Abstract
Wildfires in the Mediterranean basin, particularly in Portugal, pose significant ecological challenges by altering landscapes and ecosystems. This study examines vegetation recovery in Serra do Açor seven years after the 2017 wildfires, using remote sensing and field data to analyze post-fire dynamics. The primary goal was to assess whether fire severity, measured via the dNBR index from Sentinel-2 imagery, impacts vegetation recovery or if site-specific factors and pre-fire floristic composition are more influential. Randomly assigned plots based on previous land use and fire severity were analyzed for floristic attributes. To quantify and classify cover changes, a supervised classification methodology based on the random forest algorithm was applied to Sentinel-2 data. The results showed no clear link between fire severity and recovery; instead, local factors like soil and topography, along with dominant pre-fire species, influenced recovery. Acacia and eucalyptus communities grew faster and increased the occupied area but exhibited lower diversity than native vegetation communities. Supervised classifications achieved high accuracy (Kappa > 0.90), showing increased shrubland areas and expansion of eucalyptus and acacia. The study highlights the methodology’s effectiveness and potential for broader applications in future research.
1. Introduction
Portugal, situated within the Mediterranean basin, is highly susceptible to wildfires due to its hot and dry summers, which make it one of the European Union’s most affected countries. Over the past 30 years, approximately 700,000 fire ignitions have been recorded, burning over 4.5 million hectares. In 2017, the country experienced unprecedented wildfire activity, with 540,000 hectares consumed and more than 100 fatalities reported [1]. Asymmetric intra-annual precipitation patterns have historically influenced wildfire development, and inter-annual variations add further complexity. While wetter years may result in less intense wildfire seasons, the subsequent increase in vegetation growth can heighten the risk in following drier years [2,3]. Vegetation flammability is strongly influenced by its moisture content [4]. This steadily decreases during the dry months when extremely dry fuels facilitate rapid fire spread and increase the risk of wildfire peaking just before the onset of the rainy season, at the end of summer and/or in early autumn [5]. Catastrophic fire events are often linked to particularly dry periods and extreme weather conditions [6], as observed in Portugal during the fire seasons of 2003 [7], 2005 [8], and 2017 [9]. While climatic and meteorological factors undoubtedly contribute to the development of large and intense wildfires, they are not the sole drivers of changes in fire regimes [10,11,12]. Fire regimes have in fact undergone significant changes in recent decades, with increases not only in frequency but also in affected area and severity [13,14,15], a result of land-use dynamics that have altered fuel characteristics [16], particularly after the expansion of fire-prone communities such as monocultures (Pinus, Eucalyptus) [17,18,19], dense and highly flammable shrublands [20], and the spread of exotic species (Acacia sp.; Hakea sp.) [21]. This trend specifically affected the rural landscapes of northern and central Portugal, where larger fires (>500 ha) predominantly occur in low-population-density regions [22], even though a greater number of ignitions, mainly of anthropogenic origin [23], are registered in densely populated areas [24]. These increases in the frequency and affected areas are expected to worsen in the future as average temperatures rise and precipitation declines, which will increase evapotranspiration and, consequently, the flammability of vegetation [3,25].
Although the current frequency of wildfires is clearly linked to anthropogenic action, fire is a natural agent of Mediterranean ecosystems, and plant communities have evolved to adapt to this disturbance. The prevalence of species that rely on vegetative regeneration (sprouters) as a post-fire recovery strategy, through the rapid production of shoots from roots, stems, or branches, depending on the severity of the damage, can be seen as an indicator of adaptation [6]. Species that regenerate exclusively from seeds (seeders), which tend to develop large seed banks in the soil, are more associated with pioneer communities [17,26].
Plant communities are influenced by a range of biophysical factors, including soil characteristics (e.g., depth, composition), topographic features (e.g., slope, aspect, altitude), climatic conditions (e.g., precipitation, temperature, relative humidity), and proximity to water sources (affecting moisture availability, nutrient flux, and erosion processes). These factors collectively shape both the composition and structural traits of vegetation [27,28,29,30]. Conversely, during a wildfire, the same factors affect fire intensity, measured by the energy released during combustion, and fire severity, which reflects the damage to vegetation and soil [31]. In the short term, severity strongly correlates with vegetation recovery, as it determines the survival of plants and the damage to seed banks [32,33]. In the long term, climate conditions significantly influence vegetation recovery, particularly disturbances such as droughts, which are considered the greatest risk in climates with dry seasons [34,35].
However, several authors state that the most critical factor, across both temporal scales, is pre-fire floristic composition and its interaction with the biophysical environment [30,36,37]. In other words, dominant species and their evolutionary traits determine both the capacity and speed of regeneration [38,39]. Resilience decreases as vegetation height increases: grasslands and shrublands are less likely to alter their cover, whereas forests, despite their ability to resist low-intensity surface fires, can experience significant changes when exposed to crown fires [40]. Tree communities dominated by obligate seeders (e.g., Pinus species) are considered less stable than those dominated by sprouters such as Quercus species or Castanea sativa Mill., and require longer recovery periods to regain pre-fire development levels [41,42,43]. Here, vegetative regeneration is often the most stable strategy under frequent fire conditions, as it relies on pre-established root systems that enable a faster and more effective response, thereby minimizing fire impacts [44,45].
Under natural conditions, post-fire floristic composition and diversity tend to remain relatively stable [46,47]. However, the introduction of exotic species presents novel challenges. Adapted to fire in their native ecosystems, such as parts of Australia, many of these species have developed regeneration strategies that surpass those of Mediterranean species, giving them significant competitive advantages in the immediate post-fire environment. This often leads to the suppression of native vegetation and profound alterations in the floristic composition of plant communities [48,49]. Adaptations such as abundant seed production, heat-stimulated germination, and rapid growth rates enable these species to quickly colonize new territory [50,51,52]. This can establish a feedback loop, where changes in fire regime properties favor the further expansion of these species [53]. Such dynamics have been observed in association with previous wildfire events [54,55], which tends to reinforce their invasive potential and amplify their ecological impacts.
The dynamics of vegetation recovery and its drivers have been studied in various ecological contexts [28,35,56]. In fact, post-fire monitoring of vegetation recovery and cover changes is essential for land management and damage prevention [57], particularly in mitigating habitat loss and biodiversity decline, which are two of the many risks associated with altered fire regimes and vegetation dynamics [58,59]. Remote sensing-based methodologies offer an effective and cost-efficient approach to analyzing vegetation recovery across large and heterogeneous study areas [57,60,61]. Their growing adoption is driven by the increasing availability and quality of open-access data from Earth observation missions, with Sentinel-2 standing out due to its high spatial resolution and frequent revisit times. Disturbance-induced shifts can be detected through spectral indices that reflect biophysical variables, such as the normalized difference vegetation index (NDVI) for vegetation conditions, and the normalized burn ratio (NBR) or its differential version (dNBR), which are commonly used to delineate burnt areas and estimate fire severity [62,63]. However, these types of data are unable to capture structural complexity or vegetation recovery stages based on plant height, highlighting the importance of incorporating fieldwork to validate and complement remotely derived data [36].
To address these topics, this study aims to comprehensively assess post-fire vegetation recovery, combining both fieldwork and remote sensing data and considering varying degrees of fire severity and their effects on different land covers. The analysis aims to explore local successional dynamics within vegetation cover, based on sampling plots, and determine whether the results indicate broader landscape compositional shifts. Additionally, we apply an experimental framework based on the supervised classification of openly available satellite imagery to quantify and verify fire-induced changes on a larger spatial scale to enable the comparison of results obtained through the different approaches.
2. Materials and Methods
2.1. Study Area
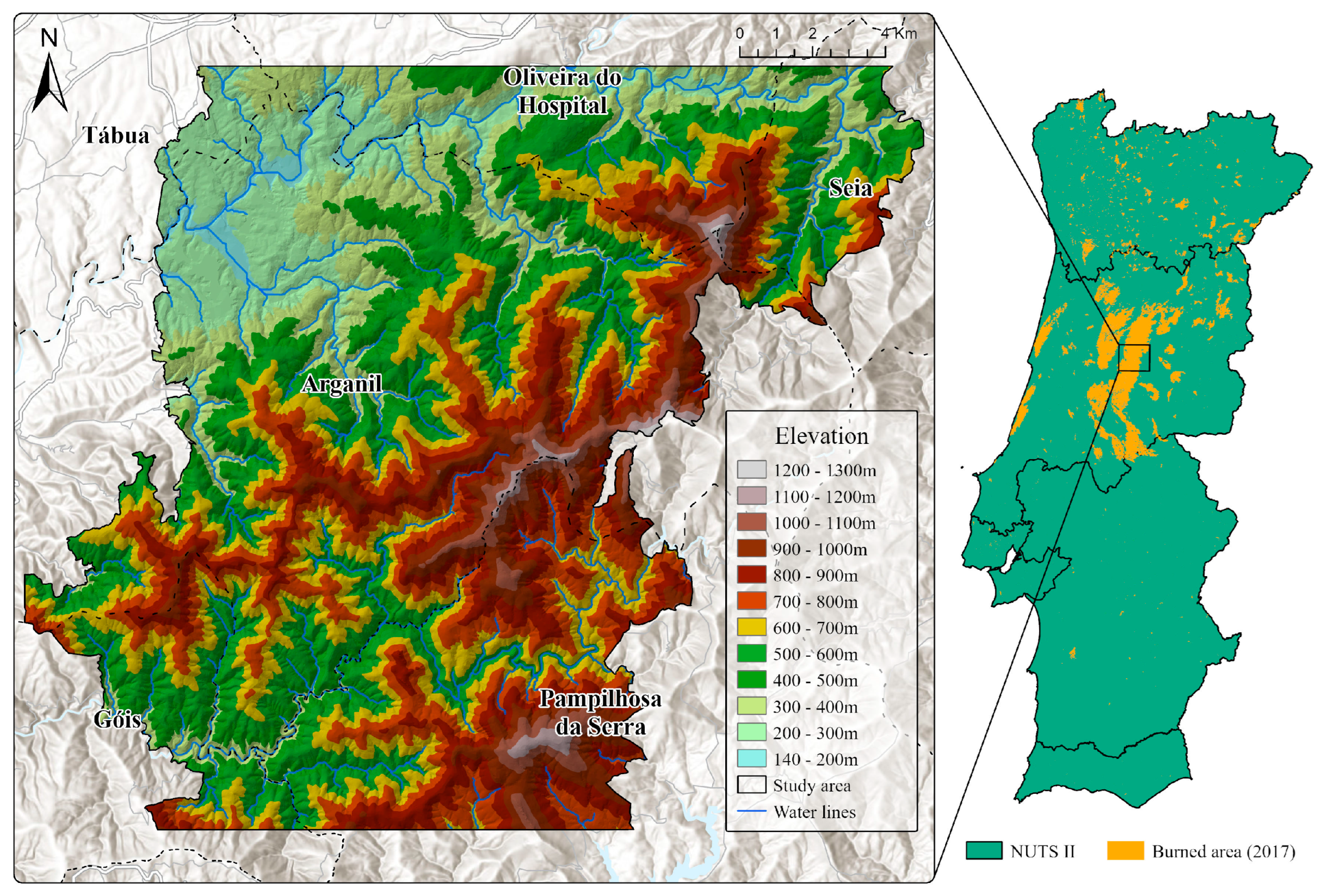
The study area, covering a total of approximately 334 km2, is located inland in central Portugal, corresponding to a section of the Serra do Açor mountain range that was significantly affected by a wildfire on the 15th of October 2017 (Figure 1). The region features a pronounced altitudinal gradient (170–1277 m), with about 75% of the area exhibiting slopes steeper than 15°. In terms of lithology, the area is largely composed of metamorphic rocks (schist), with granitic substrates being less represented. Although characterized by a Mediterranean climate, significant differences in precipitation within the study area can be observed thanks to the strong altitudinal gradient, ranging from approximately 1200 mm in the lower-altitude areas to 1700 mm in the higher-elevation sectors. Summers are hot and dry, with maximum temperatures exceeding 35 °C, average temperatures around 20 °C, and a pronounced lack of rainfall, especially in July and August. Winters are mild, with average temperatures around 10 °C, with the period between October and March recording the highest rainfall totals. Summers are hot and dry, with maximum temperatures exceeding 35 °C, average temperatures around 20 °C, and a pronounced lack of rainfall, especially in July and August.
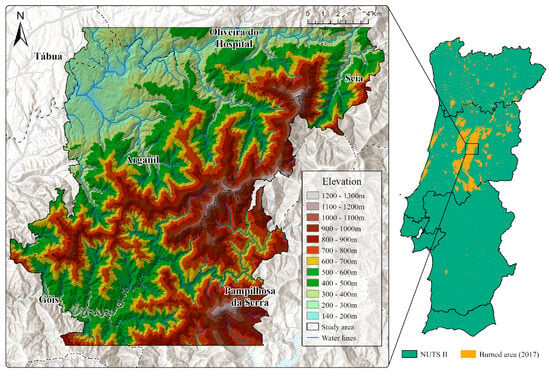
Figure 1.
Study area location and topography.
2.2. Field Sampling
Two Sentinel-2 multispectral images were acquired from the Copernicus Browser platform, captured on 2 October and 22 October 2017, corresponding to pre- and post-fire conditions. These images, preprocessed at level 2A, with a maximum of 10% cloud cover, provided the basis for assessing burned areas and fire severity. The identification of burned areas relied on the calculation of the normalized burn ratio (NBR), while fire severity was estimated through the delta normalized burn ratio (dNBR) (Table 1), following the classification thresholds established by Key and Benson [64] and also adopted by the European Forest Fire Information Service (EFFIS) (Table 2).

Table 1.
Spectral indices used.
Table 2.
dNBR classification parameters.
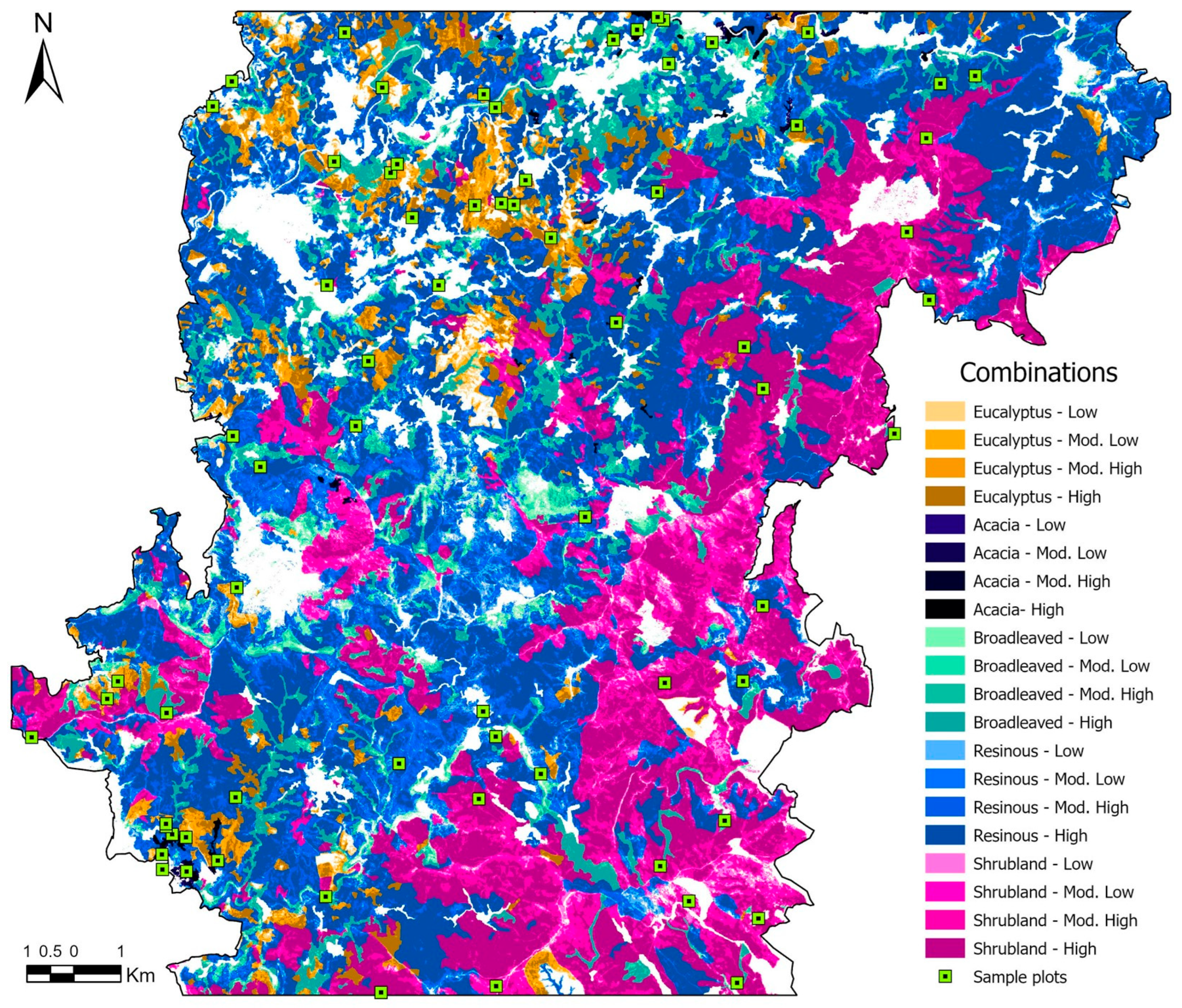
To explore the relationship between fire severity and its effects on vegetation recovery, the sampling design considered both fire severity classes and pre-fire land cover (Figure 2). In 2015, pine forests and shrublands covered the largest areas in the study area, while deciduous broad-leaved forests and eucalyptus forests were less prevalent. Acacia stands at that particular time only occupied very small areas. The spatial intersection of these five vegetation classes with the four fire severity levels was computed, resulting in 20 unique combinations for analysis. These were the basis for a stratified sampling scheme, with a minimum of three plots to be assigned per combination (Table 3). This approach allowed the inclusion of land covers of particular ecological or management interest, such as invasive acacia stands, which, despite being spatially less representative, are important for analysis due to their potentially distinct successional dynamics. Additionally, sampling efforts were reinforced for high-severity conditions within the three land covers with higher spatial representativeness (pine, broad-leaved deciduous forests and shrublands). In total, 70 sampling plots of 100 m2 (10 × 10 m) were distributed randomly across the study area (Figure 2).
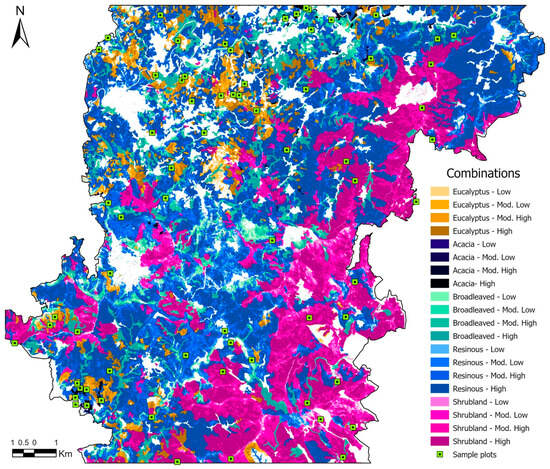
Figure 2.
Distribution of sampling plots across landcover and severity classes.
Table 3.
Stratified sampling distribution.
Field data collection took place in August 2023 and included the field plot’s physical attributes (altitude, slope, topographic position, and percentage of rock outcrops) and vegetation attributes, namely, structural and floristic parameters. Structural data included physiognomy, total cover and canopy height, while floristic data captured species cover and diversity. The collected data were organized in tabular format and processed using IBM SPSS (Version 29.0.2) statistical software to detect possible relationships between fire severity, land cover, and vegetation parameters.
2.3. Landscape Composition Change
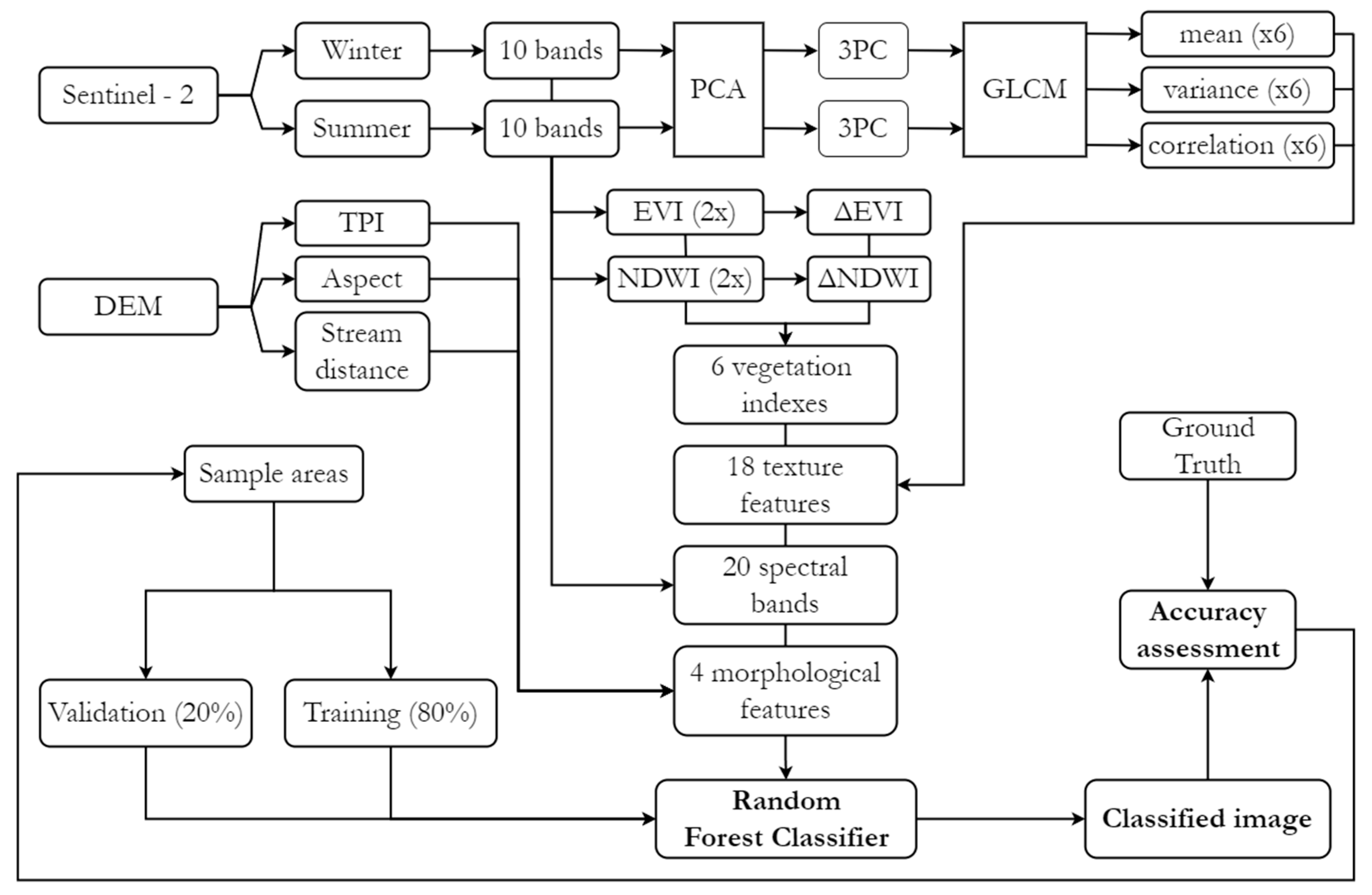
To detect and quantify landscape compositional shifts between the pre- and post-fire periods, a pixel-based supervised classification methodology was employed, focusing on the five land cover classes assessed during field sampling. Four Sentinel-2 images were selected following the same procedure described in Section 2.2, with the primary criterion being minimal cloud cover. The selected images corresponded to the winter and summer seasons for the years 2017 (pre-fire) and 2023/2024 (post-fire/present), and were captured on the following dates: 18 March 2017, 24 July 2017, 23 July 2023, and 3 February 2024. A spatial resolution of 10 m was adopted for this exercise, requiring resampling for some layers. In addition to the raw spectral data, the enhanced vegetation index (EVI) and the normalized difference water index (NDWI) were calculated for each acquisition, as well as their inter-seasonal variation (Δ) (Table 4), to highlight phenological characteristics and discard their influence on vegetation recovery analysis.
Table 4.
Spectral indices used.
Principal component analysis (PCA) was applied to each of the 10-band multispectral Sentinel-2 datasets, with the first three principal components being chosen for the grey level co-occurrence matrix (GLCM) analysis, as they captured around 97% of spectral variance across all images. This analysis, performed using ESA’s open-access Sentinel Application Platform (SNAP), statistically examines image texture based on the spatial relationships between pixels. The mean, variance, and correlation metrics (Table 4) were computed using probabilistic quantization at 32 levels using a 9 × 9 moving window, applied in four directions (0°, 45°, 90°, and 135°). Additionally, physical variables were included, namely, the digital elevation model (DEM), the topographic position index (TPI), the aspect (slope direction), and the Euclidean distance to streams (Table 4). In total, 48 independent variables were considered for each timeframe. As these layers had different value scales, normalization was applied to a 0–1 range. Finally, the variables were merged into separate multiband rasters, each containing 48 bands.
For the supervised classification, training samples were collected separately for each timeframe, as significant spectral changes were expected due to fire-induced changes in ecological conditions and successional stages. Given the relatively low spatial resolution of Sentinel-2 imagery, this process was primarily based on publicly accessible high-resolution imagery from Google Earth Pro, supplemented by Google Maps Street View captures when spatially and temporally available. Both platforms provide access to archived data, including pre-fire imagery, which would otherwise be difficult to obtain through open-access sources. Sampling included the five previously mentioned land cover classes, along with a background class encompassing all other land cover types (Table 5). Sample areas were delineated in Google Earth Pro, avoiding transitional zones between covers to minimize the inclusion of mixed pixels, which could arise due to the 10 m spatial resolution of the independent variables’ raster data. Additionally, 15 post-classification validation points were distributed manually for each class, relying on prior fieldwork knowledge or Street View captures, to ensure their accuracy.
Table 5.
Supervised classification sample areas and pixel count.
For the supervised classification (Figure 3), the random forest algorithm was employed because of its ensemble learning approach and non-parametric nature, which allow it to handle complex datasets effectively. Additionally, it provides variable importance estimates through metrics such as mean decrease Gini (MDG) and mean decrease accuracy (MDA), distinguishing it from many alternative methods. To implement this, the libraries “sf”, “terra”, “caret”, “randomForest”, and “ggplot2” were loaded in RStudio (version 2024.04.2). Sample areas were stratified into training and validation subsets at an 80/20 ratio, and the mean pixel values from the multiband raster intersecting each sample polygon were calculated. This step reduced the computational load and mitigated spatial autocorrelation issues, which could otherwise lead the algorithm to overestimate the relevance of terrain-related variables (e.g., DEM, TPI) by treating pixels within the same polygon as independent observations.
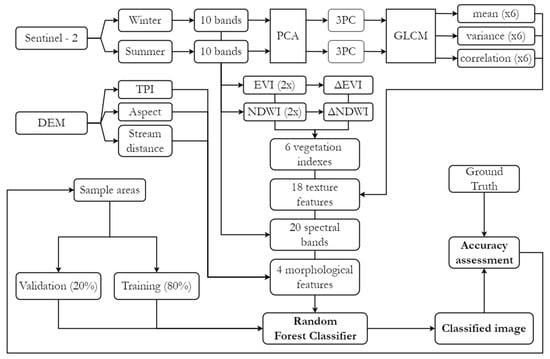
Figure 3.
Supervised classification workflow.
For model hyperparameters, commonly used default values were applied: ntree = 500, mtry = √48 (number of variables), and nodesize = 1. Once satisfactory performance levels were achieved in the validation subsets through adjustments to the sample areas, classification was extended to the entire multiband raster, and the results were exported. To reduce the salt-and-pepper effect, which is common in pixel-based approaches, clusters of three or fewer pixels were removed and reassigned to the classification of the largest neighboring area (Sieve). Finally, a confusion matrix was generated to compare the classification results with their corresponding validation points, assessing accuracy using metrics such as the Kappa index.
3. Results
Post-fire vegetation recovery assessed across 70 field plots revealed a pronounced shrub encroachment in areas occupied by native communities (shrubs, pine and broad-leaved deciduous forests), with varying structures, physiognomies, and compositions. The dominance of scrub physiognomy on recovery is only absent when exotics prevail. In the field, shrublands were identified 30 times as the predominant cover, a higher number than the 15 plots recorded in the pre-fire stage. The native tree cover, dominated by pine or deciduous broad-leaved species, despite being the most frequently sampled cover in the pre-fire stage, is scarcely represented, with only 10 plots still exhibiting such cover type. Conversely, plots with exotic trees (Acacia sp., Eucalyptus sp.) increased in the post-fire stage.
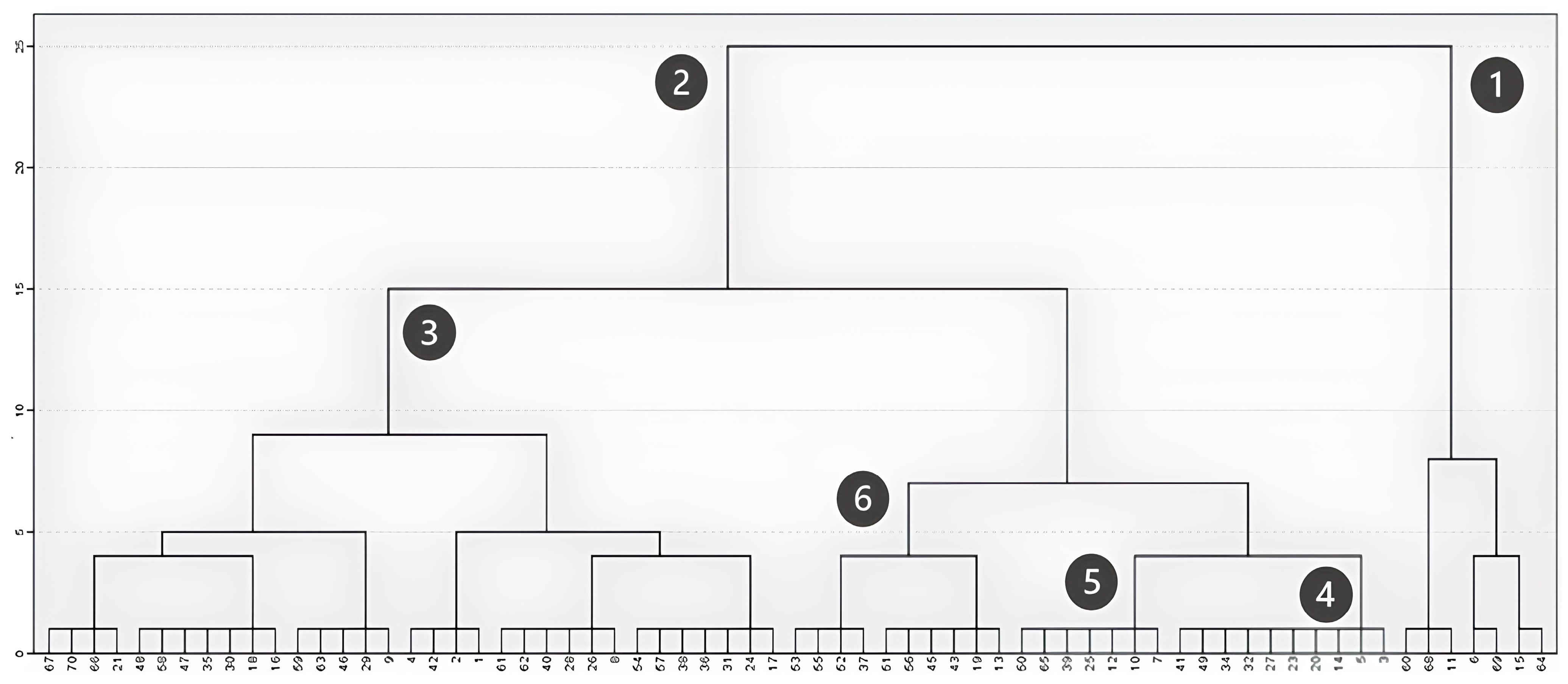
Hierarchical clustering, based on the between-group linkage method and Euclidean distance, further highlighted these successional dynamics. When considering the maximum vegetation height in field plots (Figure 4), the analysis distinctly identified two main groups: one corresponding to plots with large trees exceeding 12 m (Figure 4, Cluster 1) and another including plots where vegetation remains under 10 m (Figure 4, Cluster 2). Cluster 1 includes both plots less affected by the fire, allowing the maintenance of sparse trees, and acacia-dominated plots that fully recovered after the fire. Within the second group, three subdivisions emerged: plots dominated by shrubs with a maximum height between 1 and 6 m (Figure 4, Cluster 3); transitional plots with sparse tree cover and a maximum height around 7 m (Figure 4, Cluster 4); and plots predominantly composed of individuals between 8 and 10 m, mainly acacia individuals (Figure 4, Cluster 5) and eucalyptus (Figure 4, Cluster 6).
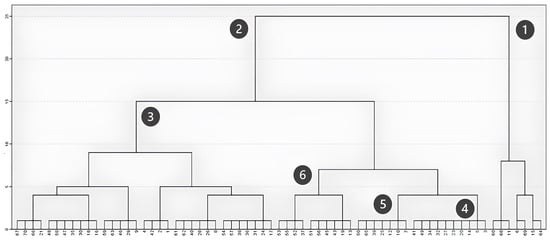
Figure 4.
Clustering based on maximum height (m).
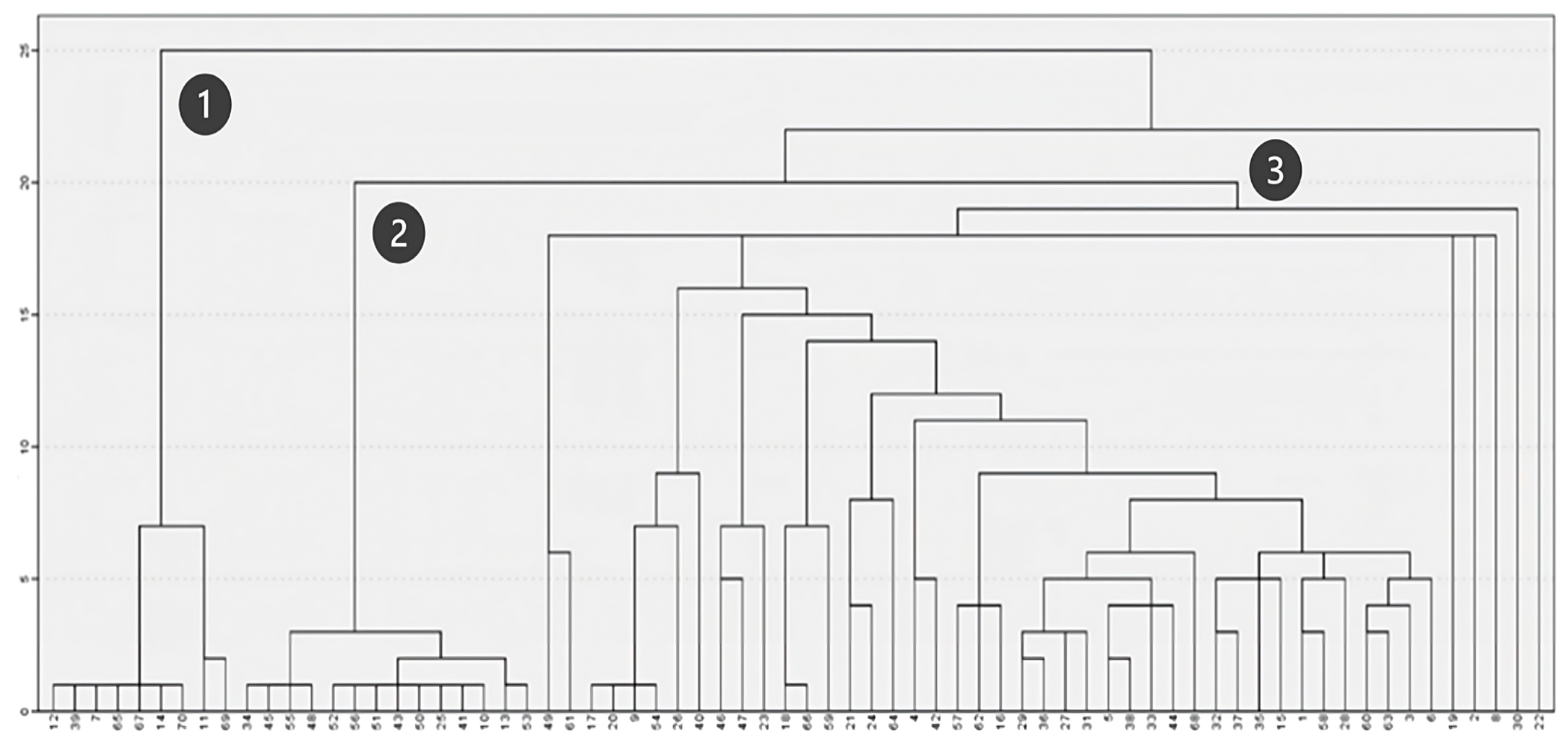
When analyzing floristic composition (Figure 5), three main groups of plots were distinguished. Two of these groups (Figure 5, Clusters 1 and 2) consist of plots with more homogeneous floristic compositions, while the third group (Figure 5, Cluster 3) includes plots with diverse species compositions, leading to the formation of multiple subgroups on the right side of the dendrogram. Accordingly, Clusters 1 and 2 correspond to plots dominated by acacia and eucalyptus, respectively, whereas Cluster 3 primarily includes plots associated with native shrubland communities.
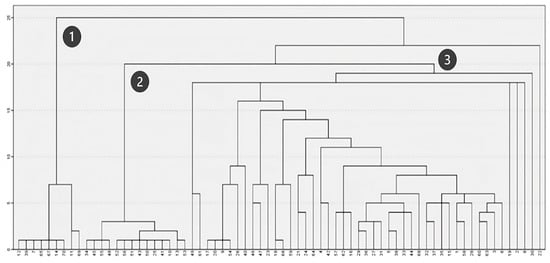
Figure 5.
Clustering based on floristic composition.
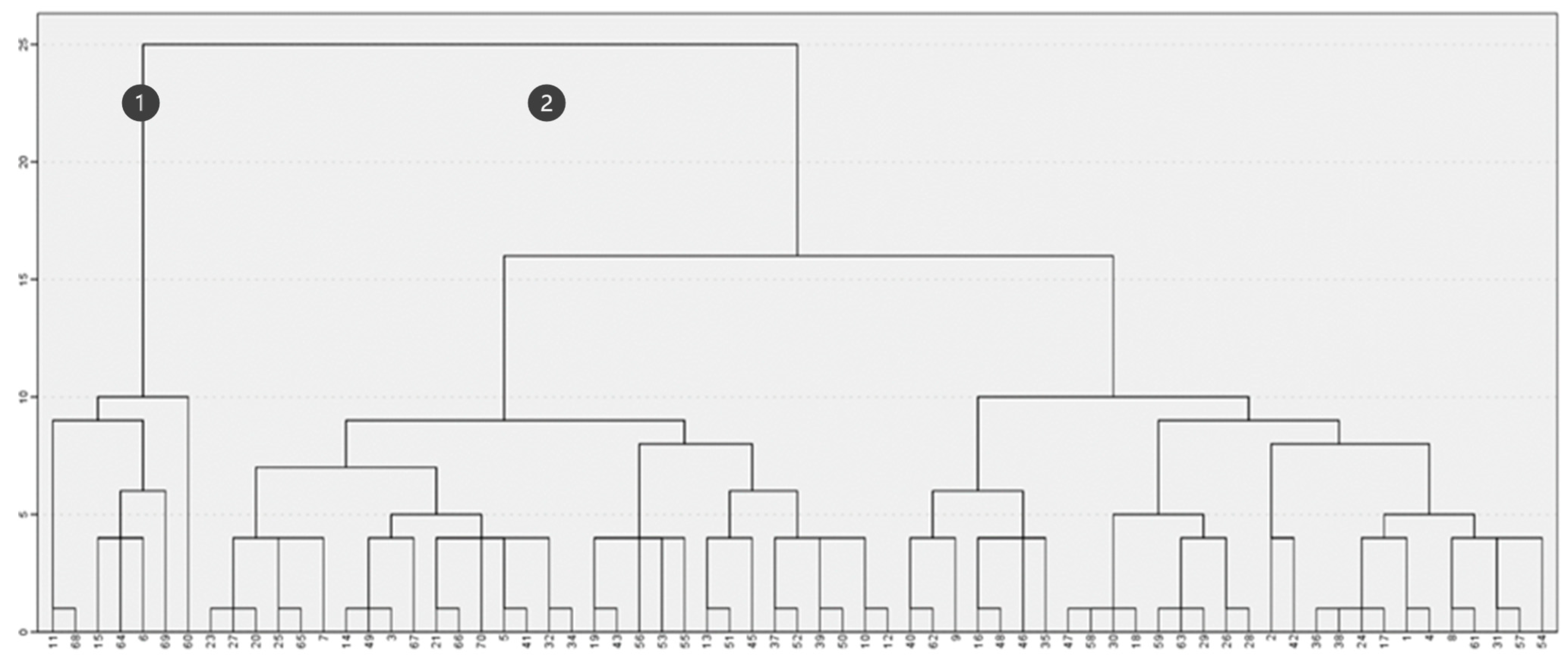
In contrast, when considering both fire severity and maximum height (Figure 6), the clustering created various subsets demonstrating the lack of common features. The two main groups (Figure 6, Clusters 1 and 2) are primarily formed based on maximum vegetation height, closely resembling the results obtained in Figure 4. In fact, intragroup association does not reflect similarities in burn severity, indicating that this factor had a limited influence on post-fire maximum vegetation height differentiation.
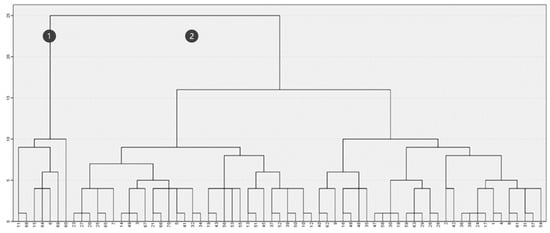
Figure 6.
Clustering based on maximum height (m) and fire severity.
Similarly, applying a one-way ANOVA test (analysis of variance) confirmed that there is no statistically significant difference in maximum vegetation height across plots with varying burn severity levels. Likewise, no significant correlations were identified between burn severity and physiognomic parameters (Table 6) or other variables, such as altitude, percentage of rock outcrop, and topographic position of the plots.
Table 6.
Correlation analysis (Spearman’s rho).
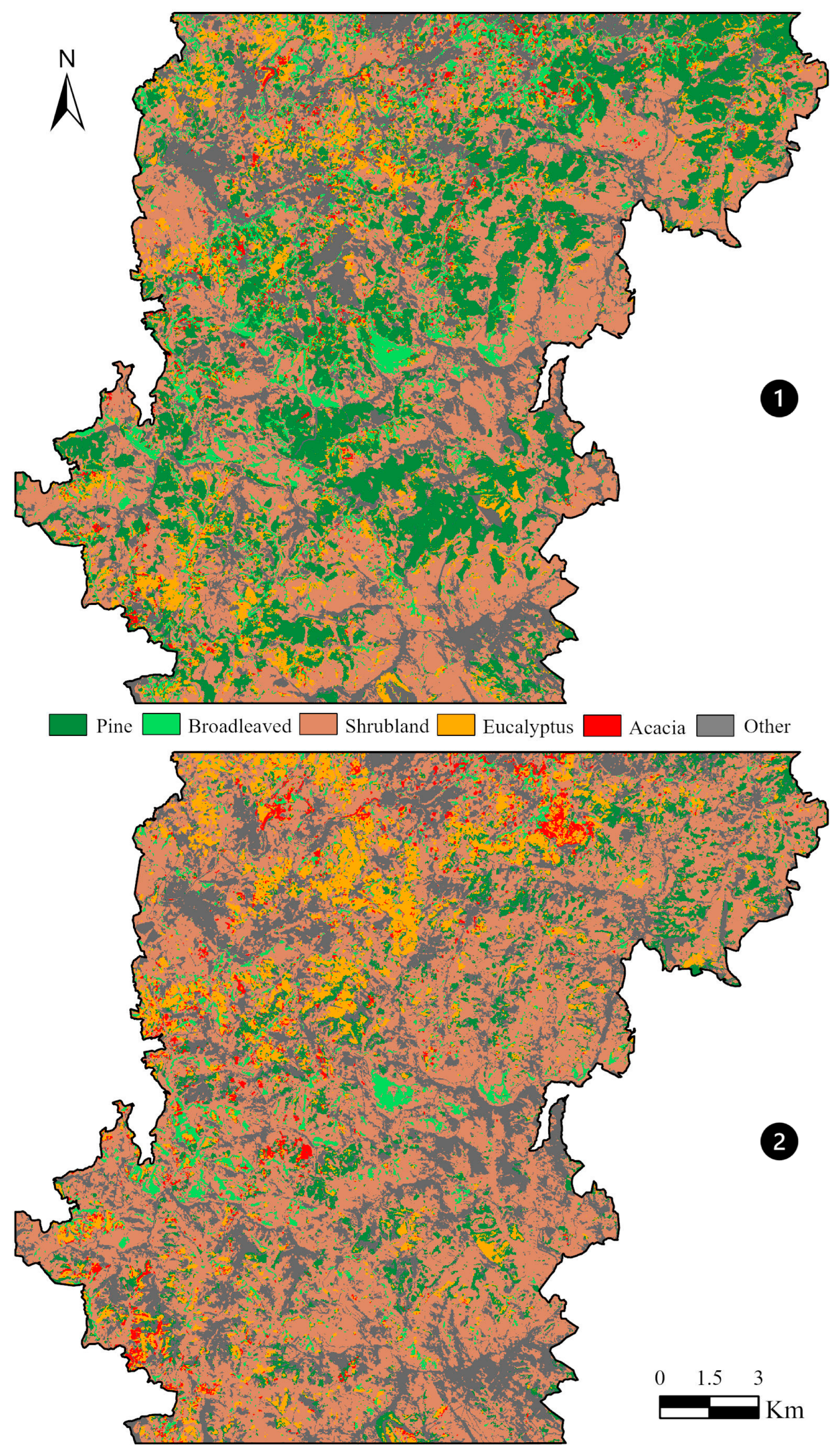
The results obtained for changes in vegetation comparing pre-fire (Figure 7—①) and post-fire (Figure 7—②) conditions for the entire study area, based on supervised classification, suggested similar shifts as detected based on plot analysis (Table 7). The expected increase in the area occupied by shrublands, which expanded by approximately 3000 hectares, occurred following a reduction in areas covered by native tree species, with pine stands being the most severely affected, experiencing a loss of nearly 4800 hectares, compared to a 1200-hectare reduction in broad-leaved deciduous forests (Table 7). Regarding exotic tree species, increases of approximately 300 hectares were recorded for both eucalyptus and acacia stands. This corresponds to a moderate expansion of eucalyptus-covered areas (+11%) and a substantial increase in acacia-dominated stands (+51%), with the latter showing the highest relative growth among all land cover types analyzed.
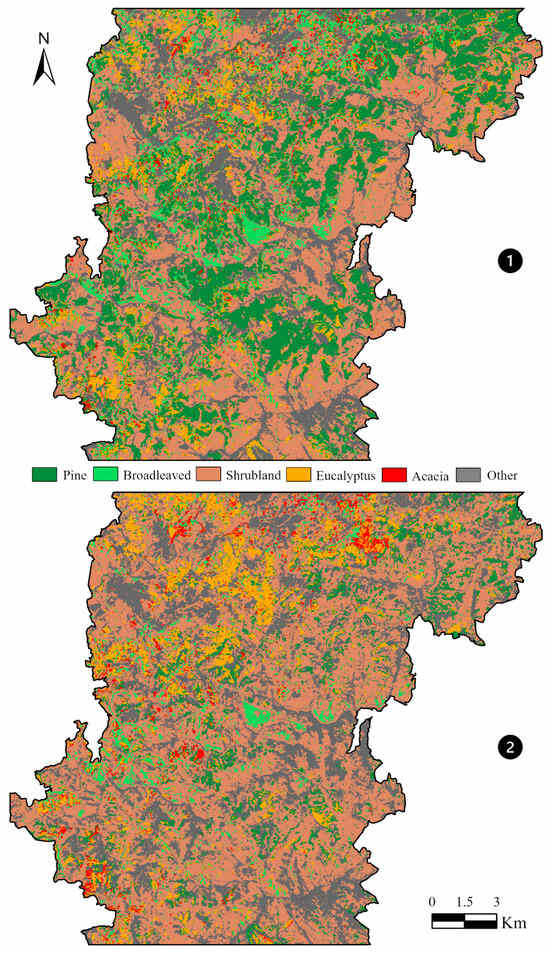
Figure 7.
Pre-fire ① and post-fire ②supervised classification results.
Table 7.
Vegetation cover change.
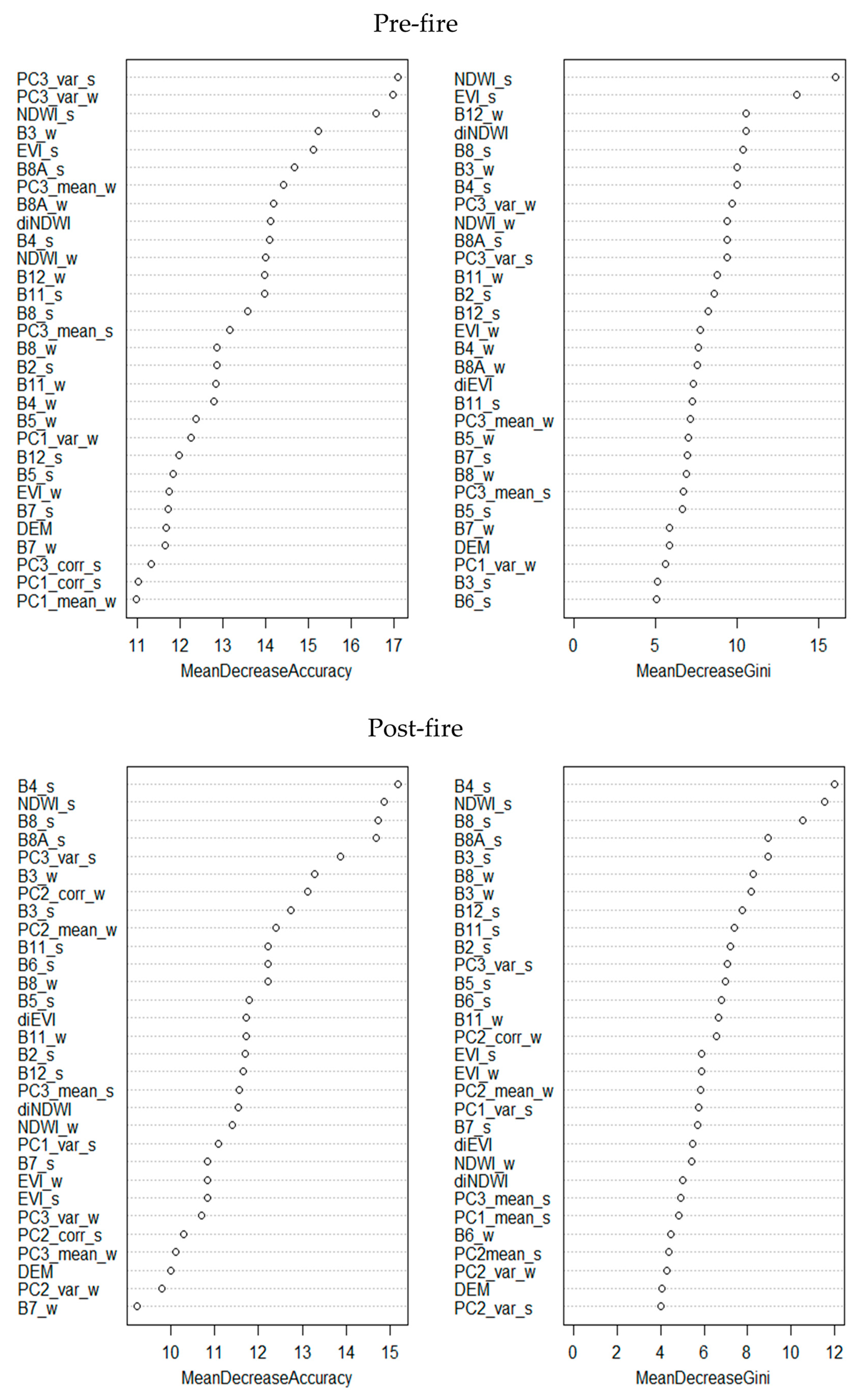
Both supervised classifications demonstrated strong performance, achieving Kappa values exceeding 0.90 in both the random forest test subset and the validation points (Table 8). The models exhibited greater ease in excluding rather than assigning results, as indicated by consistently high specificity values and comparatively greater variability in sensitivity, particularly in the post-fire timeframe. The contribution of independent variables to classification performance (Appendix A) varied primarily between timeframes but also between metrics, probably due to spectral shifts caused by fire. Of the vegetation indices, summer NDWI consistently ranked among the three most influential variables. Regarding the 10 satellite bands, B8 and/or B8A (summer), B3 (winter), and B4 (summer) exerted the greatest influence across both models. In the case of GLCM indices, statistical measures derived from the third principal component (PC) held the highest relevance, particularly the variance index (summer/winter) [67]. Topographical variables, however, showed minimal relevance in all cases.
Table 8.
Model performance metrics.
4. Discussion and Conclusions
The assessment of vegetation recovery across different plots did not reveal a clear relationship with fire severity, as no statistically significant differences were found between severity levels considering floristic composition or physiognomic attributes (Figure 6, Table 6). Our results are consistent with the findings obtained by several authors [30,31,36] supporting the idea that there is no relationship between fire severity and vegetation recovery, a process that is primarily determined by the ecosystem type and pre-fire species composition.
As expected, post-fire vegetation attributes, namely, cover and height, were primarily determined by local abiotic factors, such as topographic and climate conditions [30]. This is evident in Table 6. This shows a significant negative correlation between the physical site characteristics (altitude, topographic position, and rock outcrop cover) and physiognomic traits, such as average height and maximum height, both influenced by floristic composition. This relationship is evident in most plots, where maximum plant height 6 years after the fire remained below 10 m and the cover was dominated by native species that regenerate mainly through vegetative resprouting [35,68].
These plots exhibited distinct floristic and physiognomic characteristics from those dominated by fast-growing exotic species (Figure 5). In the latter, when associated with more favorable edaphoclimatic conditions (e.g., valley bottoms), physiognomic differences became even more pronounced. This is the case for plots dominated by Acacia dealbata, where height even exceeds 12 m, confirming the high capacity of such species to recover after fire. These results are in line with those obtained by Nunes et al. [69], who demonstrated a competitive advantage for A. dealbata, even when compared to species such as Eucalyptus globulus and Pinus pinaster.
Other plots also exhibit trees surpassing 12 m in height, but these correspond to situations where tree structure was preserved and regeneration from branches or trunks occurred. This was found on plots with cork oak (Quercus suber), chestnut (Castanea sativa) and eucalyptus (Eucalyptus sp.), cases that did not always align with low-severity fire classifications. Within the native vegetation cover, the dominance of shrublands in the landscape aligns with the expected secondary successional dynamics taking place six years after fire disturbance, with most previous tree stands having reverted to shrub-dominated cover. Our results are in line with those found by [28] in areas affected by high-severity fires, which experienced the most pronounced expansion of dense shrubland in southern Spain. As noted by [29,70], certain Mediterranean species, such as Cistus ladanifer, are particularly flammable and combustible, and their increased prevalence following wildfires can alter the fire regime itself, intensifying recurrences.
The presence of native tree species was recorded in 34 plots (49%) for maritime pine (Pinus pinaster) and in 25 plots (36%) for broad-leaved deciduous species (Quercus, Castanea, or Salix). The long-term trajectory of succession and potential climax community development will once again depend on local edaphoclimatic constraints, with shrub-dominated communities more likely to persist in higher altitude areas, although with some floristic adjustments. In contrast, communities dominated by acacia species (Acacia dealbata; A. melanoxylon) exhibit distinct post-fire dynamics. Shrubland communities largely fail to establish thanks to the rapid and efficient regeneration of exotic species, which outcompete native vegetation for space and resources [51,55] leading to low floristic diversity (Figure 5). The 10 plots dominated by acacia were nearly monospecific, with acacias accounting for 80–100% of cover in the plot.
Eucalyptus-dominated plots, while similarly floristically homogeneous, were primarily associated with commercial plantations rather than natural succession. While the attributes of acacia stands are primarily driven by the species’ intrinsic ecological characteristics, the low floristic diversity in eucalyptus stands is primarily a consequence of plantation management practices, including fuel load reduction, which may have influenced some field observations.
When propagules of these exotic species establish within native vegetation, mixed-cover landscapes emerge. Over time, and particularly under recurring disturbances, these species can significantly alter vegetation composition and structure, potentially reshaping ecosystem dynamics and diminishing biodiversity [71,72]. Within the surveyed plots, vegetation recovery in areas dominated by eucalyptus registered greater floristic diversity than those dominated by acacia species, which actively suppressed native species. Furthermore, acacias demonstrated a remarkable ability to colonize biophysically unfavorable areas, having been found on steep rocky slopes with gradients close to 70% (35°) and on ridge tops with shallow soils and large rocky outcrops, as also noted in the literature [54]. This ability to establish in diverse ecological conditions, combined with an effective response to fire, rapid growth, high stand density, and the accumulation of large amounts of litter, makes the expansion of these communities both a consequence and a driver of fire disturbance, as also suggested by Brooks et al. [53].
When it comes to changes in cover between the pre-fire and current conditions, classification results tend to align with the successional dynamics observed at a plot level. However, several limitations must be considered. Classifying the study area across two timeframes inevitably introduced uncertainties associated with variations in surface illumination, atmospheric conditions, sensor calibration, and vegetation changes [73]. Due to these factors, even under stable conditions, that is, without disturbance, the supervised classification of multispectral images from different years can yield distinct results. In this study, classification was performed on two datasets acquired not only at different time points but also under markedly different vegetation conditions. Consequently, distinct training areas had to be defined for each timeframe, leading to some inconsistencies, even in areas where conditions appeared relatively stable, due to slight differences in parameterization. Additionally, the low spatial resolution (10–20 m) further complicated classification by introducing mixed pixels, which can lead to misclassifications and distortions in land cover estimates [74].
Although both cover maps maintained the same classification schema, the vegetative characteristics of the included communities varied profoundly between the two timeframes, and methodological challenges emerged accordingly. Pre-fire landscapes consisted of compositionally and structurally complex communities with high intraclass heterogeneity, varying stand densities, and frequent mixed stands of species like maritime pine (Pinus pinaster) and eucalyptus (Eucalyptus sp.), which were difficult to differentiate.
The occurrence of fire reset most vegetation communities to an early successional stage, reducing intraclass heterogeneity while increasing interclass homogeneity. This shift made spectrally distinguishing between certain land covers more challenging, particularly pure shrublands and shrublands with tree species recovery (e.g., Pinus pinaster). Furthermore, the relatively low spatial resolution of the imagery (10–20 m) hindered the detection of areas where tree saplings were present but not yet dominant, a common feature of native vegetation recovery, as frequently observed in field plots. Additionally, mixed pixels near class boundaries introduced classification uncertainties, especially for species whose lateral expansion patterns could be misinterpreted due to parameter variations in the classification process.
Validation of past land cover conditions (e.g., 2017) is notably constrained by the availability of remote sensing data, which varies by study area and becomes increasingly scarce for older timeframes. Current sampling, referring to the present timeframe, could benefit from broader field data integration, given that its collection framework is adjusted to match the dataset’s pixel size and minimum cover area detectability. In this study, such an adjustment was not made, thus limiting the use of field plots for classification validation. Future improvements should integrate cross-validation into the learning stage of the classifier to reduce accuracy estimation biases arising from the fixed partitioning of training and test subsets. Furthermore, calibrating algorithm parameters should be explored to assess whether adjustments improve classification accuracy.
Moreover, to gain a deeper understanding of post-fire recovery processes, more variables need to be integrated into a comprehensive model, including climate variability [75], the effects of fire recurrence [28], and the implementation of management techniques such as planting seeds or seedlings [76], among other factors. Despite these challenges, we consider our approach to be robust and a strong foundation for future work, with compositional classifications expected to become more reliable as vegetation communities mature and structural complexity is restored.
The restoration of burned forest ecosystems is essential to mitigating the adverse effects of wildfires on ecological, economic, and social components [61,76]. Such an approach, based on remote sensing products and fieldwork validation, is key to developing a better understanding of vegetation recovery dynamics, and thus facilitating the creation and implementation of robust, management-linked solutions.
Author Contributions
Conceptualization, N.W., A.F. and A.N.N.; methodology, N.W. and A.F.; software, N.W. and A.F.; formal analysis, N.W., A.F. and A.N.N.; Validation, N.W. and A.F; writing—original draft preparation, N.W., A.F. and A.N.N.; visualization, N.W., A.F. and A.N.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received support from the Centre of Studies in Geography and Spatial Planning (CEGOT), funded by national funds through the Portuguese Foundation for Science and Technology (FCT) under the reference UIDB/04084/2025.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
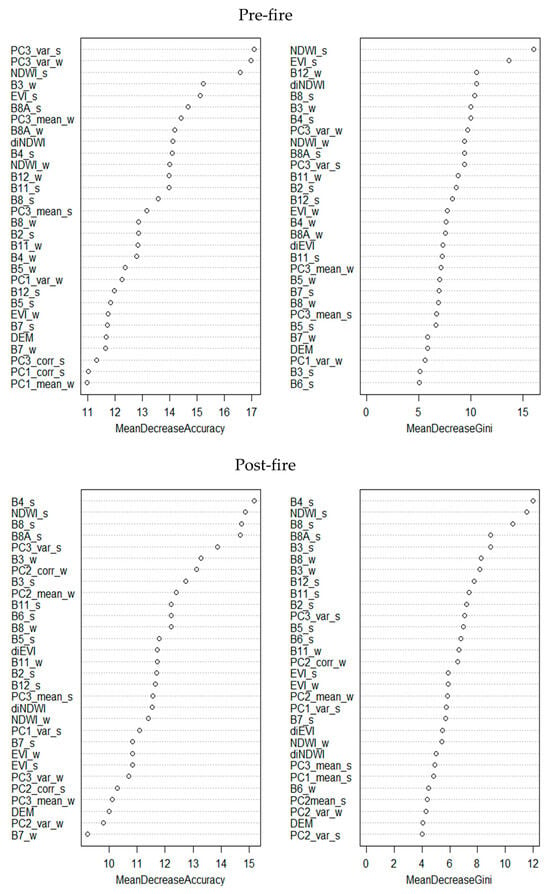
Figure A1.
Variable contribution estimation for each model.
Figure A1.
Variable contribution estimation for each model.
References
- European Commission; Joint Research Centre. Forest Fires in Europe, Middle East and North Africa 2017; Publications Office: Luxembourg, 2018. [Google Scholar]
- Turco, M.; Llasat, M.C.; von Hardenberg, J.; Provenzale, A. Impact of Climate Variability on Summer Fires in a Mediterranean Environment (Northeastern Iberian Peninsula). Clim. Chang. 2013, 116, 665–678. [Google Scholar] [CrossRef]
- Pausas, J.G. Changes in Fire and Climate in the Eastern Iberian Peninsula (Mediterranean Basin). Clim. Chang. 2004, 63, 337–350. [Google Scholar] [CrossRef]
- Rosavec, R.; Barčić, D.; Španjol, Ž.; Oršanić, M.; Dubravac, T.; Antonović, A. Flammability and Combustibility of Two Mediterranean Species in Relation to Forest Fires in Croatia. Forests 2022, 13, 1266. [Google Scholar] [CrossRef]
- Dimitrakopoulos, A.; Papaioannou, K. Flammability Assessment of Mediterranean Forest Fuels. Fire Technol. 2001, 37, 143–152. [Google Scholar] [CrossRef]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2011; ISBN 978-1-107-37699-1. [Google Scholar]
- Trigo, R.M.; Pereira, J.M.C.; Pereira, M.G.; Mota, B.; Calado, T.J.; Dacamara, C.C.; Santo, F.E. Atmospheric Conditions Associated with the Exceptional Fire Season of 2003 in Portugal. Int. J. Climatol. 2006, 26, 1741–1757. [Google Scholar] [CrossRef]
- García-Herrera, R.; Hernández, E.; Barriopedro, D.; Paredes, D.; Trigo, R.M.; Trigo, I.F.; Mendes, M.A. The Outstanding 2004/05 Drought in the Iberian Peninsula: Associated Atmospheric Circulation. J. Hydrometeorol. 2007, 8, 483–498. [Google Scholar] [CrossRef]
- Ramos, A.M.; Russo, A.; DaCamara, C.C.; Nunes, S.; Sousa, P.; Soares, P.M.M.; Lima, M.M.; Hurduc, A.; Trigo, R.M. The Compound Event That Triggered the Destructive Fires of October 2017 in Portugal. iScience 2023, 26, 106141. [Google Scholar] [CrossRef]
- Oliveira, S.; Pereira, J.M.C.; San-Miguel-Ayanz, J.; Lourenço, L. Exploring the Spatial Patterns of Fire Density in Southern Europe Using Geographically Weighted Regression. Appl. Geogr. 2014, 51, 143–157. [Google Scholar] [CrossRef]
- Pausas, J.; Keeley, J. A Burning Story: The Role of Fire in the History of Life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef]
- San-Miguel-Ayanz, J.; Moreno, J.M.; Camia, A. Analysis of Large Fires in European Mediterranean Landscapes: Lessons Learned and Perspectives. For. Ecol. Manag. 2013, 294, 11–22. [Google Scholar] [CrossRef]
- Mantero, G.; Morresi, D.; Marzano, R.; Motta, R.; Mladenoff, D.J.; Garbarino, M. The Influence of Land Abandonment on Forest Disturbance Regimes: A Global Review. Landsc. Ecol. 2020, 35, 2723–2744. [Google Scholar] [CrossRef]
- Moreno, M.V.; Conedera, M.; Chuvieco, E.; Pezzatti, G.B. Fire Regime Changes and Major Driving Forces in Spain from 1968 to 2010. Environ. Sci. Policy 2014, 37, 11–22. [Google Scholar] [CrossRef]
- Pereira, P.; Rein, G.; Martin, D. Past and Present Post-Fire Environments. Sci. Total Environ. 2016, 573, 1275–1277. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.; Catry, F.; Rego, F.; Bação, F. Size-Dependent Pattern of Wildfire Ignitions in Portugal: When Do Ignitions Turn into Big Fires? Landsc. Ecol. 2010, 25, 1405–1417. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Sousa, A.M.O.; Gonçalves, A.C.; Sousa, A.M.O. The Fire in the Mediterranean Region: A Case Study of Forest Fires in Portugal. In Mediterranean Identities—Environment, Society, Culture; IntechOpen: Rijeka, Croatia, 2017; ISBN 978-953-51-3586-9. [Google Scholar]
- Maia, P.; Keizer, J.; Vasques, A.; Abrantes, N.; Roxo, L.; Fernandes, P.; Ferreira, A.; Moreira, F. Post-Fire Plant Diversity and Abundance in Pine and Eucalypt Stands in Portugal: Effects of Biogeography, Topography, Forest Type and Post-Fire Management. For. Ecol. Manag. 2014, 334, 154–162. [Google Scholar] [CrossRef]
- ICNF. Inventário Florestal Nacional 2015. Available online: https://www.icnf.pt/florestas/flestudosdocumentosestatisticasindicadores (accessed on 15 July 2024).
- Moreira, F.; Viedma, O.; Arianoutsou, M.; Curt, T.; Koutsias, N.; Rigolot, E.; Barbati, A.; Corona, P.; Vaz, P.; Xanthopoulos, G.; et al. Landscape—Wildfire Interactions in Southern Europe: Implications for Landscape Management. J. Environ. Manag. 2011, 92, 2389–2402. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Meireles, C.I.R.; Pinto Gomes, C.J.; Almeida Ribeiro, N.M.C. Historical Development of the Portuguese Forest: The Introduction of Invasive Species. Forests 2019, 10, 974. [Google Scholar] [CrossRef]
- Catry, F.; Rego, F.C.; Moreira, F.; Bacao, F. Characterizing and Modelling the Spatial Patterns of Wildfire Ignitions in Portugal: Fire Initiation and Resulting Burned Area. WIT Trans. Ecol. Environ. 2008, 119, 213–221. [Google Scholar]
- ICNF Análise Das Causas Dos Incêndios Florestais, 2003–2013. 2014. Available online: https://www.icnf.pt/florestas/gfr/gfrgestaoinformacao/grfrelatorios/dfciprevencaoecausas (accessed on 15 July 2024).
- Catry, F.; Rego, F.; Bação, F.; Moreira, F. Modeling and Mapping Wildfire Ignition Risk in Portugal. Int. J. Wildland Fire 2009, 18, 921–931. [Google Scholar] [CrossRef]
- Piñol, J.; Terradas, J.; Lloret, F. Climate Warming, Wildfire Hazard, and Wildfire Occurrence in Coastal Eastern Spain. Clim. Chang. 1998, 38, 345–357. [Google Scholar] [CrossRef]
- Pausas, J.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are Wildfires a Disaster in the Mediterranean Basin?—A Review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Niklas, K.J.; Hammond, S.T. Biophysical Effects on Plant Competition and Coexistence. Funct. Ecol. 2013, 27, 854–864. [Google Scholar] [CrossRef]
- Ortega, M.; Lora, Á.; Yocom, L.; Zumaquero, R.; Molina, J.R. Effects of Fire Recurrence and Severity on Mediterranean Vegetation Dynamics: Implications for Structure and Composition in Southern Spain. Sci. Total Environ. 2025, 961, 178392. [Google Scholar] [CrossRef]
- Gallego, J.C.A.; Caro, J.G.; Campos, V.H.; Lobón, N.C. Effect of Leaf Litter from Cistus ladanifer L. on the Germination and Growth of Accompanying Shrubland Species. Plants 2020, 9, 593. [Google Scholar] [CrossRef]
- Spatola, M.F.; Borghetti, M.; Nolè, A. Elucidating Factors Driving Post-Fire Vegetation Recovery in the Mediterranean Forests Using Landsat Spectral Metrics. Agric. For. Meteorol. 2023, 342, 109731. [Google Scholar] [CrossRef]
- Keeley, J. Fire Intensity, Fire Severity and Burn Severity: A Brief Review and Suggested Usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Fernández-Guisuraga, J.M.; Fernandes, P.M.; Tárrega, R.; Beltrán-Marcos, D.; Calvo, L. Vegetation Recovery Drivers at Short-Term after Fire Are Plant Community-Dependent in Mediterranean Burned Landscapes. For. Ecol. Manag. 2023, 539, 121034. [Google Scholar] [CrossRef]
- Moya, D.; González-De Vega, S.; García-Orenes, F.; Morugán-Coronado, A.; Arcenegui, V.; Mataix-Solera, J.; Lucas-Borja, M.E.; De las Heras, J. Temporal Characterisation of Soil-Plant Natural Recovery Related to Fire Severity in Burned Pinus halepensis Mill. Forests. Sci. Total Environ. 2018, 640–641, 42–51. [Google Scholar] [CrossRef]
- Blanco-Rodríguez, M.Á.; Ameztegui, A.; Gelabert, P.; Rodrigues, M.; Coll, L. Short-Term Recovery of Post-Fire Vegetation Is Primarily Limited by Drought in Mediterranean Forest Ecosystems. Fire Ecol. 2023, 19, 68. [Google Scholar] [CrossRef]
- Viana-Soto, A.; Aguado, I.; Salas, J.; García, M. Identifying Post-Fire Recovery Trajectories and Driving Factors Using Landsat Time Series in Fire-Prone Mediterranean Pine Forests. Remote Sens. 2020, 12, 1499. [Google Scholar] [CrossRef]
- Smith-Ramírez, C.; Castillo-Mandujano, J.; Becerra, P.; Sandoval, N.; Fuentes, R.; Allende, R.; Paz Acuña, M. Combining Remote Sensing and Field Data to Assess Recovery of the Chilean Mediterranean Vegetation after Fire: Effect of Time Elapsed and Burn Severity. For. Ecol. Manag. 2022, 503, 119800. [Google Scholar] [CrossRef]
- Viana-Soto, A.; Aguado, I.; Martínez, S. Assessment of Post-Fire Vegetation Recovery Using Fire Severity and Geographical Data in the Mediterranean Region (Spain). Environments 2017, 4, 90. [Google Scholar] [CrossRef]
- González-De Vega, S.; De Las Heras, J.; Moya, D. Post-Fire Regeneration and Diversity Response to Burn Severity in Pinus halepensis Mill. Forests. Forests 2018, 9, 299. [Google Scholar] [CrossRef]
- Meneses, B.M. Vegetation Recovery Patterns in Burned Areas Assessed with Landsat 8 OLI Imagery and Environmental Biophysical Data. Fire 2021, 4, 76. [Google Scholar] [CrossRef]
- Huerta, S.; Marcos, E.; Fernández-García, V.; Calvo, L. Resilience of Mediterranean Communities to Fire Depends on Burn Severity and Type of Ecosystem. Fire Ecol. 2022, 18, 28. [Google Scholar] [CrossRef]
- Arnan, X.; Quevedo, L.; Rodrigo, A. Forest Fire Occurrence Increases the Distribution of a Scarce Forest Type in the Mediterranean Basin. Acta Oecologica 2013, 46, 39–47. [Google Scholar] [CrossRef]
- Catry, F.; Pausas, J.; Moreira, F.; Fernandes, P.; Rego, F. Post-Fire Response Variability in Mediterranean Basin Tree Species in Portugal. Int. J. Wildland Fire 2013, 22, 919–932. [Google Scholar] [CrossRef]
- Viana-Soto, A.; Okujeni, A.; Pflugmacher, D.; García, M.; Aguado, I.; Hostert, P. Quantifying Post-Fire Shifts in Woody-Vegetation Cover Composition in Mediterranean Pine Forests Using Landsat Time Series and Regression-Based Unmixing. Remote Sens. Environ. 2022, 281, 113239. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J. Ecology of Sprouting in Woody Plants: The Persistence Niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant Functional Traits in Relation to Fire in Crown-Fire Ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef]
- Capitanio, R.; Carcaillet, C. Post-Fire Mediterranean Vegetation Dynamics and Diversity: A Discussion of Succession Models. For. Ecol. Manag. 2008, 255, 431–439. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Delgado-Baquerizo, M.; Muñoz-Rojas, M.; Plaza-Álvarez, P.A.; Gómez-Sanchez, M.E.; González-Romero, J.; Peña-Molina, E.; Moya, D.; de las Heras, J. Changes in Ecosystem Properties after Post-Fire Management Strategies in Wildfire-Affected Mediterranean Forests. J. Appl. Ecol. 2021, 58, 836–846. [Google Scholar] [CrossRef]
- Costa, J.; Silva, J.S.; Deus, E.; Pinho, S.; Pinto, J.F.; Borralho, N. The Genetics and Ecology of Post-Fire Eucalyptus globulus Recruitment in an Isolated Stand in Central Portugal. Forests 2022, 13, 680. [Google Scholar] [CrossRef]
- dos Santos, P.; Matias, H.; Deus, E.; Águas, A.; Silva, J.S. Fire Effects on Capsules and Encapsulated Seeds from Eucalyptus globulus in Portugal. Plant Ecol. 2015, 216, 1611–1621. [Google Scholar] [CrossRef]
- Cruz, O.; Riveiro, S.F.; Arán, D.; Bernal, J.; Casal, M.; Reyes, O. Germinative Behaviour of Acacia dealbata Link, Ailanthus altissima (Mill.) Swingle and Robinia pseudoacacia L. in Relation to Fire and Exploration of the Regenerative Niche of Native Species for the Control of Invaders. Glob. Ecol. Conserv. 2021, 31, e01811. [Google Scholar] [CrossRef]
- Raposo, M.A.M.; Pinto Gomes, C.J.; Nunes, L.J.R. Evaluation of Species Invasiveness: A Case Study with Acacia Dealbata Link. on the Slopes of Cabeça (Seia-Portugal). Sustainability 2021, 13, 11233. [Google Scholar] [CrossRef]
- Riveiro, S.F.; Cruz, Ó.; Casal, M.; Reyes, O. Fire and Seed Maturity Drive the Viability, Dormancy, and Germination of Two Invasive Species: Acacia longifolia (Andrews) Willd. and Acacia mearnsii De Wild. Ann. For. Sci. 2020, 77, 60. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of Invasive Alien Plants on Fire Regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef]
- Cueva, A.V. de la Case Studies of the Expansion of Acacia Dealbata in the Valley of the River Miño (Galicia, Spain). For. Syst. 2014, 23, 3–14. [Google Scholar] [CrossRef]
- Marchante, H.; Marchante, E.; Freitas, H. Invasion of the Portuguese Dune Ecosystems by the Exotic Species Acacia longifolia (Andrews) Willd.: Effects at the Community Level. In Plant Invasions: Ecological Threats and Management Solutions; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 75–85. [Google Scholar]
- Martín-Alcón, S.; Coll, L. Unraveling the Relative Importance of Factors Driving Post-Fire Regeneration Trajectories in Non-Serotinous Pinus Nigra Forests. For. Ecol. Manag. 2016, 361, 13–22. [Google Scholar] [CrossRef]
- Chu, T.; Guo, X. Remote Sensing Techniques in Monitoring Post-Fire Effects and Patterns of Forest Recovery in Boreal Forest Regions: A Review. Remote Sens. 2014, 6, 470–520. [Google Scholar] [CrossRef]
- Chergui, B.; Fahd, S.; Santos, X. Quercus Suber Forest and Pinus Plantations Show Different Post-Fire Resilience in Mediterranean North-Western Africa. Ann. For. Sci. 2018, 75, 64. [Google Scholar] [CrossRef]
- Falcucci, A.; Maiorano, L.; Boitani, L. Changes in Land-Use/Land-Cover Patterns in Italy and Their Implications for Biodiversity Conservation. Landsc. Ecol. 2006, 22, 617–631. [Google Scholar] [CrossRef]
- Nioti, F.; Xystrakis, F.; Koutsias, N.; Dimopoulos, P. A Remote Sensing and GIS Approach to Study the Long-Term Vegetation Recovery of a Fire-Affected Pine Forest in Southern Greece. Remote Sens. 2015, 7, 7712–7731. [Google Scholar] [CrossRef]
- Pérez-Cabello, F.; Montorio, R.; Alves, D.B. Remote Sensing Techniques to Assess Post-Fire Vegetation Recovery. Curr. Opin. Environ. Sci. Health 2021, 21, 100251. [Google Scholar] [CrossRef]
- Lazzeri, G.; Frodella, W.; Rossi, G.; Moretti, S. Multitemporal Mapping of Post-Fire Land Cover Using Multiplatform Prisma Hyperspectral and Sentinel-Uav Multispectral Data: Insights from Case Studies in Portugal and Italy. Sensors 2021, 21, 3982. [Google Scholar] [CrossRef]
- Pádua, L.; Guimarães, N.; Adão, T.; Sousa, A.; Peres, E.; Sousa, J. Effectiveness of Sentinel-2 in Multi-Temporal Post-Fire Monitoring When Compared with UAV Imagery. ISPRS Int. J. Geo-Inf. 2020, 9, 225. [Google Scholar] [CrossRef]
- Key, C.H.; Benson, N.C. Landscape Assessment (LA). In FIREMON: Fire Effects Monitoring and Inventory System; General Technical Report RMRS-GTR-164-CD; Lutes, D.C., Keane, R.E., Caratti, J.F., Key, C.H., Benson, N.C., Sutherland, S., Gangi, L.J., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006; Volume 164, p. LA-1-55. [Google Scholar]
- Hall-Beyer, M. GLCM Texture: A Tutorial v. 3.0 March 2017; University of Calgary: Calgary, AB, Canada, 2017. [Google Scholar] [CrossRef]
- Haralick, R.M.; Shanmugam, K.; Dinstein, I. Textural Features for Image Classification. IEEE Trans. Syst. Man Cybern. 1973, SMC-3, 610–621. [Google Scholar] [CrossRef]
- Isbaex, C.; Sousa, A.M.O.; Gonçalves, A.C. Forest Species Mapping Using Sentinel 2A Images for the Cen-Tral Alentejo Region (Portugal). In Proceedings of the XI Congresso Ibérico de AgroIngenieria, Online, 11–12 November 2021. [Google Scholar]
- Pausas, J.G.; Paula, S. Grasses and Fire: The Importance of Hiding Buds: A Response to Moore et al. (2019) ‘Effects of Drought and Fire on Resprouting Capacity of 52 Temperate Australian Perennial Native Grasses’. New Phytol. 2020, 226, 957–959. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. The Impact of Rural Fires on the Development of Invasive Species: Analysis of a Case Study with Acacia Dealbata Link. in Casal Do Rei (Seia, Portugal). Environments 2021, 8, 44. [Google Scholar] [CrossRef]
- Molina, J.R.; Lora, A.; Prades, C.; Rodríguez Y Silva, F. Roadside Vegetation Planning and Conservation: New Approach to Prevent and Mitigate Wildfires Based on Fire Ignition Potential. For. Ecol. Manag. 2019, 444, 163–173. [Google Scholar] [CrossRef]
- Lazzaro, L.; Giuliani, C.; Fabiani, A.; Agnelli, A.E.; Pastorelli, R.; Lagomarsino, A.; Benesperi, R.; Calamassi, R.; Foggi, B. Soil and Plant Changing after Invasion: The Case of Acacia Dealbata in a Mediterranean Ecosystem. Sci. Total Environ. 2014, 497–498, 491–498. [Google Scholar] [CrossRef]
- Lorenzo, P.; Pazos-Malvido, E.; Rubido-Bará, M.; Reigosa, M.J.; González, L. Invasion by the Leguminous Tree Acacia dealbata (Mimosaceae) Reduces the Native Understorey Plant Species in Different Communities. Aust. J. Bot. 2012, 60, 669. [Google Scholar] [CrossRef]
- Foody, G.M. Status of Land Cover Classification Accuracy Assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- Lu, D.; Weng, Q. A Survey of Image Classification Methods and Techniques for Improving Classification Performance. Int. J. Remote Sens. 2007, 28, 823–870. [Google Scholar] [CrossRef]
- Harvey, B.J.; Donato, D.C.; Turner, M.G. High and Dry: Post-fire Tree Seedling Establishment in Subalpine Forests Decreases with Post-fire Drought and Large Stand-replacing Burn Patches. Glob. Ecol. Biogeogr. 2016, 25, 655–669. [Google Scholar] [CrossRef]
- Prodromou, M.; Gitas, I.; Mettas, C.; Tzouvaras, M.; Themistocleous, K.; Konstantinidis, A.; Pamboris, A.; Hadjimitsis, D. Remote-Sensing-Based Prioritization of Post-Fire Restoration Actions in Mediterranean Ecosystems: A Case Study in Cyprus. Remote Sens. 2025, 17, 1269. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).