Enrichment of Rabbit Primitive Hematopoietic Cells via MACS Depletion of CD45+ Bone Marrow Cells

 , ,
, ,
Abstract
:1. Introduction
2. Results
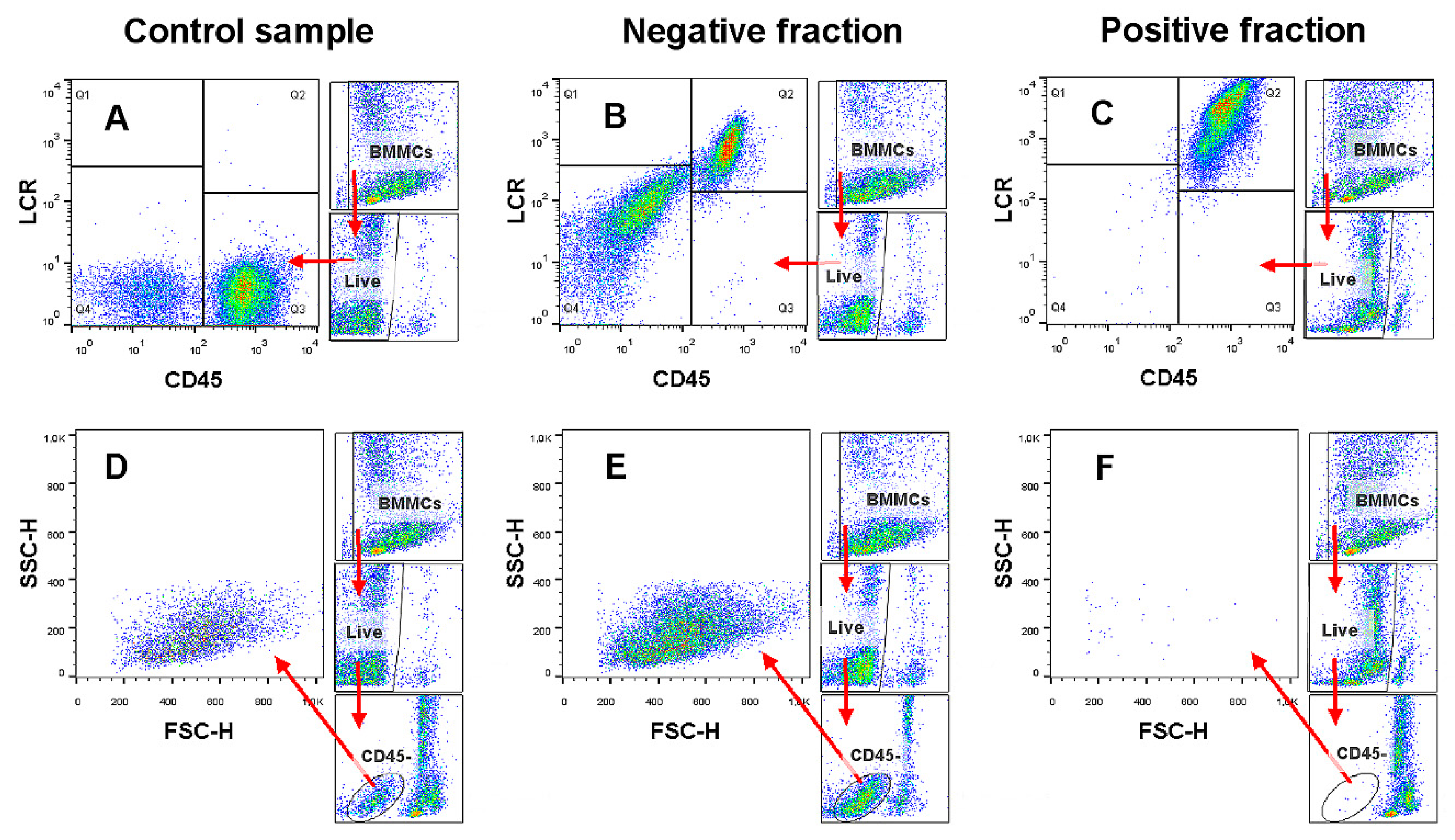
2.1. Magnetic-Activated Cell Sorting (MACS) Depletion of CD45+ Cells
2.2. Expansion of CD45 Depleted Cells
2.2.1. Cell Proliferation
2.2.2. Flow-Cytometric Analysis of the CD45− Cultured Cells
2.2.3. Real-Time Polymerase Chain Reaction (PCR) Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. MACS Sorting Efficiency and Sample Purity
4.4. Expansion of CD45 Depleted Rabbit Hematopoietic Cells
4.5. Phenotypic Analysis of the CD45− Cultured Cells
4.6. Real-Time PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hindorf, C.; Glatting, G.; Chiesa, C.; Linden, O.; Flux, G.; Committee, E.D. EANM Dosimetry Committee guidelines for bone marrow and whole-body dosimetry. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 1238–1250. [Google Scholar] [CrossRef]
- Greer, J.P.; Arber, D.A.; Glader, B.E.; List, A.F.; Means, R.M.; Rodgers, G.M. Wintrobe’s Clinical Hematology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2018; p. 2346. [Google Scholar]
- Zhao, E.; Xu, H.; Wang, L.; Kryczek, I.; Wu, K.; Hu, Y.; Wang, G.; Zou, W. Bone marrow and the control of immunity. Cell. Mol. Immunol. 2012, 9, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tencerova, M.; Kassem, M. The Bone Marrow-Derived Stromal Cells: Commitment and Regulation of Adipogenesis. Front. Endocrinol. 2016, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- Weissman, I.L. Stem cells: Units of development, units of regeneration, and units in evolution. Cell 2000, 100, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Kondo, M.; Wagers, A.J.; Manz, M.G.; Prohaska, S.S.; Scherer, D.C.; Beilhack, G.F.; Shizuru, J.A.; Weissman, I.L. Biology of hematopoietic stem cells and progenitors: Implications for clinical application. Annu. Rev. Immunol. 2003, 21, 759–806. [Google Scholar] [CrossRef] [PubMed]
- Bryder, D.; Rossi, D.J.; Weissman, I.L. Hematopoietic stem cells: The paradigmatic tissue-specific stem cell. Am. J. Pathol. 2006, 169, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, H.; Katsura, Y. A new paradigm for hematopoietic cell lineages: Revision of the classical concept of the myeloid-lymphoid dichotomy. Trends Immunol. 2009, 30, 193–200. [Google Scholar] [CrossRef]
- Watt, S.M.; Austin, E.; Armitage, S. Cryopreservation of hematopoietic stem/progenitor cells for therapeutic use. Methods Mol. Biol. 2007, 368, 237–259. [Google Scholar] [CrossRef]
- Bosio, A.; Huppert, V.; Donath, S.; Hennemann, P.; Malchow, M.; Heinlein, U.A. Isolation and enrichment of stem cells. Adv. Biochem. Eng. Biotechnol. 2009, 114, 23–72. [Google Scholar] [CrossRef]
- Porada, C.D.; Harrison-Findik, D.D.; Sanada, C.; Valiente, V.; Thain, D.; Simmons, P.J.; Almeida-Porada, G.; Zanjani, E.D. Development and characterization of a novel CD34 monoclonal antibody that identifies sheep hematopoietic stem/progenitor cells. Exp. Hematol. 2008, 36, 1739–1749. [Google Scholar] [CrossRef] [Green Version]
- Vašíček, J.; Shehata, M.; Schnabl, S.; Hilgarth, M.; Hubmann, R.; Jäger, U.; Bauer, M.; Chrenek, P. Critical assessment of the efficiency of CD34 and CD133 antibodies for enrichment of rabbit hematopoietic stem cells. Biotechnol. Prog. 2018, 34, 1278–1289. [Google Scholar] [CrossRef]
- Goodell, M.A.; Brose, K.; Paradis, G.; Conner, A.S.; Mulligan, R.C. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. J. Exp. Med. 1996, 183, 1797–1806. [Google Scholar] [CrossRef] [Green Version]
- Emery, D.W.; Sablinski, T.; Arn, J.S.; LeGuern, C.; Sachs, D.H. Bone marrow culture and transduction of stem cells in a miniature swine model. Blood Cells 1994, 20, 498–502. [Google Scholar]
- Jeong, D.K.; Sharma, N.; Kim, J.N.; Sodhi, S.; Oh, S. Optimization of Techniques for Isolation and Identification of Bovine Hematopoietic Stem Cells from Peripheral Blood. Reprod. Dev. Biol. Suppl. 2012, 36, 138. [Google Scholar]
- Oppenheim, S.M.; Moyer, A.L.; BonDurant, R.H.; Rowe, J.D.; Anderson, G.B. Evidence against humoral immune attack as the cause of sheep-goat interspecies and hybrid pregnancy failure in the doe. Theriogenology 2001, 55, 1567–1581. [Google Scholar] [CrossRef]
- Fang, J.; Jensen, E.S.; Boudreaux, M.K.; Du, L.M.; Hawkins, T.B.; Koukouritaki, S.B.; Cornetta, K.; Wilcox, D.A. Platelet gene therapy improves hemostatic function for integrin alphaIIbbeta3-deficient dogs. Proc. Natl. Acad. Sci. USA 2011, 108, 9583–9588. [Google Scholar] [CrossRef] [Green Version]
- Houdebine, L.M.; Fan, J. Rabbit Biotechnology: Rabbit Genomics, Transgenesis, Cloning and Models; Springer: Cham, The Netherlands, 2009; p. 136. [Google Scholar] [CrossRef]
- Carneiro, M.; Afonso, S.; Geraldes, A.; Garreau, H.; Bolet, G.; Boucher, S.; Tircazes, A.; Queney, G.; Nachman, M.W.; Ferrand, N. The genetic structure of domestic rabbits. Mol. Biol. Evol. 2011, 28, 1801–1816. [Google Scholar] [CrossRef] [Green Version]
- Hasenberg, M.; Kohler, A.; Bonifatius, S.; Borucki, K.; Riek-Burchardt, M.; Achilles, J.; Mann, L.; Baumgart, K.; Schraven, B.; Gunzer, M. Rapid immunomagnetic negative enrichment of neutrophil granulocytes from murine bone marrow for functional studies in vitro and in vivo. PLoS ONE 2011, 6, e17314. [Google Scholar] [CrossRef]
- Gallacher, L.; Murdoch, B.; Wu, D.M.; Karanu, F.N.; Keeney, M.; Bhatia, M. Isolation and characterization of human CD34(-)Lin(-) and CD34(+)Lin(-) hematopoietic stem cells using cell surface markers AC133 and CD7. Blood 2000, 95, 2813–2820. [Google Scholar] [CrossRef]
- Vašíček, J.; Baláži, A.; Parkányi, V.; Bauer, M. Different MACS sorting strategies for the enrichment of Lin-(CD34+ CD45-) hematopoietic progenitor cells: Preliminary study. Ann. Univ. Paedagog. Crac. Stud. Nat. 2018, 3, 83–89. [Google Scholar] [CrossRef]
- Gunzer, M.; Weishaupt, C.; Planelles, L.; Grabbe, S. Two-step negative enrichment of CD4+ and CD8+ T cells from murine spleen via nylon wool adherence and an optimized antibody cocktail. J. Immunol. Methods 2001, 258, 55–63. [Google Scholar] [CrossRef]
- Cheng, P.; Zlobin, A.; Volgina, V.; Gottipati, S.; Osborne, B.; Simel, E.J.; Miele, L.; Gabrilovich, D.I. Notch-1 regulates NF-kappaB activity in hemopoietic progenitor cells. J. Immunol. 2001, 167, 4458–4467. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.; Corzo, C.A.; Luetteke, N.; Yu, B.; Nagaraj, S.; Bui, M.M.; Ortiz, M.; Nacken, W.; Sorg, C.; Vogl, T.; et al. Inhibition of dendritic cell differentiation and accumulation of myeloid-derived suppressor cells in cancer is regulated by S100A9 protein. J. Exp. Med. 2008, 205, 2235–2249. [Google Scholar] [CrossRef]
- Ema, H.; Morita, Y.; Yamazaki, S.; Matsubara, A.; Seita, J.; Tadokoro, Y.; Kondo, H.; Takano, H.; Nakauchi, H. Adult mouse hematopoietic stem cells: Purification and single-cell assays. Nat. Protoc. 2006, 1, 2979–2987. [Google Scholar] [CrossRef]
- Nakano, A.; Harada, T.; Morikawa, S.; Kato, Y. Expression of leukocyte common antigen (CD45) on various human leukemia/lymphoma cell lines. Acta Pathol. Jpn. 1990, 40, 107–115. [Google Scholar] [CrossRef]
- Wu, X.; Dao Thi, V.L.; Huang, Y.; Billerbeck, E.; Saha, D.; Hoffmann, H.H.; Wang, Y.; Silva, L.A.V.; Sarbanes, S.; Sun, T.; et al. Intrinsic Immunity Shapes Viral Resistance of Stem Cells. Cell 2018, 172, 423–438.e25. [Google Scholar] [CrossRef]
- Agudelo, D.; Duringer, A.; Bozoyan, L.; Huard, C.C.; Carter, S.; Loehr, J.; Synodinou, D.; Drouin, M.; Salsman, J.; Dellaire, G.; et al. Marker-free coselection for CRISPR-driven genome editing in human cells. Nat. Methods 2017, 14, 615–620. [Google Scholar] [CrossRef]
- StemSpan™ CD34+ Expansion Supplement (10X). Available online: https://www.stemcell.com/stemspan-cd34-expansion-supplement-10x.html (accessed on 27 November 2020).
- Baum, C.M.; Weissman, I.L.; Tsukamoto, A.S.; Buckle, A.M.; Peault, B. Isolation of a candidate human hematopoietic stem-cell population. Proc. Natl. Acad. Sci. USA 1992, 89, 2804–2808. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, M.; Wang, J.C.; Kapp, U.; Bonnet, D.; Dick, J.E. Purification of primitive human hematopoietic cells capable of repopulating immune-deficient mice. Proc. Natl. Acad. Sci. USA 1997, 94, 5320–5325. [Google Scholar] [CrossRef] [Green Version]
- Majeti, R.; Park, C.Y.; Weissman, I.L. Identification of a hierarchy of multipotent hematopoietic progenitors in human cord blood. Cell Stem Cell 2007, 1, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of single human hematopoietic stem cells capable of long-term multilineage engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef]
- Rodin, S.; Antonsson, L.; Niaudet, C.; Simonson, O.E.; Salmela, E.; Hansson, E.M.; Domogatskaya, A.; Xiao, Z.; Damdimopoulou, P.; Sheikhi, M.; et al. Clonal culturing of human embryonic stem cells on laminin-521/E-cadherin matrix in defined and xeno-free environment. Nat. Commun. 2014, 5, 3195. [Google Scholar] [CrossRef]
- Yang, Z.; Dong, P.; Fu, X.; Li, Q.; Ma, S.; Wu, D.; Kang, N.; Liu, X.; Yan, L.; Xiao, R. CD49f Acts as an Inflammation Sensor to Regulate Differentiation, Adhesion, and Migration of Human Mesenchymal Stem Cells. Stem Cells 2015, 33, 2798–2810. [Google Scholar] [CrossRef]
- Krebsbach, P.H.; Villa-Diaz, L.G. The Role of Integrin α6 (CD49f) in Stem Cells: More than a Conserved Biomarker. Stem Cells Dev. 2017, 26, 1090–1099. [Google Scholar] [CrossRef]
- Koutna, I.; Peterkova, M.; Simara, P.; Stejskal, S.; Tesarova, L.; Kozubek, M. Proliferation and differentiation potential of CD133+ and CD34+ populations from the bone marrow and mobilized peripheral blood. Ann. Hematol 2011, 90, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Layton, D.S.; Strom, A.D.; O‘Neil, T.E.; Broadway, M.M.; Stephenson, G.L.; Morris, K.R.; Muralitharan, M.; Sandrin, M.S.; Ierino, F.L.; Bean, A.G. Development of an anti-porcine CD34 monoclonal antibody that identifies hematopoietic stem cells. Exp. Hematol. 2007, 35, 171–178. [Google Scholar] [CrossRef]
- Le Guern, A.C.; Giovino, M.A.; Abe, M.; Theodore, P.R.; Qi, J.; Down, J.D.; Sachs, D.H.; Sykes, M.; Yang, Y.G. Stem cell activity of porcine c-kit+ hematopoietic cells. Exp. Hematol. 2003, 31, 833–840. [Google Scholar] [CrossRef]
- Oka, M.; Tagoku, K.; Russell, T.L.; Nakano, Y.; Hamazaki, T.; Meyer, E.M.; Yokota, T.; Terada, N. CD9 is associated with leukemia inhibitory factor-mediated maintenance of embryonic stem cells. Mol. Biol. Cell 2002, 13, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Briddell, R.A.; Broudy, V.C.; Bruno, E.; Brandt, J.E.; Srour, E.F.; Hoffman, R. Further phenotypic characterization and isolation of human hematopoietic progenitor cells using a monoclonal antibody to the c-kit receptor. Blood 1992, 79, 3159–3167. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, J.; Machalinski, B.; Majka, M.; Kijowski, J.; Marlicz, W.; Rozmyslowicz, T.; Ostrowski, M.; Ratajczak, M.Z. Evidence that human haematopoietic stem cells (HSC) do not reside within the CD34+KIT- cell population. Ann. Transplant. 1999, 4, 22–30. [Google Scholar]
- Tsai, F.Y.; Keller, G.; Kuo, F.C.; Weiss, M.; Chen, J.; Rosenblatt, M.; Alt, F.W.; Orkin, S.H. An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature 1994, 371, 221–226. [Google Scholar] [CrossRef]
- Briegel, K.; Lim, K.C.; Plank, C.; Beug, H.; Engel, J.D.; Zenke, M. Ectopic expression of a conditional GATA-2/estrogen receptor chimera arrests erythroid differentiation in a hormone-dependent manner. Genes Dev. 1993, 7, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Akashi, K.; Traver, D.; Miyamoto, T.; Weissman, I.L. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 2000, 404, 193–197. [Google Scholar] [CrossRef]
- Ku, C.J.; Hosoya, T.; Maillard, I.; Engel, J.D. GATA-3 regulates hematopoietic stem cell maintenance and cell-cycle entry. Blood 2012, 119, 2242–2251. [Google Scholar] [CrossRef] [Green Version]
- Darr, H.; Benvenisty, N. Factors involved in self-renewal and pluripotency of embryonic stem cells. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–19. [Google Scholar]
- He, S.; Kim, I.; Lim, M.S.; Morrison, S.J. Sox17 expression confers self-renewal potential and fetal stem cell characteristics upon adult hematopoietic progenitors. Genes Dev. 2011, 25, 1613–1627. [Google Scholar] [CrossRef] [Green Version]
- Kovac, M.; Vasicek, J.; Kulikova, B.; Bauer, M.; Curlej, J.; Balazi, A.; Chrenek, P. Different RNA and protein expression of surface markers in rabbit amniotic fluid-derived mesenchymal stem cells. Biotechnol. Prog. 2017, 33, 1601–1613. [Google Scholar] [CrossRef]
- Vašíček, J.; Kováč, M.; Baláži, A.; Kulíková, B.; Tomková, M.; Olexiková, L.; Čurlej, J.; Bauer, M.; Schnabl, S.; Hilgarth, M.; et al. Combined approach for characterization and quality assessment of rabbit bone marrow-derived mesenchymal stem cells intended for gene banking. New Biotechnol. 2020, 54, 1–12. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Kulikova, B.; Kovac, M.; Bauer, M.; Tomkova, M.; Olexikova, L.; Vasicek, J.; Balazi, A.; Makarevich, A.V.; Chrenek, P. Survivability of rabbit amniotic fluid-derived mesenchymal stem cells post slow-freezing or vitrification. Acta Histochem. 2019, 121, 491–499. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucl. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Host/Isotype | Clone | Conjugate | Company |
|---|---|---|---|---|
| CD45 | mouse IgG2a | ISC18A | purified 1 | WSU |
| MHCI | mouse IgG2a | H58A | purified 1 | WSU |
| MHCII | mouse IgG2a | TH14B | purified 1 | WSU |
| CD38 | mouse IgG1 | 01 | purified 1 | Sino Biological |
| CD49f | rat IgG2a | GoH3 | AF647 | BioLegend |
| CD90 | mouse IgG1 | 5E10 | FITC | BD Biosciences |
| c-kit (CD117) | rabbit IgG | polyclonal | FITC | Biorbyt |
| Gene | Product Size (bp) | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|---|
| CD9 | 158 | 5′-CAACAAATTCCACGTCATCG-3′ | 5′-TTGAGGGGTACACCTTCCTG-3′ | XM_017343606.1 1 |
| CD34 | 155 | 5′-CTGAGGTTAGGGCTCAGTGC-3′ | 5′-GGAGTAGCTCTGGTGGCTTG-3′ | [22] |
| CD38 | 148 | 5′-CAACCTTGTGTGGTGTGGAG-3′ | 5′-GACCGTACCACAGGCAGATT-3′ | NM_001082683.1 1 |
| CD45 | 262 | 5′-TACTCTGCCTCCCGTTG-3′ | 5′-GCTGAGTGTCTGCGTGTC-3′ | [51] |
| CD49f | 123 | 5′-AGGTACAGTGGTCGGTGAGC-3′ | 5′-TTCAAAGTTGCTGTGCCAAG-3′ | XM_017342931.1 1 |
| CD90 | 293 | 5′-CTGCTGCTGCTCTCACTGTC-3′ | 5′-ACAGAAGCAGCTTTGGGAAA-3′ | [51] |
| CD172a | 162 | 5′-CCAGCATTCAGATTGGTCCT-3′ | 5′-GTCCCATTCACTTCCTCTGC-3′ | XM_017341726.1 1 |
| CDH1 (E-cadherin) | 168 | 5′-GAATCCTGGCTCTGCTCATC-3′ | 5′-GCTGGCTCAAGTCAAAGTCC-3′ | XM_002711639.3 1 |
| c-kit (CD117) | 189 | 5′-CCGGTGGACTCCAAGTTCTA-3′ | 5′-GTAAACGTGGTGGGTGCTCT-3′ | NM_001329070.1 1 |
| MHCI | 97 | 5′-AGTGGGAATTGTTGCTGGAG-3′ | 5′-TCCTTTCCCATCTGAGCTGT-3′ | NM_001171270.2 1 |
| MHCII | 142 | 5′-CTGTGAGCACAGGAAGGTGA-3′ | 5′-GGGTGGCTAGATGTCTGGAA-3′ | NM_001171118.1 1 |
| GATA1 | 164 | 5′-AAACGGGCAGGTACTCAGTG-3′ | 5′-TTCCGAGTCTGGATTCCATC-3′ | XM_002719899.3 1 |
| GATA2 | 137 | 5′-GACAAGGACGGGGTCAAGTA-3′ | 5′-TAGGAGGGGTAGGTGGGAAT-3′ | XM_008260327.2 1 |
| GATA3 | 118 | 5′-AAGGCAGGGAGTGTGTGAAC-3′ | 5′-CGGTTCTGTCCGTTCATTTT-3′ | XM_002717361.3 1 |
| NANOG | 122 | 5′-GCCAGTCGTGGAGTAACCAT-3′ | 5′-CTGCATGGAGGACTGTAGCA-3′ | [51] |
| OCT4 | 149 | 5′-GAGGAGTCCCAGGACATGAA-3′ | 5′-GTGGTTTGGCTGAACACCTT-3′ | [51] |
| SOX2 | 152 | 5′-CAGCTCGCAGACCTACATGA-3′ | 5′-TGGAGTGGGAGGAAGAGGTA-3′ | [51] |
| SOX17 | 135 | 5′-CGTGTTGTTGCGTTTTGTTT-3′ | 5′-CGGGGAACAAGTCTCTGAAG-3′ | XM_008255533.2 1 |
| B2M | 118 | 5′-ATTCACGCCCAATGATAAGG-3′ | 5′-ATCCTCAGACCTCCATGCTG-3′ | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vašíček, J.; Baláži, A.; Bauer, M.; Svoradová, A.; Tirpáková, M.; Ondruška, Ľ.; Parkányi, V.; V. Makarevich, A.; Chrenek, P. Enrichment of Rabbit Primitive Hematopoietic Cells via MACS Depletion of CD45+ Bone Marrow Cells. Magnetochemistry 2021, 7, 11. https://doi.org/10.3390/magnetochemistry7010011
Vašíček J, Baláži A, Bauer M, Svoradová A, Tirpáková M, Ondruška Ľ, Parkányi V, V. Makarevich A, Chrenek P. Enrichment of Rabbit Primitive Hematopoietic Cells via MACS Depletion of CD45+ Bone Marrow Cells. Magnetochemistry. 2021; 7(1):11. https://doi.org/10.3390/magnetochemistry7010011
Chicago/Turabian StyleVašíček, Jaromír, Andrej Baláži, Miroslav Bauer, Andrea Svoradová, Mária Tirpáková, Ľubomír Ondruška, Vladimír Parkányi, Alexander V. Makarevich, and Peter Chrenek. 2021. "Enrichment of Rabbit Primitive Hematopoietic Cells via MACS Depletion of CD45+ Bone Marrow Cells" Magnetochemistry 7, no. 1: 11. https://doi.org/10.3390/magnetochemistry7010011
APA StyleVašíček, J., Baláži, A., Bauer, M., Svoradová, A., Tirpáková, M., Ondruška, Ľ., Parkányi, V., V. Makarevich, A., & Chrenek, P. (2021). Enrichment of Rabbit Primitive Hematopoietic Cells via MACS Depletion of CD45+ Bone Marrow Cells. Magnetochemistry, 7(1), 11. https://doi.org/10.3390/magnetochemistry7010011