Abstract
Giant sequoia groves, located on the western slope of the central and southern Sierra Nevada mountains in California, USA, have been experiencing regeneration failure for more than a century due to the exclusion of wildfires. Giant sequoias are serotinous conifers and have evolved a strong relationship between high-severity fire and reproduction. While this relationship is widely recognized, only one previous peer-reviewed study has directly investigated giant sequoia reproduction and fire severity, and that study used different fires for each severity class. We conducted a study of giant sequoia reproduction and fire severity in a single fire, the KNP Complex fire of 2021, within the Redwood Mountain Grove in Sequoia and Kings Canyon National Park. We found that giant sequoia seedlings are more dominant relative to other conifer species and are growing faster in a large high-severity fire area than in adjacent low/moderate-severity areas. Distance to the nearest live sequoia seed source was not a significant factor in sequoia seedling density. Our results call into question the basis for widespread plans and projects designed to prevent high-severity fires and should reevaluate moving forward with proposed tree planting activities in high-severity fire areas within giant sequoia groves.
1. Introduction
Serotinous conifers around the world have a deep evolutionary history linking high-severity fire and reproduction, spanning tens of millions of years [1,2,3]. This includes genera such as Callitris in Australia and New Caledonia and Widdringtonia in South Africa in the Southern Hemisphere [4,5], some Pinus species in the Mediterranean [6], and numerous species in North America, such as the bishop pine (Pinus muricata D. Don., [7]), the Rocky Mountain lodgepole pine (Pinus contorta var. latifolia Engelm., [8], and jack pine (Pinus banksiana Lamb., [9,10]. Among the serotinous conifers of North America is the giant sequoia (Sequoiadendron giganteum [Lindl.] J.Buchh., [11]), which exists in several dozen groves on the western slope of the central and southern Sierra Nevada mountains in California [12].
As an evolutionary adaptation, serotiny in modern conifer families, Pinaceae and Cuppressaceae, developed during the Cretaceous, 65–106 million years ago, ensuring that seed release from conifer cones (triggered by heat from wildfire) and seedling germination occur during the optimal conditions for establishment and growth that are created by intense fire [2]. For giant sequoias, these optimal conditions, created in particular by patches of crown fire, have been described as including the consumption of all duff and litter on the forest floor and conversion of this material into a nutrient-rich bed of mineral ash, as well as high or complete mortality of the forest canopy, which provides additional sunlight that facilitates growth of seedlings and saplings [11,13]. Seedling germination and establishment are unlikely in the absence of fire.
Giant sequoias produce many new cones each year at varying rates; seeds appear to remain viable for many years (the upper limit is unknown). Seed release is mainly driven by heat from wildfires, and successful germination and seedling establishment is almost entirely limited to the conditions created by fire, particularly by more intense fire [11,13]. In some cases, sequoia seeds can be released by squirrels chewing on cones, native wood-boring insects chewing into cones, or cone-bearing branches breaking and falling during storms [11].
While the relationship between high-severity fire and serotinous conifers is well documented in general, research on this question is very limited with regard to giant sequoias. Only one peer-reviewed published study, Meyer and Safford (2011) [14], compares giant sequoia reproduction in a high-severity fire area to moderate-severity and low-severity areas. Meyer and Safford (2011) [14] concluded that giant sequoia reproduction benefits from high-severity and moderate-severity fire but not low-severity fire. However, the high-severity fire patch studied by Meyer and Safford (2011) [14] was only 20 ha in size, and the study did not compare different fire severities within the same fires.
In 2020 and 2021, three large lightning-ignited wildfires spread through numerous extant giant sequoia groves. The “KNP Complex” lightning fire of 2021 burned mostly in Sequoia and Kings Canyon National Parks and included a single high-severity fire patch encompassing >150 ha of the southern portion of the Redwood Mountain giant sequoia grove. The National Park Service recently began implementation of a tree planting project in this high-severity fire patch [15], based upon two recent studies, Stephenson et al. (2023) (pre-print) [16] and Soderberg et al. (2024, in press) [17]. These studies asserted that post-fire sequoia regeneration would be inadequate in this large high-severity fire area, and some smaller high-severity fire patches in other sequoia groves, and that these high-severity fire patches would be at risk of reproduction failure due to long distances from live giant sequoia seed source trees. Despite the conclusions by Meyer and Safford (2011) [14], Stephenson et al. (2023) [16] and Soderberg et al. (2024) [17] promoted predominantly low-severity prescribed fire as a more beneficial management approach for giant sequoia groves. However, neither of these studies investigated whether there is some post-fire sequoia seedling density threshold below which reproduction failure is likely, and neither analyzed giant sequoia reproduction in low/moderate-severity areas compared to high-severity fire areas.
There is a need to better understand the relationship between fire severity and giant sequoia reproduction, particularly in larger high-severity fire patches. In this study, we investigated giant sequoia reproduction after the KNP Complex fire in the large high-severity fire patch in Redwood Mountain grove and adjacent low/moderate-severity burned areas located in the same grove. Specifically, we explored four null hypotheses: (1) giant sequoia seedling density would not be correlated with distance to live, surviving giant sequoia seed source trees (the distance effect hypothesis); (2) giant sequoia seedling density would not differ in high-severity areas versus low/moderate-severity areas (the fire severity hypothesis); (3) giant sequoia seedling proportion, relative to other conifer species, would not differ in high-severity areas versus low/moderate-severity areas (the sequoia dominance hypothesis); and (4) giant sequoia seedling height would not differ in high-severity areas versus low/moderate-severity areas (the sequoia growth hypothesis).
2. Materials and Methods
Our study was located in the southern portion of the Redwood Mountain giant sequoia grove in Sequoia and Kings Canyon National Parks within >150 ha of burned forest that experienced high-severity fires and adjacent low/moderate-severity areas in the summer and early fall of 2023, two years after the KNP Complex fire. Fire severity in this study was determined using Relative delta Normalized Burn Ratio (RdNBR) satellite imagery data from U.S. Geological Survey’s Rapid Assessment of Vegetation Condition after Wildfire (RAVG) database (https://burnseverity.cr.usgs.gov/products/ravg, accessed on 10 January 2024). We used the Relative delta Normalized Burn Ratio (RdNBR) thresholds identified by Miller and Thode (2007) [18] to categorize fire severity, as follows: low severity (0–25% basal area mortality); moderate severity (26–75% basal area mortality); and high severity (76–100% basal area mortality).
To investigate hypothesis 1, before field visits were conducted between 30 June and 5 August 2023, we established four a priori (prior to visiting the locations) parallel transects in the large high-severity area, with eight plots in each linear transect, and plot centers spaced by 100 m (Figure 1). At each plot, we recorded the distance (m) to the nearest live, cone-bearing giant sequoia using a laser hypsometer. We checked this result for each plot with satellite imagery to ensure that no sequoias with remaining green crowns and in closer proximity to the plot center were missed. In each plot, we recorded the total number of live sequoia seedlings. We measured sequoia seedlings within circular plots extending from the plot center with radii of 1 m, 2 m, 5 m, or 10 m as needed to include one or more sequoia seedlings. We also recorded whether sequoia seedlings were from 2022 (one year post-fire) or 2023 (two years post-fire). We could identify 2023 sequoia seedlings by their small size (just a few to several cm tall by late June of 2023) and also by the reddish stems that they have when they are days or weeks old. The reddish color quickly fades thereafter.
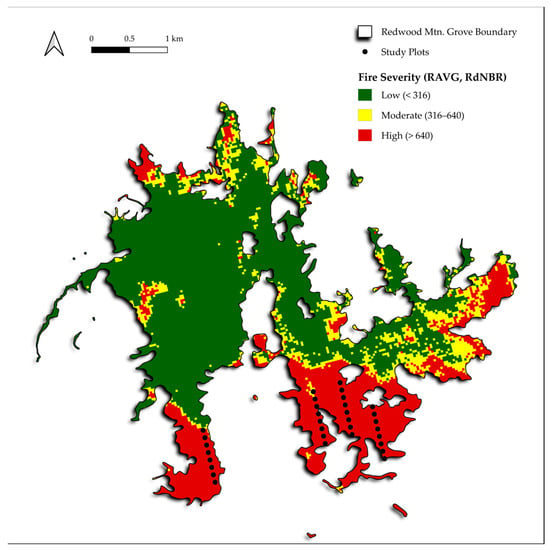
Figure 1.
The Redwood Mountain Grove study area, fire severity, and transects shown were used to investigate the correlation between sequoia seedling density and distance to live sequoia seed sources. White spaces are mixed-conifer forests outside of the Redwood Mountain Grove boundaries.
For all plots, we converted data to sequoia seedlings per hectare. We analyzed whether there is a correlation between distance to the nearest live sequoia seed tree and sequoia seedling density using Spearman’s rank correlation test [19,20]. This is an appropriate correlation test for non-parametric data when the assumptions required for parametric tests are not met due to highly skewed distributions, for example, as in the case of our data [21].
The sample plots for this study met two criteria: the plots were physically present within the Sequoia and Kings Canyon National Park-mapped sequoia grove boundary, and plot centers were located within 100 m of a (pre-fire) established sequoia. We excluded 1 of our 32 plots for not meeting both of these criteria.
Hypotheses 2 (fire severity hypothesis), 3 (sequoia dominance hypothesis), and 4 (sequoia growth hypothesis) were tested within a two-week period (to ensure the closest feasible comparison between high-severity and low/moderate-severity) from 29 September 2023 to 11 October 2023. Five a priori transects were located perpendicular to the boundary between the large high-severity patch and low/moderate severity area, with three plots 100 m, 200 m, and 300 m from the high-severity patch boundary into the low/moderate-severity area and three plots 100 m, 200 m, and 300 m from the high-severity patch boundary into the interior of the large high-severity fire patch (Figure 2). For each of the three hypotheses (2, 3, and 4), we analyzed 15 plots in the large high-severity fire patch and 15 plots in the adjacent low/moderate-severity area. We used the same variable-radius plot system that we used for hypothesis 1 to census sequoia seedlings, as described above, and converted values to seedlings per hectare. Within each circular area representing a plot, we recorded the total number of live sequoia seedlings (for the fire severity hypothesis), the total number of live non-sequoia conifer seedlings (for the sequoia dominance hypothesis), and the height (cm) of the tallest sequoia seedling (for the sequoia growth hypothesis). For hypotheses 2, 3, and 4, we analyzed whether there are differences between high-severity and low/moderate severity plots (the unit of replication) with regard to sequoia seedling density (fire severity hypothesis), sequoia seedling proportion (sequoia dominance hypothesis), and maximum sequoia seedling height (sequoia growth hypothesis) using a Mann–Whitney test [21]. The Mann–Whitney test is an appropriate test for comparing two groups when data do not meet the required assumptions for parametric tests due to highly skewed distributions, for instance.
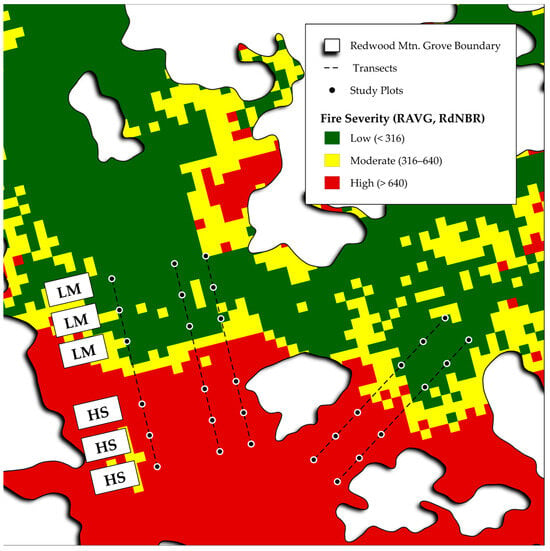
Figure 2.
The Redwood Mountain Grove study area, fire severity, and transects shown were used to investigate differences between high-severity areas and low/moderate-severity areas in terms of sequoia seedling density, maximum height, and proportion relative to other conifers.
When data are non-normally distributed, as they are in our study area (Kolmogorov–Smirnov test, p < 0.05), non-parametric tests tend to have greater statistical power [21,22].
3. Results
With regard to hypothesis 1, the distance effect hypothesis, there was no correlation between distance to the nearest live, cone-bearing giant sequoia and post-fire sequoia seedling density (rs = −0.247, p = 0.180, Figure 3), and the null hypothesis was not rejected.
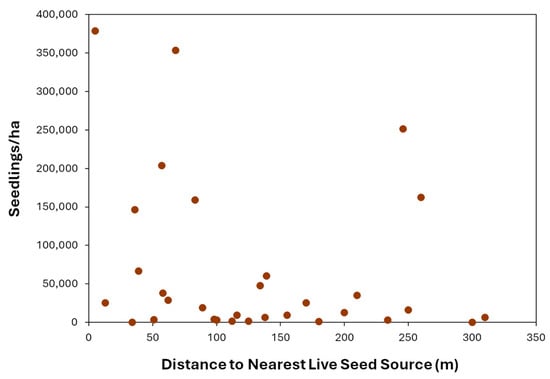
Figure 3.
Naturally regenerating giant sequoia seedlings per square meter and distance (m) to the nearest live sequoia seed source.
With regard to hypothesis 2, the comparison between the high-severity fire patch and the adjacent low/moderate-severity areas in terms of sequoia seedling density (fire severity hypothesis), there was no difference in post-fire sequoia seedling density (U = 88, p = 0.317, Table 1), and the null hypothesis was not rejected. The median sequoia seedling density at two years post-fire was 3183/ha in the low/moderate-severity areas and 12,732/ha in the high-severity areas (mean densities were much higher for both, but we do not report means due to the skewed nature of the data). The proportion of giant sequoia seedlings among all conifer reproduction (hypothesis 3, the sequoia dominance hypothesis) was significantly higher in the high-severity areas than in the low/moderate-severity areas (U = 36, p = 0.002, Table 1), and the null hypothesis was rejected. The median sequoia seedling proportion was 0.69 in low/moderate-severity areas and 1.00 in high-severity areas. Last, the maximum height per plot of giant sequoia seedlings (hypothesis 4, the sequoia growth hypothesis) was significantly greater in high-severity fire areas than in low/moderate-severity areas (U = 16.5, p < 0.001, Table 1), and the null hypothesis was rejected. The median maximum sequoia seedling height was 10 cm in low/moderate-severity areas and 32 cm in high-severity areas.

Table 1.
Summary of (a) low/moderate-severity (LM) and (b) high-severity (HS) fire plot data regarding the fire severity hypothesis (seedlings/ha), the sequoia dominance hypothesis (proportion of conifer seedlings comprised by giant sequoias), and the sequoia growth hypothesis (height in cm).
All raw data, including plot coordinates, are available in the Supplemental Material.
4. Discussion
In the large high-severity fire patch in the Redwood Mountain grove, giant sequoia reproduction two years after the KNP Complex lightning fire was more dominant (relative to other conifer species), and the height of giant sequoia seedlings was significantly greater (Figure 4 and Figure 5) than in adjacent low/moderate-severity areas. The distance to live, surviving sequoias was not a significant factor.

Figure 4.
Typical natural sequoia regeneration in the interior of the large high-severity fire patch at two years post-fire in the Redwood Mountain Grove. Shown: Ara Marderosian, Sequoia ForestKeeper. Photo by Chad Hanson.

Figure 5.
Highly abundant sequoia seedlings in the large high-severity fire patch in Redwood Mountain Grove. Photo by Chad Hanson.
The density of sequoia seedlings was not significantly different between high-severity and low/moderate-severity areas, though, numerically, the median density was three times larger in high-severity areas than in low/moderate-severity areas. It is possible that this difference would be statistically significant with a larger sample size, and additional research in other recently burned sequoia groves will be needed to answer this question.
Soderberg et al. (2024) [17] reported a substantially lower sequoia seedling density in the high-severity fire areas of the Redwood Mountain grove relative to our results. However, the results of Soderberg et al. (2024) [17] were based on field surveys conducted in 2022 at one year post-fire. We conducted our field surveys in 2023, two years post-fire, and 41% of our plots had sequoia seedlings exclusively from 2023.
Our results and previous work [11,13,14] suggest several mechanisms that work to provide advantageous conditions for giant sequoia reproduction in high-severity fire areas, including (a) intense heat that facilitates the release of a super-abundance of seeds; (b) consumption of all duff and litter, which allows sequoia seedlings to sink their roots into soil, and which provides a layer of mineral ash that aids growth over time; (c) mortality of most or all canopy trees, which provides high levels of sunlight to sequoia seedlings; (d) a competitive advantage over non-sequoia conifers in terms of the proportion of total species composition, at least in the earlier stages of natural succession; and (e) an extraordinary growth rate.
Similar to recent results regarding natural regeneration of a related species, the coast redwood (Sequoia sempervirens) [23,24], the results here are encouraging for the conservation of giant sequoias. Further research will be needed in other sequoia groves through which large fires have spread recently to determine how broadly applicable our results here may be. In the meantime, our results here call into question current projects and plans designed to prevent high-severity fire in sequoia groves and to conduct widespread planting of sequoia seedlings in high-severity fire areas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fire7020044/s1, Spreadsheets S1 (Seedling Density and Distance to Live Sequoias), S2 (Seedling Density by Fire Severity, Seedling Height, Sequoia Proportion), and S3 (Coordinates).
Author Contributions
Conceptualization, C.T.H., T.Y.C., M.K. and B.C.B.; methodology, C.T.H., T.Y.C., M.K. and B.C.B.; software, B.C.B.; validation, C.T.H., T.Y.C., M.K. and B.C.B.; formal analysis, C.T.H.; investigation, C.T.H., T.Y.C., M.K., B.C.B. and C.S.; resources, C.T.H.; data curation, C.T.H.; writing—original draft preparation, C.T.H.; writing—review and editing, C.T.H., T.Y.C., M.K. and B.C.B.; visualization, C.T.H., T.Y.C., M.K. and B.C.B.; supervision, C.T.H.; project administration, C.T.H.; funding acquisition, C.T.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Environment Now foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All raw data are available in the Supplemental Material, and all GIS data are available upon request from Bryant Baker at: bryant@wildlandmaps.org.
Acknowledgments
We thank Environment Now for supporting this research. We also thank the Academic Editor and the reviewers, whose thoughtful guidance and comments helped to improve this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- He, T.; Belcher, C.M.; Lamont, B.B.; Lim, S.L. A 350-million-year legacy of fire adaptation among conifers. J. Ecol. 2016, 104, 352–363. [Google Scholar] [CrossRef]
- Lamont, B.B.; He, T.; Yan, Z. Evolutionary history of fire-stimulated resprouting, flowering, seed release and germination. Biol. Rev. 2019, 94, 903–928. [Google Scholar] [CrossRef] [PubMed]
- Lamont, B.B.; Pausas, J.G.; He, T.; Witkowski, E.T.F.; Hanley, M.E. Fire as a Selective Agent for Both Serotiny and Non-serotiny Over Space and Time. Crit. Rev. Plant Sci. 2020, 39, 140–172. [Google Scholar] [CrossRef]
- Ladd, P.G.; Midgely, J.J.; Nield, A.P. Serotiny in southern hemisphere conifers. Aust. J. Bot. 2013, 61, 486–496. [Google Scholar] [CrossRef][Green Version]
- Ladd, P.G.; Zhao, X.; Enright, N.J. Fire regime and climate determine spatial variation in level of serotiny and population structure in a fire-killed conifer. Plant Ecol. 2022, 223, 849–862. [Google Scholar] [CrossRef]
- Fernández-García, V.; Fulé, P.Z.; Marcos, E.; Calvo, L. The role of fire frequency and severity on the regeneration of Mediterranean serotinous pines under different environmental conditions. For. Ecol. Manag. 2019, 444, 59–68. [Google Scholar] [CrossRef]
- Bisbing, S.M.; Urza, A.K.; York, R.A.; Hankin, L.E.; Putz, T.R. Persistent, viable seedbank buffers serotinous bishop pine over a broad fire return interval. Fire Ecol. 2023, 19, 35. [Google Scholar] [CrossRef]
- Schoennagel, T.; Turner, M.G.; Romme, W.H. The influence of fire interval and serotiny on postfire lodgepole pine density in Yellowstone National Park. Ecology 2003, 84, 2967–2978. [Google Scholar] [CrossRef]
- Briand, C.H.; Schwilk, D.W.; Gauthier, S.; Bergeron, Y. Does fire regime influence life history traits of jack pine in the southern boreal forest of Québec, Canada. Plant Ecol. 2015, 216, 157–164. [Google Scholar] [CrossRef]
- Pelletier, E.; de Lafontaine, G. Jack pine of all trades: Deciphering intraspecific variability of a key adaptive trait at the rear edge of a widespread fire-embracing North American conifer. Am. J. Botany 2023, 110, e16111. [Google Scholar] [CrossRef] [PubMed]
- Harvey, H.T.; Shellhammer, H.S.; Stecker, R.E. Giant Sequoia Ecology; USDI National Park Service: Washington, DC, USA, 1980. [Google Scholar]
- USDA. Giant Sequoia National Monument, Final Environmental Impact Statement; U.S.D.A. Forest Service, Sequoia National Forest: Porterville, CA, USA, 2012. [Google Scholar]
- Stephenson, N.L. Long-term dynamics of giant sequoia populations: Implications for managing a pioneer species. In The Symposium on Giant Sequoias: Their Place in the Ecosystem and Society; USDA Forest Service Pacific Southwest Research Station General Technical Report PSW-GTR-151: Albany, CA, USA, 1994; pp. 56–63. [Google Scholar]
- Meyer, M.D.; Safford, H.D. Giant sequoia regeneration in groves exposed to wildfire and retention harvest. Fire Ecol. 2011, 7, 2–16. [Google Scholar] [CrossRef]
- USDOI. Re-Establish Tree Seedlings in Severely Burned Giant Sequoia Groves and Adjacent Fisher Habitat, Revised Environmental Assessment; Sequoia and Kings Canyon National Parks; U.S. Department of the Interior: Three Rivers, CA, USA, 2023. [Google Scholar]
- Stephenson, N.L.; Caprio, A.C.; Soderberg, D.N.; Das, A.J.; Lopez, E.L.; Williams, A.P. Post-fire reference densities for giant sequoia seedlings. EcoEvoRxiv 2023. Available online: https://ecoevorxiv.org/repository/view/5457/ (accessed on 15 July 2023).
- Soderberg, D.N.; Das, A.J.; Stephenson, N.L.; Meyer, M.D.; Brigham, C.A.; Flickinger, J. Assessing giant sequoia mortality and regeneration following high severity wildfire. Ecosphere 2024, in press. [Google Scholar]
- Miller, J.D.; Thode, A.E. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens. Envir. 2007, 109, 66–80. [Google Scholar] [CrossRef]
- Rosner, A. Fundamentals of Biostatistics, 5th ed.; Pacific Grove: Duxbury, MA, USA, 2000. [Google Scholar]
- Glantz, S.A. Primer of Biostatistics, 6th ed.; McGraw-Hill: New York, NY, USA, 2005. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall/Pearson: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Yue, S.; Pilon, P.; Cavadias, G. Power of Mann-Kendall and Spearman’s rho tests for detecting monotonic trends in hydrological series. J. Hydrol. 2002, 259, 254–271. [Google Scholar] [CrossRef]
- Mahdizadeh, M.; Russell, W. Initial Floristic Response to High Severity Wildfire in an Old-Growth Coast Redwood (Sequoia sempervirens (D. Don) Endl.) Forest. Forests 2021, 12, 1135. [Google Scholar] [CrossRef]
- Peltier, D.M.P.; Carbone, M.S.; Enright, M.; Marshall, M.C.; Trowbridge, A.M.; LeMoine, J.; Koch, G.; Richardson, A.D. Old reserves and ancient buds fuel regrowth of coast redwood after catastrophic fire. Nat. Plants 2023, 9, 1978–1985. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).