatures [10,27]. It is therefore likely that temperature signals such as these contribute to the rapid floral transition among Xinjiang precocious walnuts under natural low-chilling, long-day conditions. Future studies should explore this dynamic epigenetic regulation of the FLC locus in walnut in depth.
4. Conclusions
This brief report summarizes our investigation into juvenility among two precocious varieties of Xinjiang walnut. We identified the key flowering-time gene FLC as a driver of precocity in these varieties. JrFLC had a high sequence identity with AtFLC and, like AtFLC, was shown to be nuclear-localized. In addition, JrFLC overexpression in Arabidopsis delayed flowering time. Together, the results of this study indicate that JrFLC played an important role in regulating the floral transition among Xinjiang precocious walnuts. This gene is therefore a strong candidate for future genetic improvement in flowering time in walnut.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9050582/s1, Table S1: Full-length FLC coding sequence and list of primers used in this study; Figure S1: Dynamic expression levels of JrFLC in ‘Xinwen 179’.
Author Contributions
Conceptualization, Q.J.; methodology, Q.J.; software, Q.J. and R.Z.; validation, Q.J. and L.C.; formal analysis, R.Z. and L.C.; investigation, Q.J.; resources, Q.J. and R.Z.; writing—original draft preparation, Q.J.; writing—review and editing, Z.L.; visualization, L.C.; supervision, L.C. and Z.L.; project administration, L.C. and Z.L.; funding acquisition, L.C. and Q.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Major Scientific and Technological Projects of XPCC (2017DB006), the National Natural Science Foundation of China (31260463, 32160698), and the Agricultural AI Application and Innovation Team of Tarim University (TDZKCX202308).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bernard, A.; Lheureux, F.; Dirlewanger, E. Walnut: Past and future of genetic improvement. Tree Genet. Genomes 2018, 14, 1–28. [Google Scholar] [CrossRef]
- Jahanban-Esfahlan, A.; Ostadrahimi, A.; Tabibiazar, M.; Amarowicz, R.; Tabibiazar, M.; Amarowicz, R. A comprehensive review on the chemical constituents and functional uses of walnut (Juglans spp.) husk. Int. J. Mol. Sci. 2019, 20, 3920. [Google Scholar] [CrossRef]
- Sheng, F.; Hu, B.; Jin, Q.; Wang, J.; Wu, C.; Luo, Z. The analysis of phenolic compounds in walnut husk and pellicle by UPLC-Q-Orbitrap HRMS and HPLC. Molecules 2021, 26, 3013. [Google Scholar] [CrossRef]
- Hardman, W.E. Walnuts have potential for cancer prevention and treatment in mice. J. Nutr. 2014, 144, 555S–560S. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Trandafir, I.; Cosmulescu, S.; Nour, V. Phenolic profile and antioxidant capacity of walnut extract as influenced by the extraction method and solvent. Int. J. Food Eng. 2017, 13, 1–8. [Google Scholar] [CrossRef]
- Gu, X.; Wang, Y.; He, Y. Photoperiodic regulation of flowering time through periodic histone deacetylation of the florigen gene FT. PLoS Biol. 2013, 11, e1001649. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Jiang, D.; Wang, Y.; Bachmair, A.; He, Y. Repression of the floral transition via histone H2B monoubiquitination. Plant J. 2009, 57, 522–533. [Google Scholar] [CrossRef]
- Jiang, D.; Gu, X.; He, Y. Establishment of the winter-annual growth habit via FRIGIDA-mediated histone methylation at FLOWERING LOCUS C in Arabidopsis. Plant Cell 2009, 21, 1733–1746. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Amasino, R.M. Role of chromatin modification in flowering-time control. Trends Plant Sci. 2005, 10, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.A.; Zhang, Q.; Huang, M.M.; Yakup, A. The breeding of six Xinjiang dwarf walnut cultivars. Acta Hortic. 2014, 1050, 151–160. [Google Scholar] [CrossRef]
- Jin, Q.; Mo, R.; Chen, W.; Zhang, Q.; Sheng, F.; Wu, C.; Zhang, R.; Luo, Z. Identification and comparative analysis of genes and microRNAs involved in the floral transition of the Xinjiang early-flowering walnut (Juglans regia L.). Horticulturae 2022, 8, 136. [Google Scholar] [CrossRef]
- Yuan, X.; Niu, J.; Zhou, L.; Quan, S.; Ma, L.; Xu, H. Evolutionary analysis of MIR156/157 family during floral induction in walnut (Juglans regia L.). J. Hortic. Sci. Biotechnol. 2022, 97, 697–707. [Google Scholar] [CrossRef]
- Zhu, T.; Wang, L.; You, F.M.; Rodriguez, J.C.; Deal, K.R.; Chen, L.; Li, J.; Chakraborty, S.; Balan, B.; Jiang, C.Z.; et al. Sequencing a Juglans regia × J. microcarpa hybrid yields high-quality genome assemblies of parental species. Hortic. Res. 2019, 6, 55. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Zhang, P.; Zhu, C.; Geng, Y.; Wang, Y.; Yang, Y.; Liu, Q.; Guo, W.; Chachar, S.; Riaz, A.; Yan, S.; et al. Rice and Arabidopsis homologs of yeast chromosome transmission fidelity protein 4 commonly interact with Polycomb complexes but exert divergent regulatory functions. Plant Cell 2021, 33, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.; Wang, Y.; Guo, W.; Chachar, S.; Riaz, A.; Geng, Y.; Gu, X.; Yang, L. PRMT6 physically associates with nuclear factor Y to regulate photoperiodic flowering in Arabidopsis. aBIOTECH 2021, 2, 403–414. [Google Scholar] [CrossRef]
- Jiang, D.; Yang, W.; He, Y.; Amasino, R.M. Arabidopsis relatives of the human lysine-specific demethylase1 repress the expression of FWA and FLOWERING LOCUS C and thus promote the floral transition. Plant Cell 2007, 19, 2975–2987. [Google Scholar] [CrossRef]
- Chen, W.; Tamada, Y.; Yamane, H.; Matsushita, M.; Osako, Y.; Gao-takai, M. H3K4me3 plays a key role in establishing permissive chromatin states during bud dormancy and bud break in apple. Plant J. 2022, 111, 1015–1031. [Google Scholar] [CrossRef] [PubMed]
- Hassankhah, A.; Rahemi, M.; Ramshini, H.; Sarikhani, S.; Vahdati, K. Flowering in Persian walnut: Patterns of gene expression during flower development. BMC Plant Biol. 2020, 20, 136. [Google Scholar] [CrossRef] [PubMed]
- Vahdati, K.; Mohseniazar, M. Early bearing genotypes of walnut: A suitable material for breeding and high density orchards. Acta Hortic. 2016, 1139, 101–105. [Google Scholar] [CrossRef]
- Breton, C.; Cornu, D.; Chriqui, D.; Sauvanet, A.; Capelli, P.; Germain, E.; Jay-Allemand, C. Somatic embryogenesis, micropropagation and plant regeneration of “Early Mature” walnut trees (Juglans regia) that flower in vitro. Tree Physiol. 2004, 24, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, A.; Zarei, A.; Zamani Fardadonbeh, M.; Lawson, S. Evaluation of genetic variability among “Early Mature” Juglans regia using microsatellite markers and morphological traits. PeerJ 2017, 5, e3834. [Google Scholar] [CrossRef]
- He, Y. Chromatin regulation of flowering. Trends Plant Sci. 2012, 17, 556–562. [Google Scholar] [CrossRef]
- He, Y.; Li, Z. Epigenetic environmental memories in plants: Establishment, maintenance, and reprogramming. Trends Genet. 2018, 34, 856–866. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).