
Impact of Hyperferritinemia on Immune Modulation in Septic Diabetic Patients


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. Primary Culture
2.3. CD4+ T Cell Isolation
2.4. qRT-PCR
2.5. qRT-PCR Primer Sequences
2.6. Flow Cytometry
2.7. Proliferation Analysis
2.8. Measurement of Oxygen Consumption Rate (OCR)
2.9. Statistical Analysis
3. Results
3.1. High Ferritin Levels in Diabetic Patients Are Associated with Lymphopenia and Increased Mortality Rates
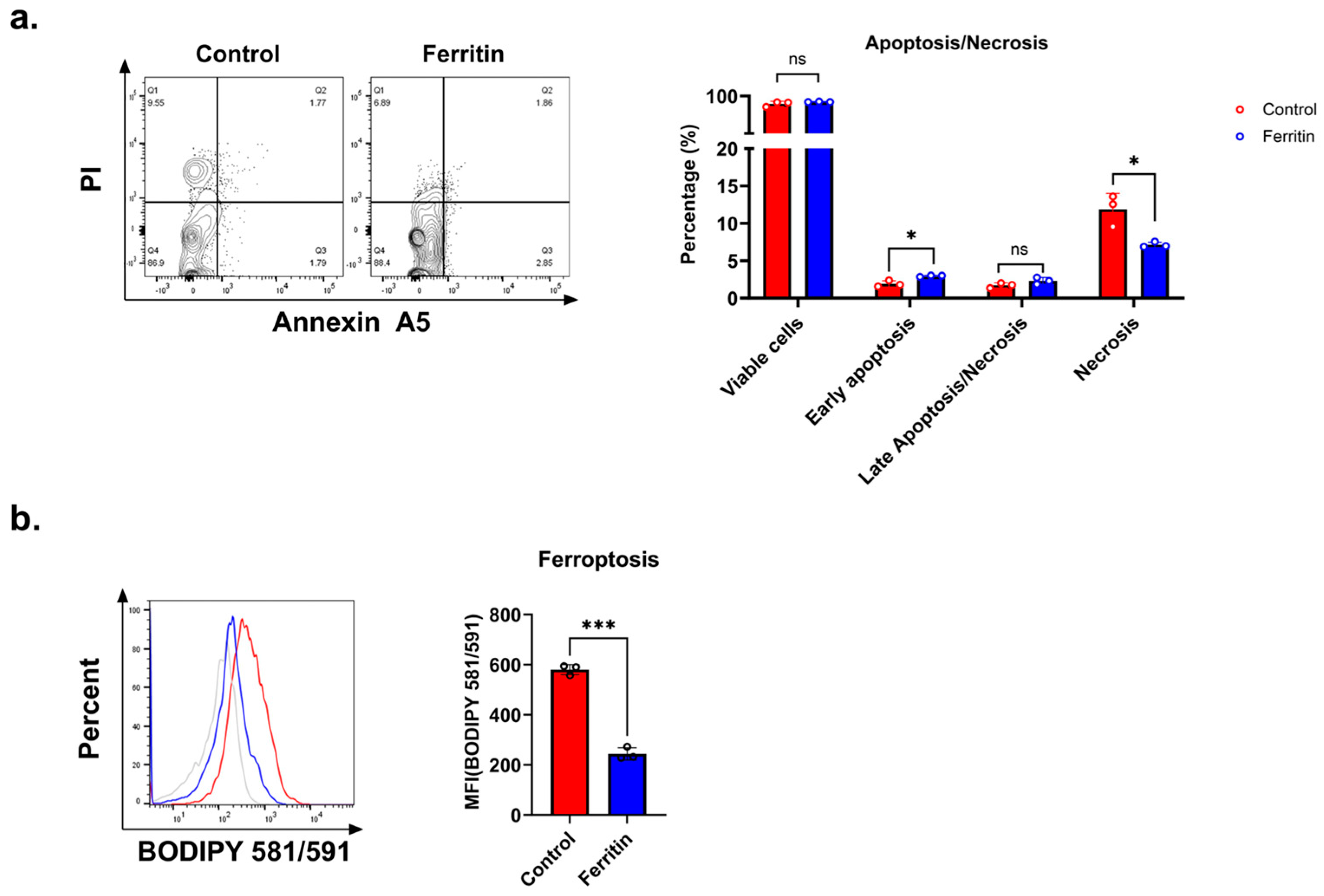
3.2. Elevated Ferritin Levels Do Not Directly Cause Increased CD4+ T Cell Death
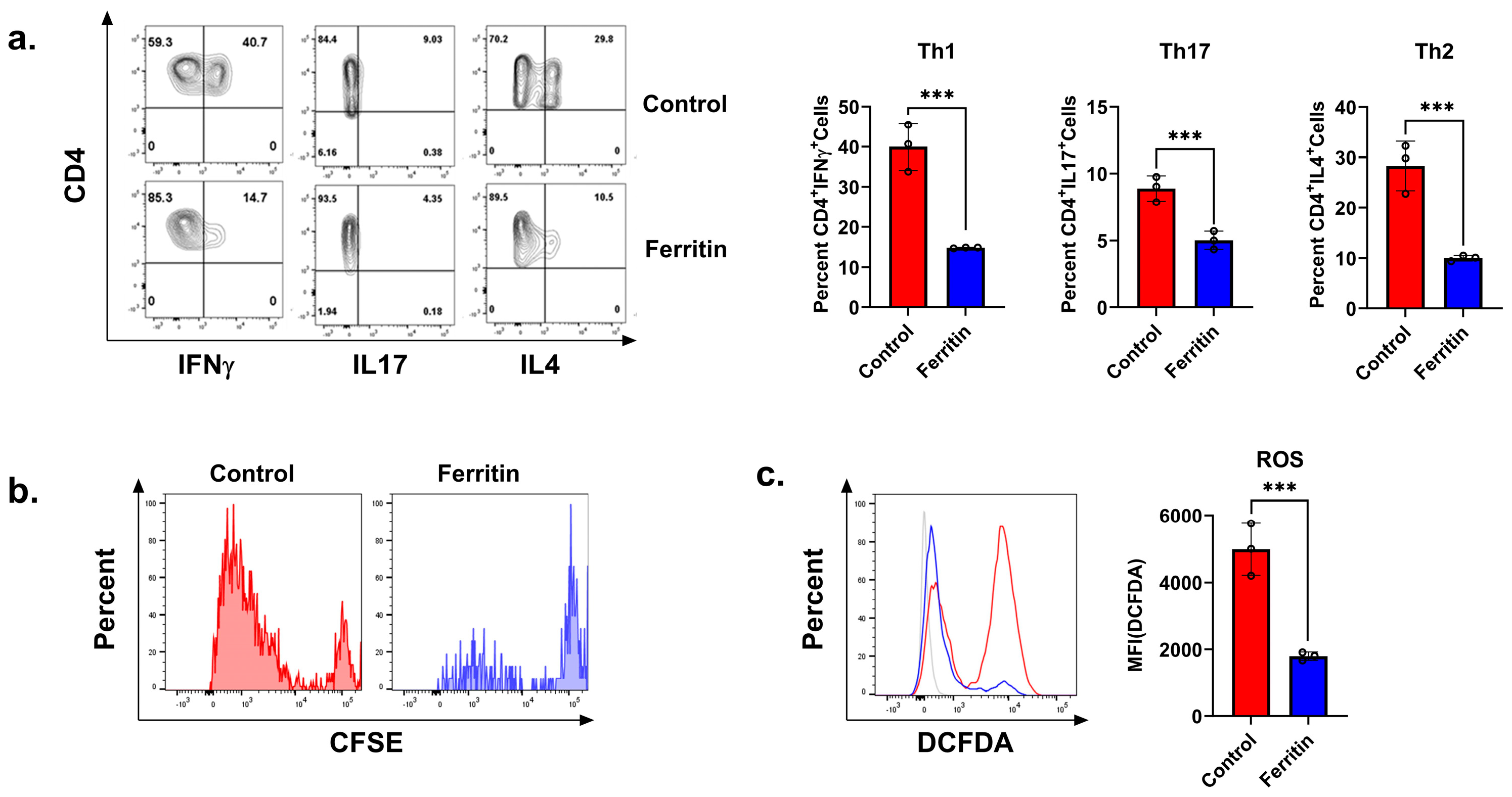
3.3. Ferritin Inhibits CD4+ T Cell Activation and Proliferation
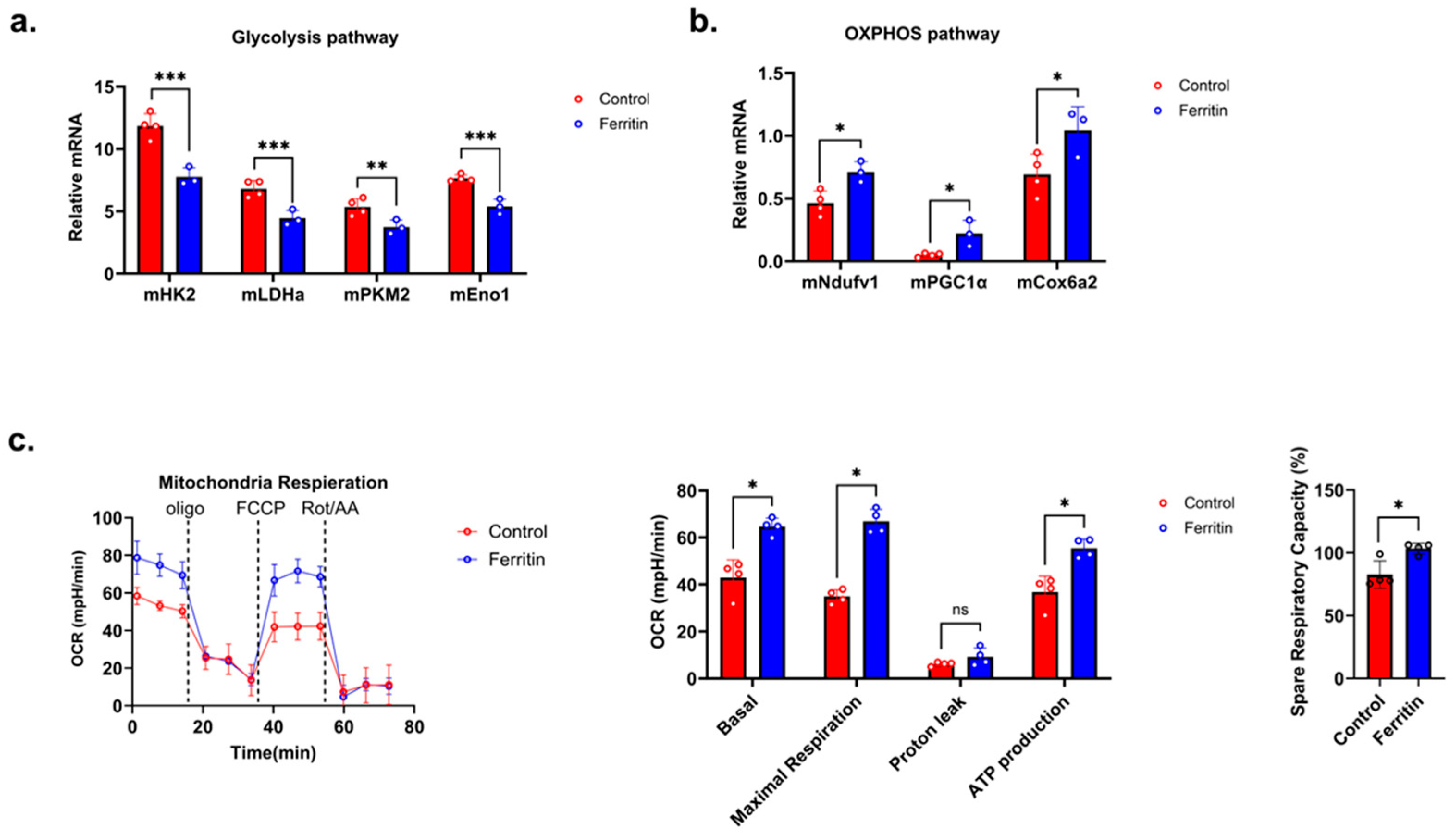
3.4. Ferritin Modulates CD4+ T Cell Proliferation by Enhancing OXPHOS and Suppressing Glycolytic Shift
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef]
- Turina, M.; Fry, D.E.; Polk, H.C., Jr. Acute hyperglycemia and the innate immune system: Clinical, cellular, and molecular aspects. Crit. Care Med. 2005, 33, 1624–1633. [Google Scholar] [CrossRef]
- Xiu, F.; Stanojcic, M.; Diao, L.; Jeschke, M.G. Stress hyperglycemia, insulin treatment, and innate immune cells. Int. J. Endocrinol. 2014, 2014, 486403. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan-Bogdan, M.; McDonnell, M.E.; Shin, H.; Rehman, Q.; Hasturk, H.; Apovian, C.M.; Nikolajczyk, B.S. Elevated proinflammatory cytokine production by a skewed T cell compartment requires monocytes and promotes inflammation in type 2 diabetes. J. Immunol. 2011, 186, 1162–1172. [Google Scholar] [CrossRef]
- Kernan, K.F.; Carcillo, J.A. Hyperferritinemia and inflammation. Int. Immunol. 2017, 29, 401–409. [Google Scholar] [CrossRef]
- Zarjou, A.; Black, L.M.; McCullough, K.R.; Hull, T.D.; Esman, S.K.; Boddu, R.; Varambally, S.; Chandrashekar, D.S.; Feng, W.; Arosio, P.; et al. Ferritin Light Chain Confers Protection Against Sepsis-Induced Inflammation and Organ Injury. Front. Immunol. 2019, 10, 131. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Wang, P.; Cui, Y.; Liu, Y.; Li, Z.; Bai, H.; Zhao, Y.; Chang, Y.Z. Mitochondrial ferritin alleviates apoptosis by enhancing mitochondrial bioenergetics and stimulating glucose metabolism in cerebral ischemia reperfusion. Redox Biol. 2022, 57, 102475. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.P.; Chinta, K.C.; Saini, V.; Glasgow, J.N.; Hull, T.D.; Traylor, A.; Rey-Stolle, F.; Soares, M.P.; Madansein, R.; Rahman, M.A.; et al. Ferritin H Deficiency in Myeloid Compartments Dysregulates Host Energy Metabolism and Increases Susceptibility to Mycobacterium tuberculosis Infection. Front. Immunol. 2018, 9, 860. [Google Scholar] [CrossRef]
- Paillon, N.; Ung, T.P.L.; Dogniaux, S.; Stringari, C.; Hivroz, C. Label-free single-cell live imaging reveals fast metabolic switch in T lymphocytes. Mol. Biol. Cell 2024, 35, ar11. [Google Scholar] [CrossRef]
- Nastasi, C.; Willerlev-Olsen, A.; Dalhoff, K.; Ford, S.L.; Gadsboll, A.O.; Buus, T.B.; Gluud, M.; Danielsen, M.; Litman, T.; Bonefeld, C.M.; et al. Inhibition of succinate dehydrogenase activity impairs human T cell activation and function. Sci. Rep. 2021, 11, 1458. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Pretorius, E. Serum ferritin is an important inflammatory disease marker, as it is mainly a leakage product from damaged cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef]
- Babitt, J.L.; Lin, H.Y. Molecular Mechanisms of Hepcidin Regulation: Implications for the Anemia of CKD. Am. J. Kidney Dis. 2010, 55, 726–741. [Google Scholar] [CrossRef]
- Zhang, D.-L.; Rouault, T.A. How does hepcidin hinder ferroportin activity? Blood 2018, 131, 840–842. [Google Scholar] [CrossRef]
- Afsar, R.E.; Kanbay, M.; Ibis, A.; Afsar, B. In-depth review: Is hepcidin a marker for the heart and the kidney? Mol. Cell. Biochem. 2021, 476, 3365–3381. [Google Scholar] [CrossRef]
- Haschka, D.; Hoffmann, A.; Weiss, G. Iron in immune cell function and host defense. Semin. Cell Dev. Biol. 2021, 115, 27–36. [Google Scholar] [CrossRef]
- Girelli, D.; Marchi, G.; Busti, F.; Vianello, A. Iron metabolism in infections: Focus on COVID-19. Semin. Hematol. 2021, 58, 182–187. [Google Scholar] [CrossRef]
- Li, Y.; Du, Y.; Zhou, Y.; Chen, Q.; Luo, Z.; Ren, Y.; Chen, X.; Chen, G. Iron and copper: Critical executioners of ferroptosis, cuproptosis and other forms of cell death. Cell Commun. Signal. 2023, 21, 327. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.C.; Teles, M.J.; Silva, T.; Bento, C.M.; Alves, I.S.; Pereira, L.; Guimarães, J.T.; Porto, G.; Oliveira, P.; Gomes, M.S. Iron Related Biomarkers Predict Disease Severity in a Cohort of Portuguese Adult Patients during COVID-19 Acute Infection. Viruses 2021, 13, 2482. [Google Scholar] [CrossRef] [PubMed]
- Zitt, E.; Sturm, G.; Kronenberg, F.; Neyer, U.; Knoll, F.; Lhotta, K.; Weiss, G. Iron Supplementation and Mortality in Incident Dialysis Patients: An Observational Study. PLoS ONE 2014, 9, e114144. [Google Scholar] [CrossRef]
- Zager, R.A.; Johnson, A.C.; Hanson, S.Y. Parenteral iron therapy exacerbates experimental sepsis Rapid Communication. Kidney Int. 2004, 65, 2108–2112. [Google Scholar] [CrossRef]
- Cheng, Y.; Yue, L.; Wang, Z.; Zhang, J.; Xiang, G. Hyperglycemia associated with lymphopenia and disease severity of COVID-19 in type 2 diabetes mellitus. J. Diabetes Complicat. 2021, 35, 107809. [Google Scholar] [CrossRef]
- Ash, S.; Yarkoni, S.; Askenasy, N. Lymphopenia is detrimental to therapeutic approaches to type 1 diabetes using regulatory T cells. Immunol. Res. 2014, 58, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Muller, Y.D.; Golshayan, D.; Ehirchiou, D.; Wyss, J.C.; Giovannoni, L.; Meier, R.; Serre-Beinier, V.; Puga Yung, G.; Morel, P.; Buhler, L.H.; et al. Immunosuppressive effects of streptozotocin-induced diabetes result in absolute lymphopenia and a relative increase of T regulatory cells. Diabetes 2011, 60, 2331–2340. [Google Scholar] [CrossRef]
- Monneret, G.; Venet, F. A rapidly progressing lymphocyte exhaustion after severe sepsis. Crit. Care 2012, 16, 140. [Google Scholar] [CrossRef]
- Boomer, J.S.; Shuherk-Shaffer, J.; Hotchkiss, R.S.; Green, J.M. A prospective analysis of lymphocyte phenotype and function over the course of acute sepsis. Crit. Care 2012, 16, R112. [Google Scholar] [CrossRef] [PubMed]
- Brady, J.; Horie, S.; Laffey, J.G. Role of the adaptive immune response in sepsis. Intensive Care Med. Exp. 2020, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Yuan, S.; Zhang, J. The potential immunological mechanisms of sepsis. Front. Immunol. 2024, 15, 1434688. [Google Scholar] [CrossRef]
- Kaye, T.B.; Guay, A.T. Increased serum ferritin levels in patients with diabetes mellitus. Mayo Clin. Proc. 1994, 69, 498–499. [Google Scholar] [CrossRef]
- Finfer, S.; Venkatesh, B.; Hotchkiss, R.S.; Sasson, S.C. Lymphopenia in sepsis-an acquired immunodeficiency? Immunol. Cell Biol. 2023, 101, 535–544. [Google Scholar] [CrossRef]
- Girardot, T.; Rimmele, T.; Venet, F.; Monneret, G. Apoptosis-induced lymphopenia in sepsis and other severe injuries. Apoptosis 2017, 22, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Gerriets, V.A.; Kishton, R.J.; Nichols, A.G.; Macintyre, A.N.; Inoue, M.; Ilkayeva, O.; Winter, P.S.; Liu, X.; Priyadharshini, B.; Slawinska, M.E.; et al. Metabolic programming and PDHK1 control CD4+ T cell subsets and inflammation. J. Clin. Investig. 2015, 125, 194–207. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DM (n = 69) | Non-DM (n = 192) | p | |
|---|---|---|---|
| Age (years) | 56.4 ± 12.3 | 57.9 ± 10.2 | 0.07 |
| Sex, male, n (%) | 40 (58.00%) | 96 (53.49%) | 0.32 |
| Death within 28 days, n (%) | 15 (21.7%) | 22 (11.5%) | 0.05 |
| Lymphocyte counts (103/µL) | 1.2 ± 1.6 | 2.1 ± 3.6 | 0.007 |
| Platelet counts (103/µL) | 180.5 ± 141.5 | 244.8 ± 135.2 | 0.001 |
| Ferritin (ng/mL) | 1945.1 ± 329.6 | 907.3 ± 250.8 | 0.021 |
| Iron (µg/dL) | 41.0 ± 26.5 | 56.6 ± 38.7 | 0.189 |
| CRP (mg/L) | 14.8 ± 12.2 | 8.7 ± 10.0 | <0.001 |
| Procalcitonin (ng/mL) | 23.9 ± 31.8 | 17.8 ± 29.9 | 0.203 |
| AST (U/L) | 153.5 ± 467.1 | 167.8 ± 947.4 | 0.874 |
| ALT (U/L) | 78.4 ± 162.0 | 65.5 ± 233.3 | 0.624 |
| Cr (mg/dL) | 1.9 ± 1.9 | 1.4 ± 1.5 | 0.058 |
| eGFR (mL/min/1.73 m2) | 42.1 ± 31.9 | 55.4 ± 42.0 | 0.023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-J.; Choi, E.-J.; Lee, I.-K. Impact of Hyperferritinemia on Immune Modulation in Septic Diabetic Patients. Diabetology 2024, 5, 344-355. https://doi.org/10.3390/diabetology5030026
Kim M-J, Choi E-J, Lee I-K. Impact of Hyperferritinemia on Immune Modulation in Septic Diabetic Patients. Diabetology. 2024; 5(3):344-355. https://doi.org/10.3390/diabetology5030026
Chicago/Turabian StyleKim, Min-Ji, Eun-Jung Choi, and In-Kyu Lee. 2024. "Impact of Hyperferritinemia on Immune Modulation in Septic Diabetic Patients" Diabetology 5, no. 3: 344-355. https://doi.org/10.3390/diabetology5030026