
Germination of Pyrodinium bahamense Cysts from a Pristine Lagoon in San José Island, Gulf of California: Implications of Long-Term Survival

 , ,
, ,  ,
,  and
and 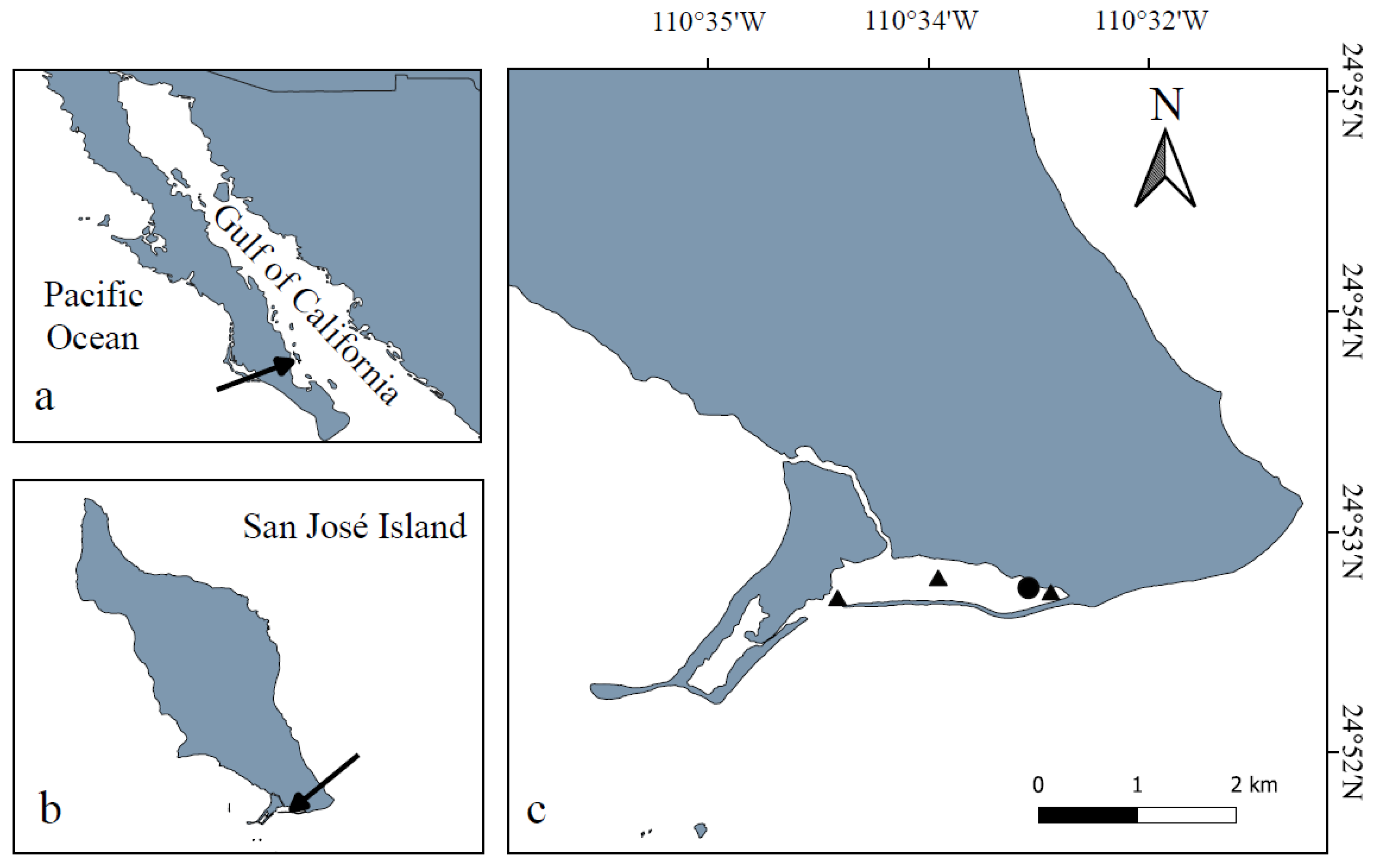{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
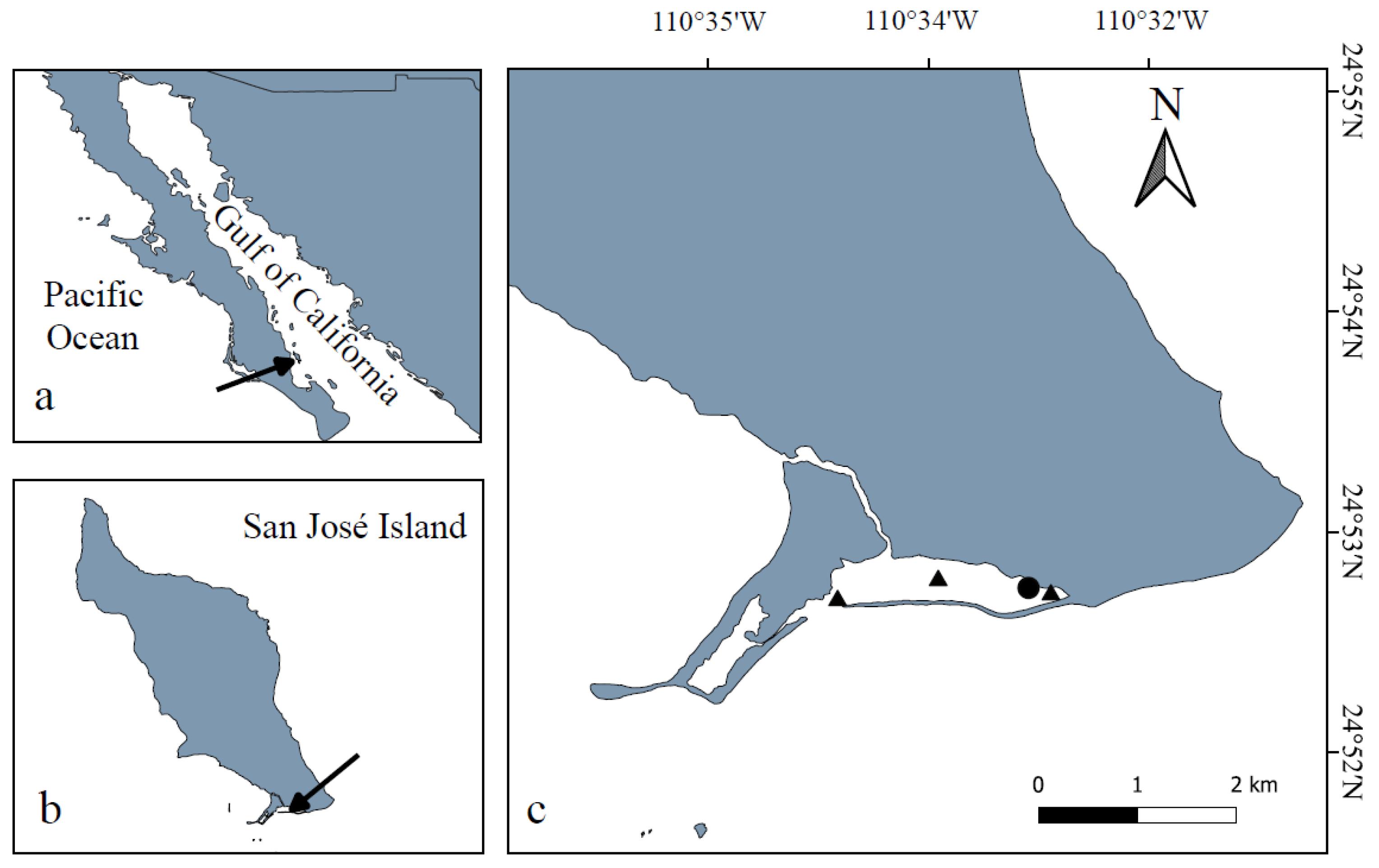
2.1. Study Area
2.2. Sampling
2.3. Laboratory Analysis
2.3.1. Core Dating
2.3.2. Germination Experiments with Cysts from Surface Sediments: Effects of Light and Nutrients Conditions
2.3.3. Abundance and Viability of Dinoflagellate Cysts from the Sediment Core
2.4. Statistical Analysis
3. Results
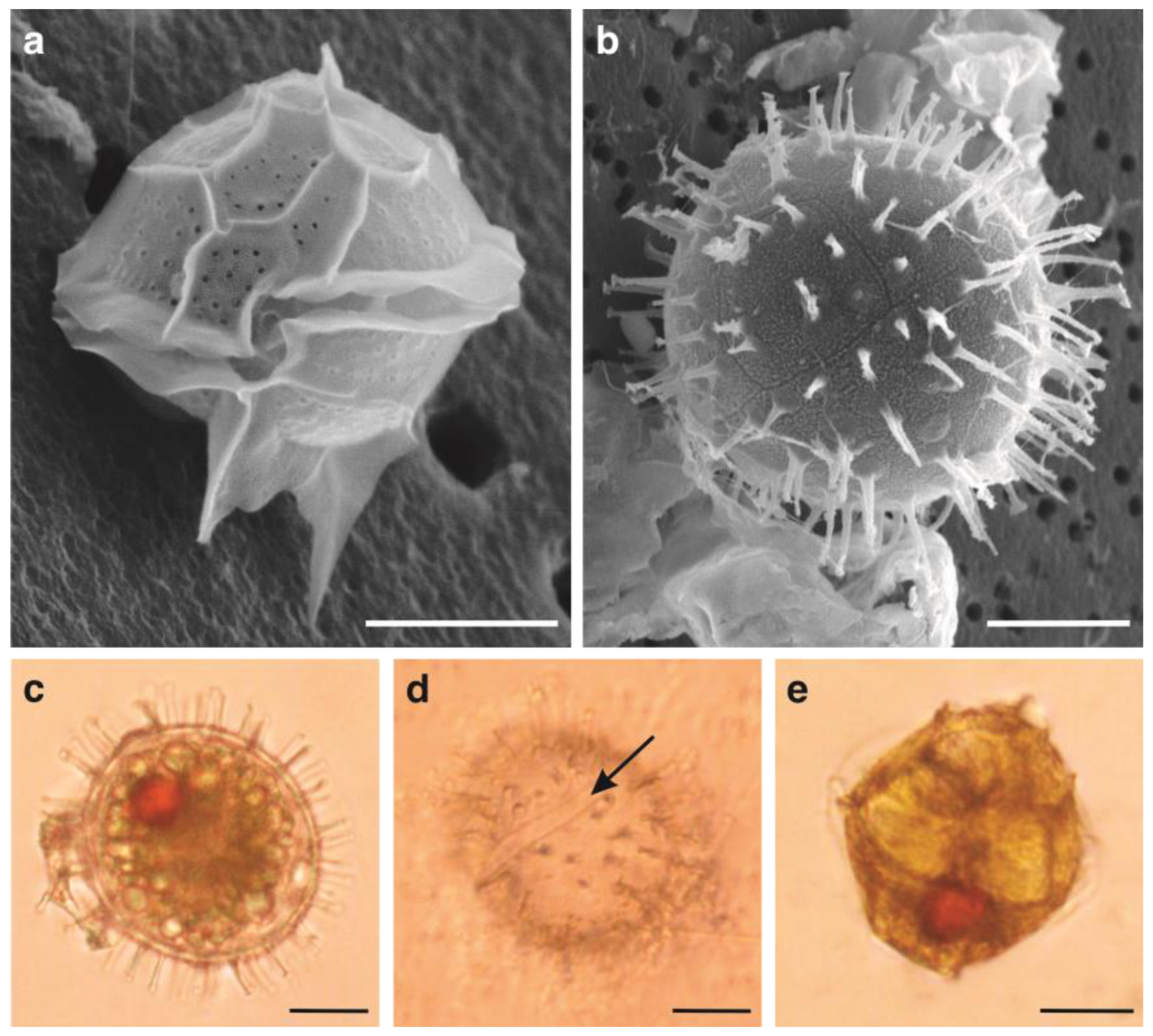
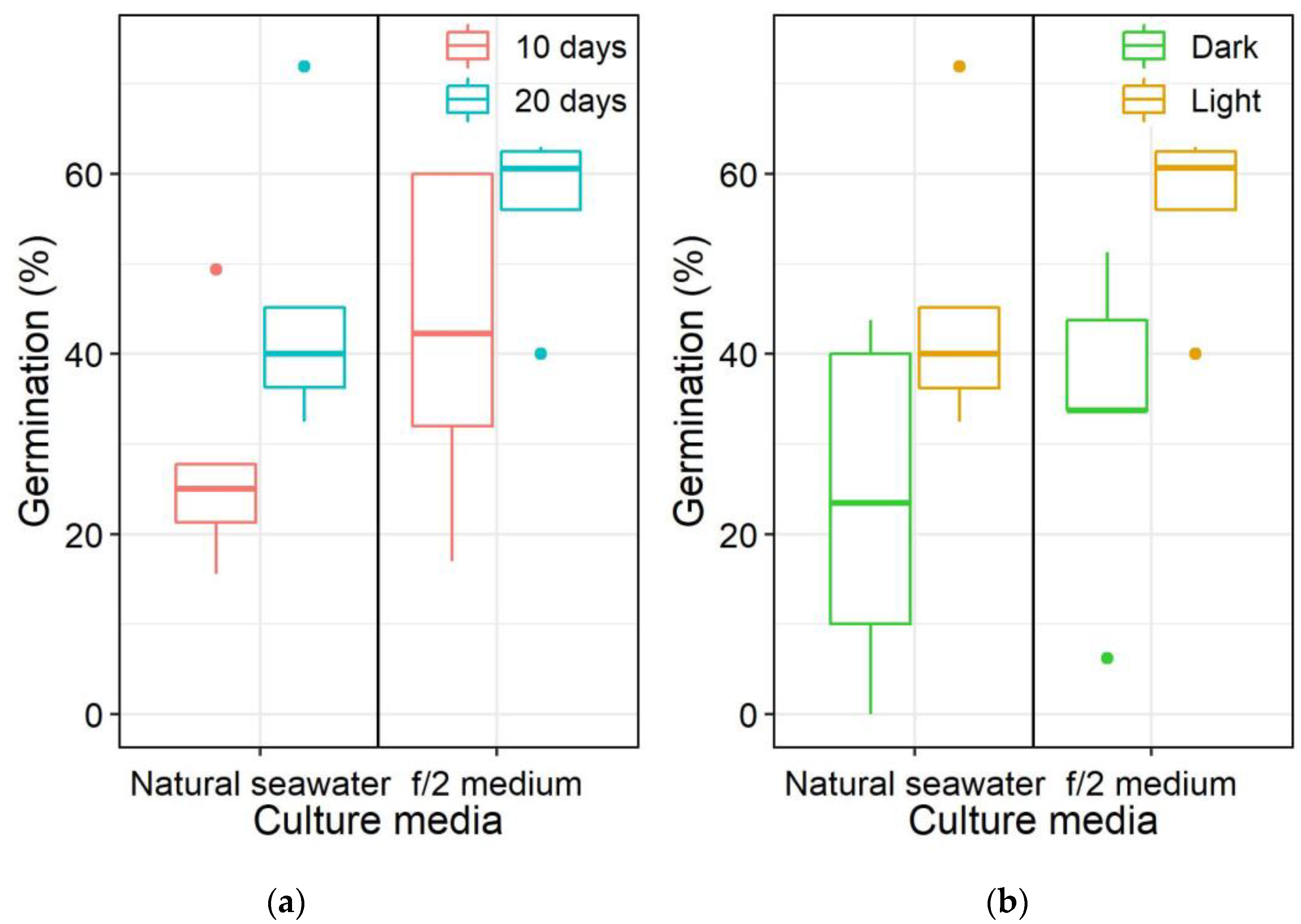
3.1. Germination of Pyrodinium bahamense Cysts from Surface Sediments
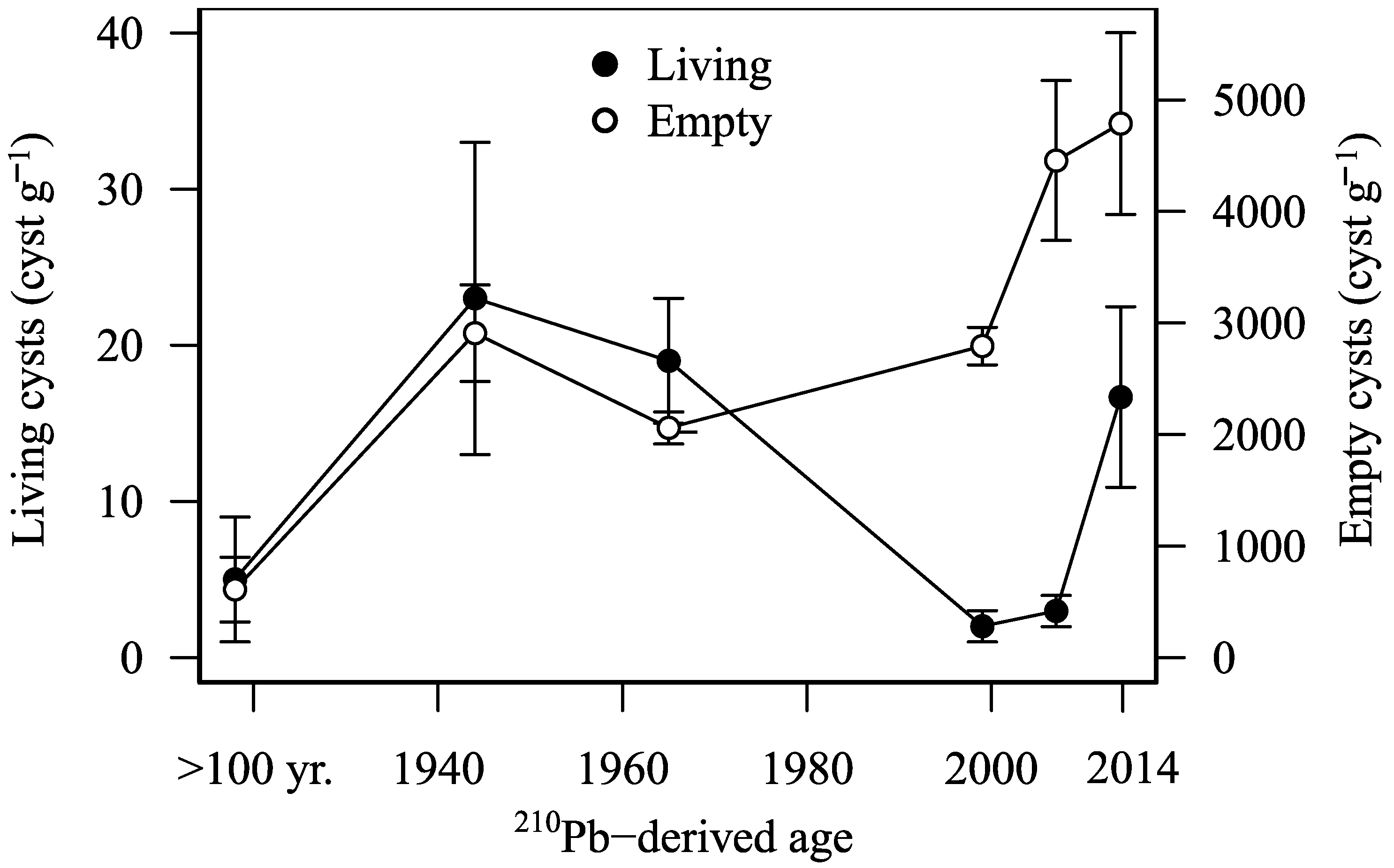
3.2. Abundance of Living Pyrodinium bahamense Cysts in the Sediment Core
3.3. Long-term Viability of Dinoflagellate Cysts from San José Core Sections
4. Discussion
4.1. Effects of Light and Nutrients on Pyrodinium bahamense Cyst Germination
4.2. Long-Term Viability of Living Pyrodinium bahamense Cysts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saldarriaga, J.F.; Taylor, J.R. Dinoflagellata. In Handbook of the Protists; Archibald, J.M., Alastair, G.B., Simpson, C.H.S., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–54. [Google Scholar] [CrossRef]
- Lundholm, N.; Churro, C.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Larsen, J.; Mertens, K.; Moestrup, Ø.; Zingone, A. (Eds.) IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. 2009. Available online: https://www.marinespecies.org/hab (accessed on 28 October 2022).
- Head, M. Modern dinoflagellate cysts and their biological affinities. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Dallas, TX, USA, 1996; pp. 1197–1248. [Google Scholar]
- Matsuoka, K.; Head, M.J.; Lewis, J.M.; Marret, F.; Bradley, L. Clarifying cyst–motile stage relationships in dinoflagellates. In Biological and Geological Perspectives of Dinoflagellates; Lewis, J.M., Marret, F., Bradley, L., Eds.; Geological Society: London, UK, 2013; pp. 325–350. [Google Scholar]
- Likumahua, S.; Sangiorgi, F.; de Boer, M.K.; Tatipatta, W.M.; Pelasula, D.D.; Polnaya, D.; Hehuwat, J.; Siahaya, D.M.; Buma, A.G.J. Dinoflagellate cyst distribution in surface sediments of Ambon Bay (eastern Indonesia): Environmental conditions and harmful blooms. Mar. Pollut. Bull. 2021, 166, 112269. [Google Scholar] [CrossRef] [PubMed]
- Kremp, A. Diversity of dinoflagellate life cycles. In Biological and Geological Perspectives of Dinoflagellates; Lewis, J.M., Marret, F., Bradley, L., Eds.; Geological Society: London, UK, 2013; pp. 197–205. [Google Scholar]
- Butman, B.; Aretxabaleta, A.L.; Dickhudt, P.J.; Dalyander, P.S.; Sherwood, C.R.; Anderson, D.M.; Keafer, B.A.; Signell, R.P. Investigating the importance of sediment resuspension in Alexandrium fundyense cyst population dynamics in the Gulf of Maine. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 103, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Wall, D. Taxonomy and cysts of red-tide dinoflagellates. In Proceedings of the 1st International Conference on Toxic Dinoflagellate Blooms, Boston, MA, USA, 4 November 1974; Massachusetts Science and Technology Foundation: Cambridge, MA, USA, 1975; pp. 249–255. [Google Scholar]
- Matsuoka, K.; Fukuyo, Y. Technical Guide for Modern Dinoflagellate Cyst Study; Japan Society for the Promotion of Science: Tokyo, Japan, 2000; pp. 6–9. [Google Scholar]
- Montresor, M.; Nuzzo, L.; Mazzocchi, M.G. Viability of dinoflagellate cysts after the passage through the copepod gut. J. Exp. Mar. Bio. Ecol. 2003, 287, 209–221. [Google Scholar] [CrossRef]
- Laabir, M.; Amzil, Z.; Lassus, P.; Masseret, E.; Tapilatu, Y.; De Vargas, R.; Grzebyk, D. Viability, growth and toxicity of Alexandrium catenella and Alexandrium minutum (Dinophyceae) following ingestion and gut passage in the oyster Crassostrea gigas. Aquat. Living Resour. 2007, 20, 51–57. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Gu, H.; Wang, Z.; Liu, D.; Wang, Y.; Lu, D.; Hu, Z.; Deng, Y.; Shang, L.; Qi, Y. Exploration of resting cysts (stages) and their relevance for possibly HABs-causing species in China. Harmful Algae 2021, 107, 102050. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Stock, C.A.; Keafer, B.A.; Nelson, A.B.; Thompson, B.; McGillicuddy Jr, D.J.; Keller, M.; Matrai, P.A.; Martin, J. Alexandrium fundyense cyst dynamics in the Gulf of Maine. Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2522–2542. [Google Scholar] [CrossRef]
- Anderson, D.M.; Keafer, B.A. An endogenous annual clock in the toxic marine dinoflagellate Gonyaulax tamarensis. Nature 1987, 325, 616–617. [Google Scholar] [CrossRef]
- Matrai, P.; Thompson, B.; Keller, M. Circannual excystment of resting cysts of Alexandrium spp. from eastern Gulf of Maine populations. Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2560–2568. [Google Scholar] [CrossRef]
- Fischer, A.D.; Brosnahan, M.L.; Anderson, D.M. Quantitative response of Alexandrium catenella cyst dormancy to cold exposure. Protist 2018, 169, 645–661. [Google Scholar] [CrossRef]
- Lopez, C.B.; Karim, A.; Murasko, S.; Marot, M.; Smith, C.G.; Corcoran, A.A. Temperature mediates secondary dormancy in resting cysts of Pyrodinium bahamense (Dinophyceae). J. Phycol. 2019, 55, 924–935. [Google Scholar] [CrossRef]
- Brosnahan, M.L.; Fischer, A.D.; Lopez, C.B.; Moore, S.K.; Anderson, D.M. Cyst-forming dinoflagellates in a warming climate. Harmful Algae 2020, 91, 101728. [Google Scholar] [CrossRef] [PubMed]
- Genovesi-Giunti, B.; Laabir, M.; Vaquer, A. The benthic resting cysts: A key factor in harmful dinoflagellate blooms—A review. Vie Milieu 2006, 56, 327–337. [Google Scholar]
- Anderson, D.M.; Taylor, C.D.; Armbrust, E.V. The effects of darkness and anaerobiosis on dinoflagellate cyst germination. Limnol. Oceanogr. 1987, 32, 340–351. [Google Scholar] [CrossRef]
- Blanco, E.P.; Lewis, J.; Aldridge, J. The germination characteristics of Alexandrium minutum (Dinophyceae), a toxic dinoflagellate from the Fal estuary (UK). Harmful Algae 2009, 8, 518–522. [Google Scholar] [CrossRef]
- Kremp, A.; Anderson, D.M. Factors regulating germination of resting cysts of the spring bloom dinoflagellate Scrippsiella hangoei from the northern Baltic Sea. J. Plankton Res. 2000, 22, 1311–1327. [Google Scholar] [CrossRef]
- Kim, B.; Park, M.; Hwang, S.; Han, M. Excystment patterns of the freshwater dinoflagellate Peridinium bipes (Dinophyceae) in Juam Reservoir, Korea. Aquat. Microb. Ecol. 2007, 47, 213–221. [Google Scholar] [CrossRef]
- Nuzzo, L.; Montresor, M. Different excystment patterns in two calcareous cyst-producing species of the dinoflagellate genus Scrippsiella. J. Plankton Res. 1999, 21, 2009–2018. [Google Scholar] [CrossRef]
- Cannon, J.A. Germination of the toxic dinoflagellate, Alexandrium minutum, from sediments in the Port River, South Australia. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: New York, NY, USA, 1993; pp. 103–107. [Google Scholar]
- Bravo, I.; Anderson, D.M. The effects of temperature, growth medium and darkness on excystment and growth of the toxic dinoflagellate Gymnodinium catenatum from northwest Spain. J. Plankton Res. 1994, 16, 513–525. [Google Scholar] [CrossRef]
- Genovesi, B.; Laabir, M.; Masseret, E.; Collos, Y.; Vaquer, A.; Grzebyk, D. Dormancy and germination features in resting cysts of Alexandrium tamarense species complex (Dinophyceae) can facilitate bloom formation in a shallow lagoon (Thau, southern France). J. Plankton Res. 2009, 31, 1209–1224. [Google Scholar] [CrossRef]
- Vahtera, E.; Crespo, B.G.; McGillicuddy, D.J.; Olli, K.; Anderson, D.M. Alexandrium fundyense cyst viability and germling survival in light vs. dark at a constant low temperature. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 103, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.C.; Roy, S.; Levasseur, M.; Anderson, D.M. Control of germination of Alexandrium tamarense (Dinophyceae) cysts from the lower St. Lawrence estuary (Canada). J. Phycol. 1998, 34, 242–249. [Google Scholar] [CrossRef]
- Band-Schmidt, C.J.; Lechuga-Devéze, C.H.; Kulis, D.M.; Anderson, D.M. Culture studies of Alexandrium affine (Dinophyceae), a non-toxic cyst forming dinoflagellate from Bahía Concepción, Gulf of California. Bot. Mar. 2003, 46, 44–54. [Google Scholar] [CrossRef]
- Shikata, T.; Nagasoe, S.; Matsubara, T.; Yamasaki, Y.; Shimasaki, Y.; Oshima, Y.; Uchida, T.; Jenkinson, I.R.; Honjo, T. Encystment and excystment of Gyrodinium instriatum Freudenthal et Lee. J. Oceanogr. 2008, 64, 355–365. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I.; Garcés, E. Effects of nutritional factors and different parental crosses on the encystment and excystment of Alexandrium catenella (Dinophyceae) in culture. Phycologia 2005, 44, 658–670. [Google Scholar] [CrossRef]
- Binder, B.J.; Anderson, D.M. Physiological and environmental control of germination in Scrippsiella trochoidea (Dinophyceae) resting cysts. J. Phycol. 1987, 23, 99–107. [Google Scholar] [CrossRef]
- Hernández-Becerril, D.U.; Alonso-Rodríguez, R.; Álvarez-Góngora, C.; Barón-Campis, S.A.; Ceballos-Corona, G.; Herrera-Silveira, J.; Meave del Castillo, M.E.; Juárez-Ruíz, N.; Merino-Virgilio, F.; Morales-Blake, A.; et al. Toxic and harmful marine phytoplankton and microalgae (HABs) in Mexican coasts. J. Environ. Sci. Health Part A 2007, 42, 1349–1363. [Google Scholar] [CrossRef]
- García-Mendoza, E.; Quijano-Scheggia, S.; Olivos-Ortiz, A.; Núñez-Vázquez, E.; Pérez-Morales, A. Introducción General. In Florecimientos Algales Nocivos en México; García-Mendoza, E., Quijano-Scheggia, S.I., Olivos-Ortiz, A., Núñez-Vázquez, E.J., Eds.; CICESE: Ensenada, Mexico, 2016; pp. 10–19. [Google Scholar]
- Morquecho, L. Pyrodinium bahamense one the most significant harmful dinoflagellate in Mexico. Front. Mar. Sci. 2019, 6, 1. [Google Scholar] [CrossRef]
- Morquecho, L.; Alonso-Rodríguez, R.; Arreola-Lizárraga, J.A.; Reyes-Salinas, A. Factors associated with moderate blooms of Pyrodinium bahamense in shallow and restricted subtropical lagoons in the Gulf of California. Bot. Mar. 2012, 55, 611–623. [Google Scholar] [CrossRef]
- Martínez-Hernández, E.; Hernández-Campos, H.E. Distribución de quistes de dinoflagelados y acritarcas en sedimentos holocénicos del Golfo de California. Paleontol. Mex. 1991, 57, 1–133. [Google Scholar]
- Helenes, J.; Carreño, A.L.; Carrillo, R.M. Middle to late Miocene chronostratigraphy and development of the northern Gulf of California. Mar. Micropaleontol. 2009, 72, 10–25. [Google Scholar] [CrossRef]
- Castañeda-Quezada, R.; Helenes, J.; García-Mendoza, E.; Ramírez-Mendoza, R. Assemblages of dinoflagellate resistance cysts and copepod eggs in superficial sediments at the upper Gulf of California. Cont. Shelf Res. 2022, 235, 104648. [Google Scholar] [CrossRef]
- Morquecho, L.; Alonso-Rodríguez, R.; Martínez-Tecuapacho, G.A. Cyst morphology, germination characteristics, and potential toxicity of Pyrodinium bahamense in the Gulf of California. Bot. Mar. 2014, 57, 303–314. [Google Scholar] [CrossRef]
- Cuellar-Martinez, T.; Alonso-Rodríguez, R.; Ruiz-Fernández, A.C.; de Vernal, A.; Morquecho, L.; Limoges, A.; Henry, M.; Sanchez-Cabeza, J.A. Environmental forcing on the flux of organic-walled dinoflagellate cysts in recent sediments from a subtropical lagoon in the Gulf of California. Sci. Total Environ. 2018, 621, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Bernal, G.; Ripa, P.; Herguera, J.C. Oceanographic and climatic variability in the lower Gulf of California: Links with the tropics and north Pacific. Cienc. Mar. 2001, 27, 595–617. [Google Scholar] [CrossRef]
- Lluch-Cota, S.E.; Parés-Sierra, A.; Magaña-Rueda, V.O.; Arreguín-Sánchez, F.; Bazzino, G.; Herrera-Cervantes, H.; Lluch-Belda, D. Changing climate in the Gulf of California. Prog. Oceanogr. 2010, 87, 114–126. [Google Scholar] [CrossRef]
- Martínez-López, A.; Flores-Castillo, O.D.L.A.; Saldívar-Lucio, R.; Escobedo-Urías, D.C.; Verdugo-Díaz, G.; Pérez-Cruz, L.; Acevedo-Acosta, J.D. Paleoclimate of the Gulf of California (Northwestern Mexico) during the last 2000 years. In The Holocene and Anthropocene Environmental History of Mexico; Torrescano-Valle, N., Islebe, G.A., Roy, P.D., Eds.; Springer: Cham, Switzerland, 2019; pp. 7–38. [Google Scholar] [CrossRef]
- Goñi, M.A.; Hartz, M.D.; Thunell, R.C.; Tappa, E. Oceanographic considerations for the application of the alkenone-based paleotemperature U37K´ index in the Gulf of California. Geoch. et Cosmoch. Acta 2001, 65, 545–555. [Google Scholar] [CrossRef]
- Sanchez-Cabeza, J.A.; Herrera-Becerril, C.A.; Carballo, J.L.; Yáñez, B.; Alvarez-Sanchez, L.F.; Cardoso-Mohedano, J.G.; Ruiz-Fernández, A.C. Rapid surface water warming and impact of the recent (2013–2016) temperature anomaly in shallow coastal waters at the eastern entrance of the Gulf of California. Prog. Oceanogr. 2022, 202, 102746. [Google Scholar] [CrossRef]
- Cuervo-Robayo, A.P.; Ureta, C.; Gómez-Albores, M.A.; Meneses-Mosquera, A.K.; Téllez-Valdés, O.; Martínez-Meyer, E. One hundred years of climate change in Mexico. PLoS ONE 2020, 15, e0209808. [Google Scholar] [CrossRef]
- Martínez-Austria, P.F.; Jano-Pérez, J.A. Climate change and extreme temperature trends in the Baja California Peninsula, Mexico. Air Soil Water Res. 2021, 14, 117862212110107. [Google Scholar] [CrossRef]
- Usup, G.; Kulis, D.M.; Anderson, D.M. Growth and toxin production of the toxic dinoflagellate Pyrodinium bahamense var. compressum in laboratory cultures. Nat. Toxins 1994, 2, 254–262. [Google Scholar] [CrossRef]
- Lundholm, N.; Ribeiro, S.; Andersen, T.J.; Koch, T.; Godhe, A.; Ekelund, F.; Ellegaard, M. Buried alive—Germination of up to a century-old marine protist resting stages. Phycologia 2011, 50, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.; Berge, T.; Lundholm, N.; Andersen, T.J.; Abrantes, F.; Ellegaard, M. Phytoplankton growth after a century of dormancy illuminates past resilience to catastrophic darkness. Nat. Commun. 2011, 2, 311. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, A.; Nagai, S.; Kudo, I.; Tanizawa, K. Viability of Alexandrium tamarense cysts in the sediment of Funka Bay, Hokkaido, Japan: Over a hundred year survival times for cysts. Harmful Algae 2012, 16, 81–88. [Google Scholar] [CrossRef]
- Ellegaard, M.; Ribeiro, S.; Lundholm, N.; Andersen, T.J.; Berge, T.; Ekelund, F.; Godhe, A. Using the sediment archive of living dinoflagellate cysts and other protist resting stages to study temporal population dynamics. In Biological and Geological Perspectives of Dinoflagellates; Lewis, J.M., Marret, F., Bradley, L.R., Eds.; Geological Society of London: London, UK, 2013; pp. 149–153. [Google Scholar]
- SMN Normales Climatológicas por Estación. Sistema Meteorológico Nacional, La Soledad 03031, La Paz, México. 2015. Available online: http://smn.cna.gob.mx/ (accessed on 2 July 2022).
- SG Secretaría de Gobernación. Diario Oficial de la Federación 02/08/1978. 1978. Available online: http://dof.gob.mx/nota_detalle.php?codigo=4720542&fecha=02/08/1978 (accessed on 9 January 2020).
- Anderson, D.M. Cysts as factors in Pyrodinium bahamense ecology. In Biology, Epidemiology, and Management of Pyrodinium Red Tides, Proceedings of the Management and Training Workshop, Bandar Seri Begawan, Brunei Darussalam, 23–30 May 1989; Fisheries Department, Ministry of Development and International Center for Living Aquatic Resources Management: Manila, Philippines, 1989. [Google Scholar]
- Cuellar-Martinez, T.; Ruiz-Fernández, A.C.; Sanchez-Cabeza, J.A.; Alonso-Rodríguez, R. Sedimentary record of recent climate impacts on an insular coastal lagoon in the Gulf of California. Quat. Sci. Rev. 2017, 160, 138–149. [Google Scholar] [CrossRef]
- Ruiz-Fernández, A.C.; Hillaire-Marcel, C. 210Pb-derived ages for the reconstruction of terrestrial contaminant history into the Mexican Pacific coast: Potential and limitations. Mar. Pollut. Bull. 2009, 59, 134–145. [Google Scholar] [CrossRef]
- Sanchez-Cabeza, J.A.; Ruiz-Fernández, A.C. 210Pb sediment radiochronology: An integrated formulation and classification of dating models. Geochim. Cosmochim. Acta. 2012, 82, 183–200. [Google Scholar] [CrossRef]
- Sanchez-Cabeza, J.A.; Ruiz-Fernández, A.C.; Ontiveros-Cuadras, J.F.; Pérez Bernal, L.H.; Olid, C. Monte Carlo uncertainty calculation of 210Pb chronologies and accumulation rates of sediments and peat bogs. Quat. Geochronol. 2014, 23, 80–93. [Google Scholar] [CrossRef]
- Ruiz-Fernández, A.C.; Maanan, M.; Sanchez-Cabeza, J.A.; Pérez-Bernal, L.H.; López-Mendoza, P.; Limoges, A. Chronology of recent sedimentation and geochemical characteristics of sediments in Alvarado Lagoon, Veracruz (southwestern Gulf of Mexico). Ciencias Mar. 2014, 40, 291–303. [Google Scholar] [CrossRef]
- Wall, D.; Dale, B. The “hystrichospherid” resting spore of the dinoflagellate Pyrodinium bahamense Plate, 1906. J. Phycol. 1969, 5, 140–149. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Springer: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Strickland, J.; Parsons, T. A Practical Handbook of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada (Bulletin Fisheries Research Board of Canada, 167): Ottawa, Canada, 1972; 310p. [Google Scholar]
- Kawachi, M.; Noël, M.H. Sterilization and sterile technique. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 65–81. [Google Scholar]
- Kremp, A. Effects of cyst resuspension on germination and seeding of two bloom-forming dinoflagellates in the Baltic Sea. Mar. Ecol. Prog. Ser. 2001, 216, 57–66. [Google Scholar] [CrossRef]
- Matsuoka, K.; Joyce, L.B.; Kotani, Y.; Matsuyama, Y. Modern dinoflagellate cysts in hypertrophic coastal waters of Tokyo Bay, Japan. J. Plankton Res. 2003, 25, 1461–1470. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Education Ltd.: Harlow, UK, 2014. [Google Scholar]
- Johnson, V.E. Evidence from marginally significant t statistics. Am. Stat. 2019, 73, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Sombrito, E.Z.; Bulos, A.M.; Sta Maria, E.J.; Honrado, M.C.V.; Azanza, R.V.; Furio, E.F. Application of 210Pb-derived sedimentation rates and dinoflagellate cyst analyses in understanding Pyrodinium bahamense harmful algal blooms in Manila Bay and Malampaya Sound, Philippines. J. Environ. Radioact. 2004, 76, 177–194. [Google Scholar] [CrossRef]
- Villanoy, C.L.; Azanza, R.V.; Altemerano, A.; Casil, A.L. Attempts to model the bloom dynamics of Pyrodinium, a tropical toxic dinoflagellate. Harmful Algae 2006, 5, 156–183. [Google Scholar] [CrossRef]
- Martínez-López, A.; Ulloa-Pérez, A.; Escobedo-Urias, D. First record of vegetative cells of Pyrodinium bahamense (Gonyaulacales: Goniodomataceae) in the Gulf of California. Pac. Sci. 2007, 61, 289–293. [Google Scholar]
- Morquecho, L. Morphology of Pyrodinium bahamense Plate (Dinoflagellata) near Isla San José, Gulf of California, Mexico. Harmful Algae 2008, 7, 664–670. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I.; González-Armas, R. Occurrence of Pyrodinium bahamense var. compressum along the southern coast of the Baja California Peninsula. Mar. Pollut. Bull. 2011, 62, 626–630. [Google Scholar] [CrossRef]
- Onda, D.F.L.; Lluisma, A.O.; Azanza, R.V. Development, morphological characteristics and viability of temporary cysts of Pyrodinium bahamense var. compressum (Dinophyceae) in vitro. Eur. J. Phycol. 2014, 49, 265–275. [Google Scholar] [CrossRef]
- Kremp, A.; Hinners, J.; Klais, R.; Leppänen, A.-P.; Kallio, A. Patterns of vertical cyst distribution and survival in 100-year-old sediment archives of three spring dinoflagellate species from the Northern Baltic Sea. Eur. J. Phycol. 2018, 53, 135–145. [Google Scholar] [CrossRef]
- Ellegaard, M.; Godhe, A.; Ribeiro, S. Time capsules in natural sediment archives-Tracking phytoplankton population genetic diversity and adaptation over multidecadal timescales in the face of environmental change. Evol. Appl. 2018, 11, 11–16. [Google Scholar] [CrossRef]
- Delebecq, G.; Schmidt, S.; Ehrhold, A.; Latimier, M.; Siano, R. Revival of ancient marine dinoflagellates using molecular biostimulation. J. Phycol. 2020, 56, 1077–1089. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J. Cyst germination of two dinoflagellate species from Galicia (NW Spain). Sci. Mar. 1990, 54, 287–291. [Google Scholar]
- Martin, W.R. Chemical processes in estuarine sediments. In Elements of Physical Oceanography: A Derivative of the Encyclopedia of Ocean Sciences, 2nd ed.; Steele, J.H., Thorpe, S.A., Turekian, K.K., Eds.; Academic Press: London, UK, 2009; pp. 539–550. [Google Scholar]
- Pakhomova, S.V.; Hall, P.O.J.; Kononets, M.Y.; Rozanov, A.G.; Tengberg, A.; Vershinin, A.V. Fluxes of iron and manganese across the sediment–water interface under various redox conditions. Mar. Chem. 2007, 107, 319–331. [Google Scholar] [CrossRef]
- Newton, A.; Icely, J.; Cristina, S.; Perillo, G.M.E.; Turner, R.E.; Ashan, D.; Cragg, S.; Luo, Y.; Tu, C.; Li, Y.; et al. Anthropogenic, Direct Pressures on Coastal Wetlands. Front. Ecol. Evol. 2020, 8, 144. [Google Scholar] [CrossRef]
- Moore, S.K.; Trainer, V.L.; Mantua, N.J.; Parker, M.S.; Laws, E.A.; Backer, L.C.; Fleming, L.E. Impacts of climate variability and future climate change on harmful algal blooms and human health. Environ. Health 2008, 7, S4. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.J.; Eriksen, R.; Hallegraeff, G.M.; Rochester, W.; Pitcher, G.C.; Burford, M. Observing changes in harmful algal blooms over time: Long-term observations for studying impacts of climate change (Chapter 2). In Guidelines for the Study of Climate Change Effects on HABs; Wells, M.L., Burford, M., Kremp, A., Montresor, M., Pitcher, G.C., Eds.; GlobalHAB, UNESCO-IOC/SCOR (IOC Manuals and Guides no 88): Paris, France, 2021; pp. 13–35. [Google Scholar]
- Hidalgo-González, R.M.; Álvarez-Borrego, S. Total and new production in the Gulf of California estimated from ocean color data from the satellite sensor SeaWIFS. Deep-Sea Res. II Trop. Stud. Oceanogr. 2004, 51, 739–752. [Google Scholar] [CrossRef]
- Álvarez-Borrego, S. Phytoplankton biomass and production in the Gulf of California: A review. Bot. Mar. 2012, 55, 119–128. [Google Scholar] [CrossRef]
- Lim, P.T.; Leaw, C.P.; Kobiyama, A.; Ogata, T. Growth and toxin production of tropical Alexandrium minutum Halim (Dinophyceae) under various nitrogen to phosphorus ratios. J. Appl. Phycol. 2010, 22, 203–210. [Google Scholar] [CrossRef]
- Natsuike, M.; Oikawa, H.; Matsuno, K.; Yamaguchi, A.; Imai, I. The physiological adaptations and toxin profiles of the toxic Alexandrium fundyense on the eastern Bering Sea and Chukchi Sea shelves. Harmful Algae 2017, 63, 13–22. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuellar-Martinez, T.; Morquecho, L.; Alonso-Rodríguez, R.; Ruiz-Fernández, A.C.; Sanchez-Cabeza, J.-A. Germination of Pyrodinium bahamense Cysts from a Pristine Lagoon in San José Island, Gulf of California: Implications of Long-Term Survival. Phycology 2023, 3, 65-78. https://doi.org/10.3390/phycology3010005
Cuellar-Martinez T, Morquecho L, Alonso-Rodríguez R, Ruiz-Fernández AC, Sanchez-Cabeza J-A. Germination of Pyrodinium bahamense Cysts from a Pristine Lagoon in San José Island, Gulf of California: Implications of Long-Term Survival. Phycology. 2023; 3(1):65-78. https://doi.org/10.3390/phycology3010005
Chicago/Turabian StyleCuellar-Martinez, Tomasa, Lourdes Morquecho, Rosalba Alonso-Rodríguez, Ana Carolina Ruiz-Fernández, and Joan-Albert Sanchez-Cabeza. 2023. "Germination of Pyrodinium bahamense Cysts from a Pristine Lagoon in San José Island, Gulf of California: Implications of Long-Term Survival" Phycology 3, no. 1: 65-78. https://doi.org/10.3390/phycology3010005
APA StyleCuellar-Martinez, T., Morquecho, L., Alonso-Rodríguez, R., Ruiz-Fernández, A. C., & Sanchez-Cabeza, J. -A. (2023). Germination of Pyrodinium bahamense Cysts from a Pristine Lagoon in San José Island, Gulf of California: Implications of Long-Term Survival. Phycology, 3(1), 65-78. https://doi.org/10.3390/phycology3010005