
Differential Post-Fire Recovery of Tree and Shrub Growth and Water-Use Efficiency in a Mediterranean Coastal Dune System

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
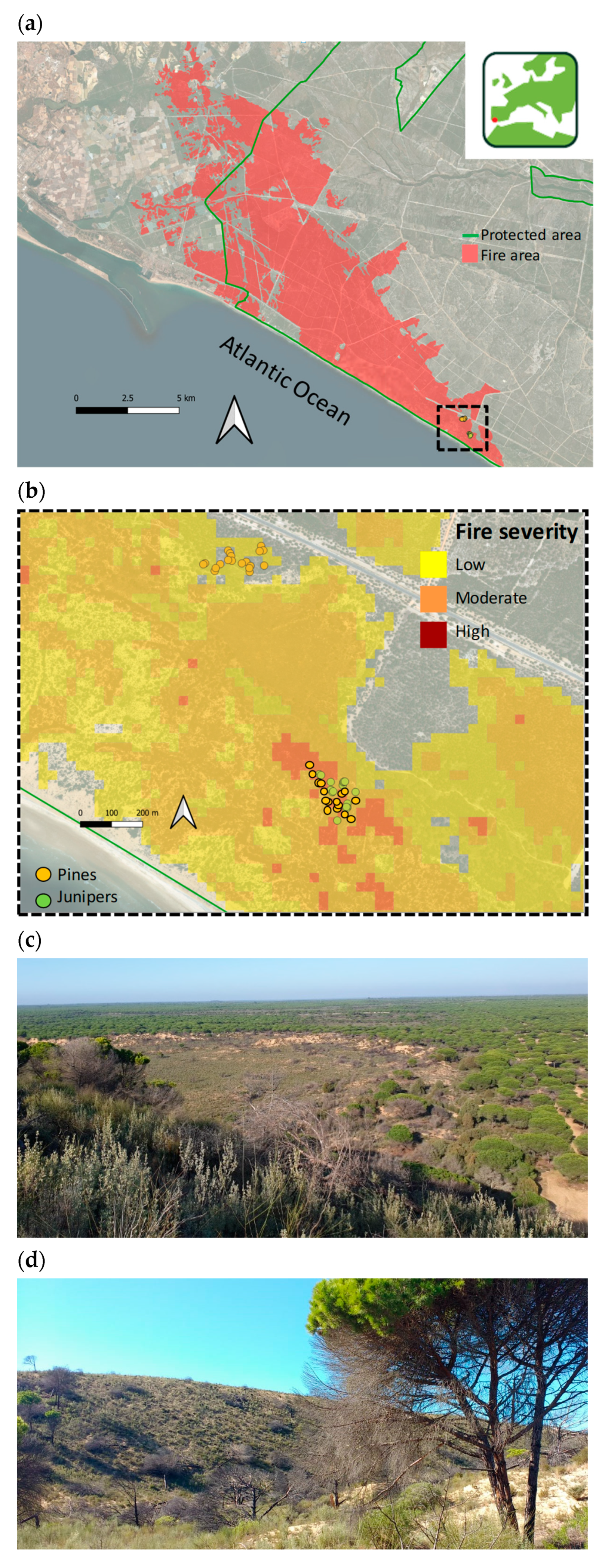
2.1. Study Area and Wildfire Features
2.2. Field Sampling
2.3. Processing Wood Samples and Tree-Ring Width Data
2.4. Calculating Intrinsic Water-Use Efficiency (WUEi)
2.5. Statistical Analyses
3. Results
3.1. Fire Impacts on Dune and Inter-Dune Pines and Junipers: Crown Damage and Bark Char
3.2. Fire Impacts on Growth in Dune and Slack Pines and Junipers
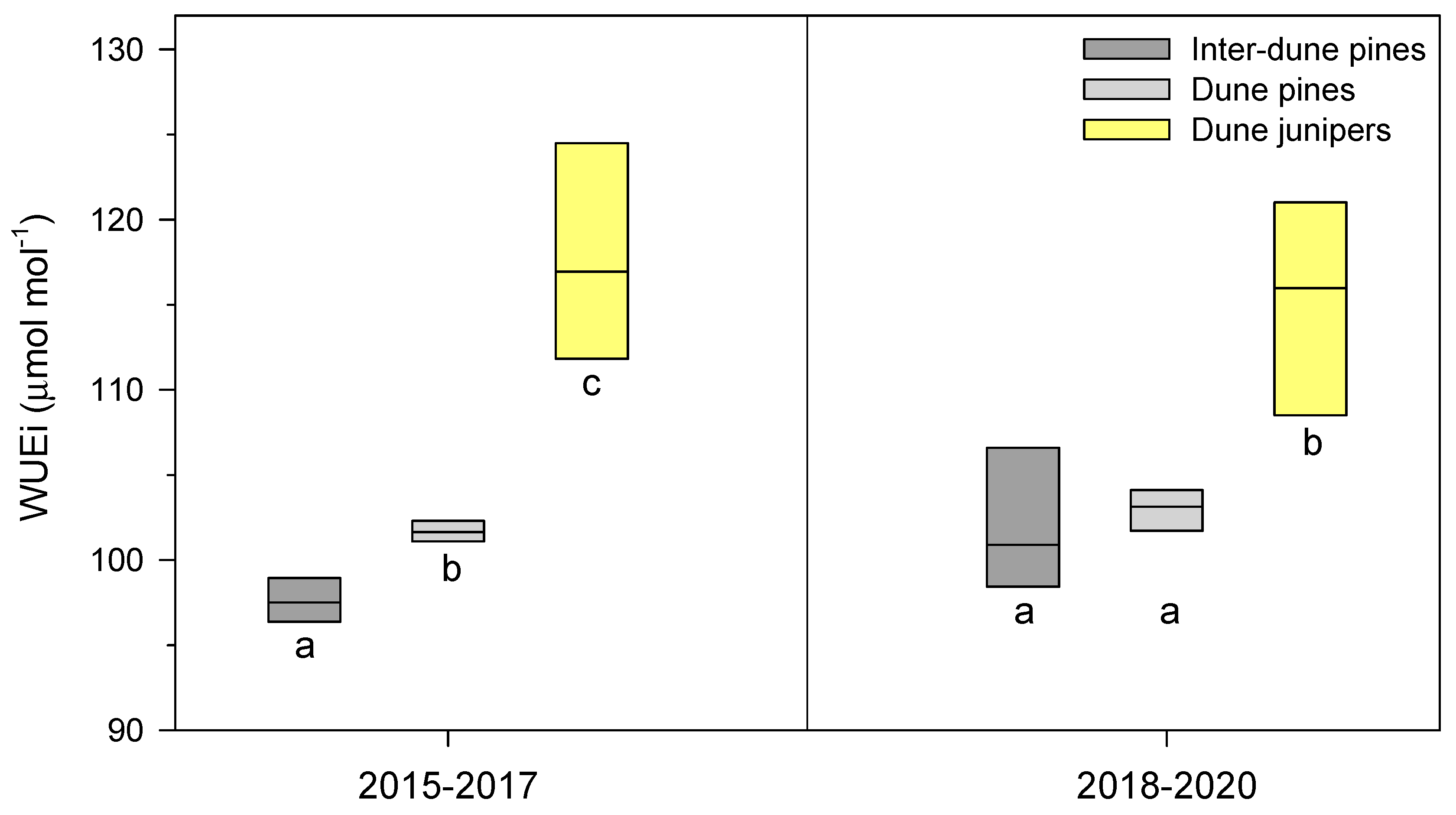
3.3. Fire Impacts on Water-Use Efficiency in Dune and Inter-Dune Pines and Junipers
4. Discussion
4.1. Fire Impacts on Pine and Juniper Growth Depend on Site Conditions
4.2. Fire Impacts on Water-Use Efficiency: Converging Values in Dune and Inter-Dune Pines
4.3. Management Implications and Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC) IPCC. Summary for Policymakers. In Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2022; pp. 1–35. [Google Scholar]
- Turco, M.; Rosa-Cánovas, J.J.; Bedia, J.; Jerez, S.; Montávez, J.P.; Llasat, M.C.; Provenzale, A. Exacerbated fires in Mediterranean Europe due to anthropogenic warming projected with non-stationary climate-fire models. Nat. Commun. 2018, 9, 3821. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the earth system. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Batllori, E.; Lloret, F.; Aakala, T.; Anderegg, W.R.L.; Aynekulu, E.; Bendixsen, D.P.; Bentouati, A.; Bigler, C.; Burk, C.J.; Camarero, J.J.; et al. Forest and woodland replacement patterns following drought-related mortality. Proc. Natl. Acad. Sci. USA 2020, 117, 29720–29729. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Verdú, M. Plant persistence traits in fire-prone ecosystems of the Mediterranean basin: A phylogenetic approach. Oikos 2005, 109, 196–202. [Google Scholar] [CrossRef]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of south-eastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Quevedo, L.; Rodrigo, A.; Espelta, J.M. Post-fire resprouting ability of 15 non-dominant shrub and tree species in Mediterranean areas of NE Spain. Ann. For. Sci. 2007, 64, 883–890. [Google Scholar] [CrossRef]
- Bär, A.; Michaletz, S.T.; Mayr, S. Fire effects on tree physiology. New Phytol. 2019, 223, 1728–1741. [Google Scholar] [CrossRef]
- Zunzunegui, M.; Díaz Barradas, M.C.; Ain-Lhout, F.; Alvarez-Cansino, L.; Esquivias, M.P.; García Novo, F. Seasonal physiological plasticity and recovery capacity after summer stress in Mediterranean scrub communities. Plant Ecol. 2011, 212, 127–142. [Google Scholar] [CrossRef]
- Armas, C.; Padilla, F.M.; Pugnaire, F.I.; Jackson, R.B. Hydraulic lift and tolerance to salinity of semiarid species, consequences for species interactions. Oecologia 2010, 162, 11–21. [Google Scholar] [CrossRef]
- Lloret, F.; Granzow-de la Cerda, I. Plant competition and facilitation after extreme drought episodes in Mediterranean shrub-land, does damage to vegetation cover trigger replacement by juniper woodland? J. Veg. Sci. 2013, 24, 1020–1032. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sánchez-Salguero, R.; Sangüesa-Barreda, G.; Díaz-Delgado, R.; Casals, R. Dieback and mortality of junipers caused by drought: Dissimilar growth and wood isotope patterns preceding shrub death. Agric. For. Meteorol. 2020, 91, 108078. [Google Scholar] [CrossRef]
- Baquedano, F.J.; Castillo, F.J. Drought tolerance in the Mediterranean species Quercus coccifera, Quercus ilex, Pinus halepensis and Juniperus phoenicea. Photosynthetica 2007, 45, 229–238. [Google Scholar] [CrossRef]
- Altieri, S.; Mereu, S.; Cherubini, P.; Castaldi, S.; Sirignano, C.; Lubritto, C.; Battipaglia, G. Tree-ring carbon and oxygen isotopes indicate different water use strategies in three Mediterranean shrubs at Capo Caccia (Sardinia, Italy). Trees 2015, 29, 1593–1603. [Google Scholar] [CrossRef]
- García Murillo, P. El incendio de las Peñuelas de 2017. In Ecología de la Regeneración de Zonas Incendiadas; García Novo, F., Casal, M., Pausas, J.G., Eds.; Junta de Andalucía: Sevilla, Spain, 2018; pp. 175–195. [Google Scholar]
- Informe Ejecutivo del Incendio de ‘Las Peñuelas’ Moguer (Huelva). Consejería de Medio Ambiente y Ordenación del Territorio. Junta de Andalucía, Sevilla, Spain. 2017. Available online: https://www.juntadeandalucia.es/medioambiente/portal_web/web/temas_ambientales/incendios_forestales/incendio_de_moguer/informe_ejecutivo_penuelas.pdf (accessed on 21 July 2022).
- García Murillo, P.; Sousa, A. El paisaje vegetal de la zona oeste del Parque Natural de Doñana (Huelva). Lagascalia 1999, 21, 111–131. [Google Scholar]
- Prada, A.; Gordo, J.; De Miguel, J.; Mutke, S.; Catalán, G.; Iglesias, S.; Gil, L. Regiones de Procedencia de Pinus pinea L. en España; O.A. Parques Nacionales: Madrid, Spain, 1997. [Google Scholar]
- Tapias, R.; Climent, J.; Pardos, J.A.; Gil, L. Life histories of Mediterranean pines. Plant Ecol. 2004, 171, 53–68. [Google Scholar] [CrossRef]
- Granados, M.; Martín, A.; García Novo, F. Long-term vegetation changes on the stabilized dunes of Doñana National Park (SW Spain). Vegetatio 1988, 75, 73–80. [Google Scholar]
- Ojeda Ribera, F. Políticas forestales y medio ambiente en Doñana y su entorno. Agric. Soc. 1992, 11, 303–357. [Google Scholar]
- García García, F. Doñana en su historia. In Organismo Autónomo de Parques Nacionales; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2016. [Google Scholar]
- Fernández, S.; Carrión, J.S.; Ochando, J.; González-Sampériz, P.; Munuera, M.; Munuera, M.; Amorós, G.; Postigo-Mijarra, J.M.; Morales-Molino, C.; García-Murillo, P.; et al. New palynological data from the Late Pleistocene glacial refugium of South-West Iberia: The case of Doñana. Rev. Palaeobot. Palynol. 2021, 290, 104431. [Google Scholar] [CrossRef]
- Manzano, S.; Carrión, J.S.; López-Merino, L.; Ochando, J.; Munuera, M.; Fernández, S.; González-Sampériz, P. Early to Mid-Holocene spatiotemporal vegetation changes and tsunami impact in a paradigmatic coastal transitional system (Doñana National Park, southwestern Europe). Glob. Planet. Chang. 2018, 161, 66–81. [Google Scholar] [CrossRef]
- Sousa, A.; García-Murillo, P. Historia ecológica y evolución de las lagunas peridunares del Parque Nacional de Doñana. In Madrid: Naturaleza y Parques Nacionales-Serie Técnica; Organismo Autónomo de Parques Nacionales, Ministerio de Medio Ambiente: Madrid, Spain, 2005. [Google Scholar]
- Shive, K.L.; Wuenschel, A.; Hardlund, L.J.; Morris, S.; Meyer, M.D.; Hood, S.M. Ancient trees and modern wildfires: Declining resilience to wildfire in the highly fire-adapted giant sequoia. For. Ecol. Manag. 2002, 511, 120110. [Google Scholar] [CrossRef]
- Cansler, C.A.; Hood, S.M.; Varner, J.M.; Van Mantgem, P.J.; Agne, M.C.; Andrus, R.A.; Ayres, M.P.; Ayres, B.D.; Bakker, J.D.; Battaglia, M.A.; et al. The Fire and Tree Mortality Database, for empirical modeling of individual tree mortality after fire. Sci. Data 2020, 7, 194. [Google Scholar] [CrossRef] [PubMed]
- Woolley, T.; Shaw, D.C.; Ganio, L.M.; Fitzgerald, S. A review of logistic regression models used to predict post-fire tree mortality of western North American conifers. Int. J. Wildl. Fire 2012, 21, 1. [Google Scholar] [CrossRef]
- Varner, J.M.; Hood, S.M.; Aubrey, D.P.; Yedinak, K.; Hiers, J.K.; Jolly, W.M.; Shearman, T.M.; McDaniel, J.K.; O’Brien, J.J.; Rowell, E.M. 2021. Tree crown injury from wildland fires: Causes, measurement and ecological and physiological consequences. New Phytol. 2021, 231, 1676–1685. [Google Scholar] [CrossRef] [PubMed]
- Fritts, H.C. Tree-Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Larsson, L.A.; Larsson, P.O. CDendro and CooRecorder (ver. 9.3.1); Cybis Elektronik and Data AB: Saltsjöbaden, Sweden, 2018. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 68–78. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 29 July 2022).
- Saurer, M.; Siegwolf, R.T.W.; Schweingruber, F.H. Carbon isotope discrimination indicates improving water–use efficiency of trees in northern Eurasia over the last 100 years. Glob. Chang. Biol. 2004, 10, 2109–2120. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 1982, 9, 121. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Ann. Rev. Plant Physiol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Beghin, R.; Cherubini, P.; Battipaglia, G.; Siegwolf, R.; Saurer, M.; Bovio, G. Tree-ring growth and stable isotopes (13C and 15N) detect effects of wildfires on tree physiological processes in Pinus sylvestris L. Trees Struct. Funct. 2011, 25, 627–636. [Google Scholar] [CrossRef]
- Battipaglia, G.; Strumia, S.; Esposito, A.; Giuditta, E.; Sirignano, C.; Altieri, S.; Rutigliano, F.A. The effects of prescribed burning on Pinus halepensis mill. as revealed by dendrochronological and isotopic analyses. For. Ecol. Manag. 2014, 334, 201–208. [Google Scholar] [CrossRef]
- Alfaro-Sánchez, R.; Camarero, J.J.; Sangüesa-Barreda, G.; de las Heras, J. Post-fire Aleppo pine growth, C and N isotope composition depend on site dryness. Trees Struct. Funct. 2016, 30, 581–595. [Google Scholar] [CrossRef]
- Battipaglia, G.; Savi, T.; Ascoli, D.; Castagneri, D.; Esposito, A.; Mayr, S.; Nardini, A. Effects of prescribed burning on ecophysiological, anatomical and stem hydraulic properties in Pinus pinea L. Tree Physiol. 2016, 36, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Valor, T.; Casals, P.; Altieri, S.; González-Olabarria, J.R.; Piqué, M.; Battipaglia, G. Disentangling the effects of crown scorch and competition release on the physiological and growth response of Pinus halepensis Mill. using δ13C and δ18O isotopes. For. Ecol. Manag. 2018, 424, 276–287. [Google Scholar] [CrossRef]
- Oliveras, I.; Martinez-Vilalta, J.; Jimenez-Ortiz, T.; Lledó, J.M.; Escarré, A.; Piñol, J. Hydraulic properties of Pinus halepensis, Pinus pinea and Tetraclinis articulata in a dune ecosystem of Eastern Spain. Plant Ecol. 2003, 169, 131–141. [Google Scholar] [CrossRef]
- Antunes, C.; Díaz Barradas, M.C.; Zunzunegui, M.; Vieira, S.; Pereira, A.; Anjos, A.; Correia, O.; Pereira, M.J.; Máguas, C. Contrasting plant water-use responses to groundwater depth in coastal dune ecosystems. Funct. Ecol. 2018, 32, 1931–1943. [Google Scholar] [CrossRef]
- Rigolot, E. Predicting postfire mortality of Pinus halepensis Mill. and Pinus pinea L. Plant Ecol. 2004, 171, 139–151. [Google Scholar] [CrossRef]
- Fernandes, P.; Vega, J.A.; Jimenez, E.; Rigolot, E. Fire resistance of European pines. For. Ecol. Manag. 2008, 256, 246–255. [Google Scholar] [CrossRef]
- Madrigal, J.; Souto-García, J.; Calama, R.; Guijarro, M.; Picos, J.; Hernando, C. Resistance of Pinus pinea L. bark to fire. Int. J. Wildland Fire 2019, 28, 342–353. [Google Scholar] [CrossRef]
- Vales, J.J.; Pino, I.; Granado, L.; Prieto, R.; Méndez, E.; Rodríguez, M.; Giménez de Azcárate, F.; Ortega, E.; Moreira, J.M. Damage assessment and recovery mapping for the “Las Peñuelas” wildfire, Moguer (Huelva). Satellite Imagery. Year 2017. Rev. Teledetección 2020, 57, 79–94. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Johnson, E.A. How forest fires kill trees: A review of the fundamental biophysical processes. Scand. J. For. Res. 2007, 22, 500–515. [Google Scholar] [CrossRef]
- Barker, J.S.; Gray, A.N.; Fried, J.S. The effects of crown scorch on post-fire delayed mortality are modified by drought exposure in California (USA). Fire 2022, 5, 21. [Google Scholar] [CrossRef]
- Alcahud, M.; Martínez, J.J.; Orozco, E. Influencia de los incendios forestales sobre la regeneración y distribución de la sabina negral (Juniperus phoenicea L.) en las provincias de Albacete y Murcia. In Actas del II Congreso Forestal Español; SECF: Pamplona, Spain, 1997; Volume 5, pp. 27–29. [Google Scholar]
- van Mantgem, P.J.; Falk, D.A.; Williams, E.C.; Das, A.J.; Stephenson, N.L. Pre-fire drought and competition mediate post-fire conifer mortality in western U.S. National Parks. Ecol. Appl. 2018, 28, 1730–1739. [Google Scholar] [CrossRef]
- Jones, M.W.; Abatzoglou, J.T.; Veraverbeke, S.; Andela, N.; Lasslop, G.; Forkel, M.; Smith, A.J.P.; Burton, C.; Betts, R.A.; van der Werf, G.R.; et al. Global and regional trends and drivers of fire under climate change. Rev. Geophys. 2022, 60, e2020RG000726. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name (Type) | Latitude (N) | Longitude (W) | Elevation (m a.s.l.) | Slope (º) |
|---|---|---|---|---|
| Corral del Muerto (slack) | 37.06007 | 6.65684 | 35 | 0–5 |
| Duna (dune) | 37.05849 | 6.65761 | 70 | 20–45 |
| Species | Site | No. Individuals | DBH (cm) | Height (m) | Crown Damage (%) | Bark Char (%) |
|---|---|---|---|---|---|---|
| Pine | Inter-dune | 30 | 31.9 ± 0.7b | 9.0 ± 0.3b | 46.4 ± 6.1a | 20.7 ± 2.7a |
| Dune | 17 | 16.2 ± 0.9a | 3.4 ± 0.2a | 69.4 ± 3.7b | 33.2 ± 3.0b | |
| Juniper | Inter-dune | 17 | 7.1 ± 0.3 | 2.7 ± 0.2 | 48.4 ± 4.9 | Not assessed |
| Dune | 20 | 6.8 ± 0.4 | 2.4 ± 0.1 | 56.0 ± 6.0 | Not assessed |
| Species | Site | Period | BAI (cm2) | MSx | rbar | EPS |
|---|---|---|---|---|---|---|
| Pine | Inter-dune | 1954–2020 | 6.94 ± 0.38b | 0.32 | 0.57 | 0.92 |
| Dune | 1963–2020 | 2.15 ± 0.18a | 0.36 | 0.71 | 0.97 | |
| Juniper | Inter-dune | 1948–2020 | 1.13 ± 0.07b | 0.20 | 0.32 | 0.87 |
| Dune | 1926–2020 | 0.87 ± 0.05a | 0.25 | 0.37 | 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camarero, J.J.; Díaz-Delgado, R.; Colangelo, M.; Valeriano, C.; Sánchez-Salguero, R.; Madrigal, J. Differential Post-Fire Recovery of Tree and Shrub Growth and Water-Use Efficiency in a Mediterranean Coastal Dune System. Fire 2022, 5, 135. https://doi.org/10.3390/fire5050135
Camarero JJ, Díaz-Delgado R, Colangelo M, Valeriano C, Sánchez-Salguero R, Madrigal J. Differential Post-Fire Recovery of Tree and Shrub Growth and Water-Use Efficiency in a Mediterranean Coastal Dune System. Fire. 2022; 5(5):135. https://doi.org/10.3390/fire5050135
Chicago/Turabian StyleCamarero, Jesús Julio, Ricardo Díaz-Delgado, Michele Colangelo, Cristina Valeriano, Raúl Sánchez-Salguero, and Javier Madrigal. 2022. "Differential Post-Fire Recovery of Tree and Shrub Growth and Water-Use Efficiency in a Mediterranean Coastal Dune System" Fire 5, no. 5: 135. https://doi.org/10.3390/fire5050135
APA StyleCamarero, J. J., Díaz-Delgado, R., Colangelo, M., Valeriano, C., Sánchez-Salguero, R., & Madrigal, J. (2022). Differential Post-Fire Recovery of Tree and Shrub Growth and Water-Use Efficiency in a Mediterranean Coastal Dune System. Fire, 5(5), 135. https://doi.org/10.3390/fire5050135