


Seed Priming as an Effective Technique for Enhancing Salinity Tolerance in Plants: Mechanistic Insights and Prospects for Saline Agriculture with a Special Emphasis on Halophytes



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methodology for Literature Search
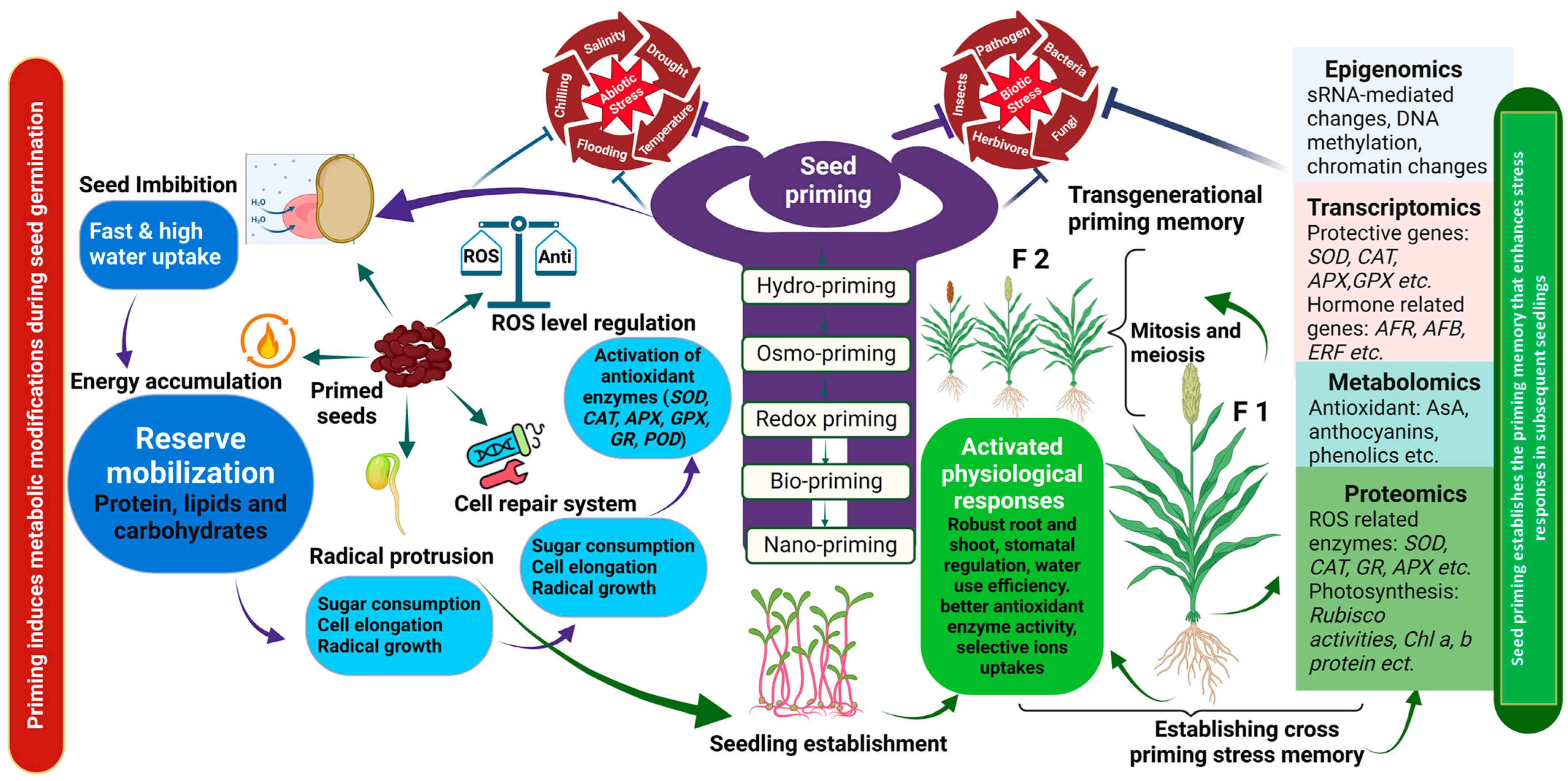
3. General Procedure and Types of Seed Priming
4. Seed Priming and Salinity Tolerance in Halophytes
5. Comparative Analysis of Seed Priming Techniques
6. Mechanistic Insights: Physiochemical and Molecular Footprints
6.1. Physio-Chemical Changes
6.2. Molecular Changes
7. ROS and NO as Potential Signaling Cues of Seed Priming
8. Limitations and Applications of Seed Priming in Saline Agriculture
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, A. Soil Salinity: A Global Threat to Sustainable Development. Soil Use Manag. 2022, 38, 39–67. [Google Scholar] [CrossRef]
- Tarolli, P.; Luo, J.; Park, E.; Barcaccia, G.; Masin, R. Soil Salinization in Agriculture: Mitigation and Adaptation Strategies Combining Nature-Based Solutions and Bioengineering. iScience 2024, 27, 108830. [Google Scholar] [CrossRef] [PubMed]
- Oelviani, R.; Adiyoga, W.; Bakti, I.G.M.Y.; Suhendrata, T.; Malik, A.; Chanifah, C.; Samijan, S.; Sahara, D.; Sutanto, H.A.; Wulanjari, M.E.; et al. Climate Change Driving Salinity: An Overview of Vulnerabilities, Adaptations, and Challenges for Indonesian. Agric. Weather Clim. Soc. 2023, 16, 29–49. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate Change Impacts on Soil Salinity in Agricultural Areas. Eur. J. Soil Sci 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Bridges, E.M.; Oldeman, L.R. Global Assessment of Human-Induced Soil Degradation. Arid Soil Res. Rehabil. 1999, 13, 319–325. [Google Scholar] [CrossRef]
- Butcher, K.; Wick, A.F.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil Salinity: A Threat to Global Food Security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Golla, B. Agricultural Production System in Arid and Semi-Arid Regions. Int. J. Agric. Sci. Food technol. 2021, 7, 234–244. [Google Scholar]
- Aksoy, S.; Yildirim, A.; Gorji, T.; Hamzehpour, N.; Tanik, A.; Sertel, E. Assessing the Performance of Machine Learning Algorithms for Soil Salinity Mapping in Google Earth Engine Platform Using Sentinel-2A and Landsat-8 OLI Data. Adv. Space Res. 2022, 69, 1072–1086. [Google Scholar] [CrossRef]
- Nadathur, S.; Wanasundara, P.; Scanlin, L. Feeding the Globe Nutritious Food in 2050: Obligations and Ethical Choices. In Sustainable Protein Sources Advances for a Healthier Tomorrow; Nadathur, S., Scanlin, L., Wanasundara, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2024; pp. 649–668. [Google Scholar] [CrossRef]
- Ghosh, A.; Kumar, A.; Biswas, G. Exponential Population Growth and Global Food Security: Challenges and Alternatives. In Bioremediation of Emerging Contaminants from Soils: Soil Health Conservation for Improved Ecology and Food Security; Kumar, P., Srivastav, A.L., Chaudhary, V., Hullebusch, E.D., Busquets, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2024; pp. 1–20. [Google Scholar] [CrossRef]
- Siegel, F.R. Food 2050: More Mouths to Feed—Food Availability and Access. In The Earth’s Human Carrying Capacity: Limitations Assessed, Solutions Proposed; Siegel, F.R., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 87–107. [Google Scholar] [CrossRef]
- Grigore, M.N.; Vicente, O. Wild Halophytes: Tools for Understanding Salt Tolerance Mechanisms of Plants and for Adapting Agriculture to Climate Change. Plants 2023, 12, 221. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of Halophytes: Multiple Origins of Salt Tolerance in Land Plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Hameed, A.; Khan, M.A. Halophytes: Biology and economic potentials. Karachi Univ. J. Sci. 2011, 39, 40–44. [Google Scholar]
- Qadir, M.; Tubeileh, A.; Akhtar, J.; Larbi, A.; Minhas, P.S.; Khan, M.A. Productivity Enhancement of Salt-Affected Environments through Crop Diversification. Land Degrad. Dev. 2008, 19, 429–453. [Google Scholar] [CrossRef]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte Agriculture: Success Stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Mirza, S.R.; Ilyas, N.; Batool, N. Seed Priming Enhanced Seed Germination Traits of Wheat under Water, Salt and Heat Stress. Pure Appl. Biol. 2021, 4, 650–658. [Google Scholar] [CrossRef]
- Hussain, S.; Nisar, F.; Bhatt, A.; Nielsen, B.L.; Gul, B.; Hameed, A. Redox Priming Alleviates Dormancy and Improves Salinity Tolerance of Seeds and Seedlings of Medicinal Halophyte Zygophyllum simplex L. Appl. Res. Med. Aromat. Plants 2022, 30, 100384. [Google Scholar] [CrossRef]
- Hameed, A.; Hussain, S.; Rasheed, A.; Ahmed, M.Z.; Abbas, S. Exploring the Potentials of Halophytes in Addressing Climate Change-Related Issues: A Synthesis of Their Biological, Environmental, and Socioeconomic Aspects. World 2024, 5, 36–57. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination Strategies of Halophyte Seeds under Salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Schabes, F.I.; Sigstad, E.E. Calorimetric Studies of Quinoa (Chenopodium quinoa Willd.) Seed Germination under Saline Stress Conditions. Thermochim. Acta 2005, 428, 71–75. [Google Scholar] [CrossRef]
- Al-Khateeb, S.A. Effect of Salinity and Temperature on Germination, Growth and Ion Relations of Panicum turgidum Forssk. Bioresour. Technol. 2006, 97, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Ahmed, M.; Gulzar, S.; Gul, B.; Alam, J.; Hegazy, A.; Alatar, A.; Khan, M.A. Seed germination and recovery responses of suaeda heterophylla to abiotic stresses. Pak. J. Bot. 2013, 45, 1649–1656. [Google Scholar] [CrossRef]
- Amir, M.; Prasad, D.; Khan, F.A.; Khan, A.; Ahmad, B.; Astha, A. Seed Priming: An Overview of Techniques, Mechanisms, and Applications. Plant Sci. Today 2024, 11, 553–563. [Google Scholar] [CrossRef]
- Hernández, J.A.; Barba-Espín, G.; Díaz-Vivancos, P. Seed Priming Technology: Current Perspectives. Seeds 2024, 3, 540–543. [Google Scholar] [CrossRef]
- Hussain, S.; Nisar, F.; Gul, B.; Hameed, A. Seed Priming with Melatonin Improved Salinity Tolerance of Halophytes during Early Life-Cycle Stages. Plant Growth Regul. 2024, 103, 351–368. [Google Scholar] [CrossRef]
- Jarrar, H.; El-Keblawy, A.; Albawab, M.; Ghenai, C.; Sheteiwy, M. Seed Priming as a Promising Technique for Sustainable Restoration of Dryland. Restor. Ecol. 2024, 32, e14182. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed Priming as a Cost Effective Technique for Developing Plants with Cross Tolerance to Salinity Stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Diya, A.; Beena, R.; Jayalekshmy, V.G. Physiological, Biochemical and Molecular Mechanisms of Seed Priming: A Review. Legume Res. Int. J. 2024, 47, 159–166. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed Priming to Alleviate Salinity Stress in Germinating Seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Tamindžić, G.; Ignjatov, M.; Miljaković, D.; Červenski, J.; Milošević, D.; Nikolić, Z.; Vasiljević, S. Seed Priming Treatments to Improve Heat Stress Tolerance of Garden Pea (Pisum sativum L.). Agriculture 2023, 13, 439. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Gao, G.; Ali, I.; Wu, X.; Tang, M.; Chen, L.; Jiang, L.; Liang, T. Effects of Various Seed Priming on Morphological, Physiological, and Biochemical Traits of Rice under Chilling Stress. Front. Plant Sci. 2023, 14, 1146285. [Google Scholar] [CrossRef] [PubMed]
- Devika, O.S.; Singh, S.; Sarkar, D.; Barnwal, P.; Suman, J.; Rakshit, A. Seed Priming: A Potential Supplement in Integrated Resource Management Under Fragile Intensive Ecosystems. Front. Sustain. Food Syst. 2021, 5, 654001. [Google Scholar] [CrossRef]
- Wang, J.; Tao, M.; Xu, L.; Fan, N.; Zhao, C.; Xiao, Z.; Wang, Z. Chitosan Nanocarriers Loaded with Salicylic Acid for Controlling Fall Armyworm (Spodoptera frugiperda) and Alleviating Oxidative Stress in Maize Plants. Environ. Sci. Nano 2023, 10, 3295–3306. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Bhat, J.A.; Antoniou, C.; Kandhol, N.; Singh, V.P.; Fernie, A.R.; Fotopoulos, V. Redox Regulation by Priming Agents Toward a Sustainable Agriculture. Plant Cell Physiol. 2024, 65, 1087–1102. [Google Scholar] [CrossRef]
- Khondoker, M.; Mandal, S.; Gurav, R.; Hwang, S. Freshwater Shortage, Salinity Increase, and Global Food Production: A Need for Sustainable Irrigation Water Desalination—A Scoping Review. Earth 2023, 4, 223–240. [Google Scholar] [CrossRef]
- Sumbal, S.; Ali, A.; Nasser Binjawhar, D.; Ullah, Z.; Eldin, S.M.; Iqbal, R.; Sher, H.; Ali, I. Comparative Effects of Hydropriming and Iron Priming on Germination and Seedling Morphophysiological Attributes of Stay-Green Wheat. ACS Omega 2023, 8, 23078–23088. [Google Scholar] [CrossRef] [PubMed]
- Jisha, K.C.; Puthur, J.T. Seed Halopriming Outdo Hydropriming in Enhancing Seedling Vigor and Osmotic Stress Tolerance Potential of Rice Varieties. J. Crop Sci. Biotechnol. 2014, 17, 209–219. [Google Scholar] [CrossRef]
- Jiang, B.; Wang, L.; Xu, C.; Yan, M. Hydropriming Enhances the Germination of Aged Ultra-Dry Wheat Seeds. Seed Sci. Technol. 2020, 48, 57–63. [Google Scholar] [CrossRef]
- Mosavian, S.N.; Esmaeilzade-Moridani, M. Effect of Hydro-Priming Duration on Germination and Early Seedling Growth of Rapeseed under Salinity Stress. Afr. J. Agric. Res. 2016, 11, 4395–4400. [Google Scholar] [CrossRef]
- Akter, L.; Fakir, O.A.; Alam, M.K.; Islam, M.U.; Chakraborti, P.; Alam, M.J.; Rashid, M.H.; Begum, M.; Kader, M.A. Amelioration of Salinity Stress in Maize Seed Germination and Seedling Growth Attributes through Seed Priming. Open J. Soil Sci. 2018, 8, 137–146. [Google Scholar] [CrossRef]
- Matias, J.R.; Torres, S.B.; Leal, C.C.P.; Leite, M.D.S.; Carvalho, S.M.C. Hydropriming as Inducer of Salinity Tolerance in Sunflower Seeds. Rev. Bras. Eng. Agríc. Ambient. 2018, 22, 255–260. [Google Scholar] [CrossRef]
- Farahani, H.A.; Maroufi, K. Effect of Hydropriming on Seedling Vigour in Basil (Ocimum basilicum L.) under Salinity Conditions. Adv. Environ. Biol. 2011, 5, 828–834. [Google Scholar]
- Corbineau, F.; Taskiran-Özbingöl, N.; El-Maarouf-Bouteau, H. Improvement of Seed Quality by Priming: Concept and Biological Basis. Seeds 2023, 2, 101–115. [Google Scholar] [CrossRef]
- Sadeghi, H.; Khazaei, F.; Yari, L.; Sheidaei, S. Effect of seed osmopriming on seed germination behavior and vigor of soybean (Glycine max L.). J. Agric. Biol. Sci. 2011, 6, 39–43. [Google Scholar]
- Chen, K.; Arora, R. Dynamics of the Antioxidant System during Seed Osmopriming, Post-Priming Germination, and Seedling Establishment in Spinach (Spinacia oleracea). Plant Sci. 2011, 180, 212–220. [Google Scholar] [CrossRef]
- Mauromicale, G.; Cavallaro, V. Effects of Seed Osmopriming on Germination of Tomato at Different Water Potential. Seed Sci. Technol. 1995, 23, 393–403. [Google Scholar]
- Hamidi, R.; Pirasteh-Anosheh, H.; Izadi, M. Effect of Seed Halo-Priming Compared with Hydro-Priming on Wheat Germination and Growth. Intl. J. Agron. Plant Prod. 2013, 4, 1611–1615. [Google Scholar]
- Tania, S.S.; Rhaman, M.S.; Hossain, M.M. Hydro-Priming and Halo-Priming Improve Seed Germination, Yield and Yield Contributing Characters of Okra (Abelmoschus esculentus L.). Trop. Plant Res. 2020, 7, 86–93. [Google Scholar] [CrossRef]
- Afzal, I.; Ashraf, S.; Qasim, M.; Basra, S.M.A.; Shahid, M. Does Halopriming Improve G ermination and Seedling Vigour in Marigold (Tagetes spp.). Seed Sci. Technol. 2009, 37, 436–445. [Google Scholar] [CrossRef]
- Ulfat, A.; Majid, S.A.; Hameed, A. Hormonal seed Priming Improves Wheat (Triticum aestivum L.) Field Performance under Drought and Non-stress Conditions. Pak. J. Bot. 2017, 49, 1239–1253. [Google Scholar]
- Sneideris, L.C.; Gavassi, M.A.; Campos, M.L.; D’Amico-Damião, V.; Carvalho, R.F. Effects of Hormonal Priming on Seed Germination of Pigeon Pea under Cadmium Stress. An. Acad. Bras. Cienc. 2015, 87, 1847–1852. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Noor, M.A.; Bakhtavar, M.A.; Ahmad, A.; Haq, Z. Improvement of Spring Maize Performance through Physical and Physiological Seed Enhancements. Seed Sci. Technol. 2015, 43, 238–249. [Google Scholar] [CrossRef]
- Khan, H.A.; Pervez, M.A.; Ayub, C.M.; Ziaf, K.; Balal, R.M.; Shahid, M.A.; Akhtar, N. Hormonal Priming Alleviates Salt Stress in Hot Pepper (Capsicum annuum L.). Soil Environ. 2009, 28, 130–135. [Google Scholar]
- Silva, P.C.C.; Azevedo Neto, A.D.D.; Gheyi, H.R.; Ribas, R.F.; Silva, C.R.D.R.; Cova, A.M.W. Salt Tolerance Induced by Hydrogen Peroxide Priming on Seed Is Related to Improvement of Ion Homeostasis and Antioxidative Defense in Sunflower Plants. J. Plant Nutr. 2021, 44, 1207–1221. [Google Scholar] [CrossRef]
- Hemalatha, G.; Renugadevi, J.; Eevera, T. Studies on Seed Priming with Hydrogen Peroxide for Mitigating Salt Stress in Rice. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 691–695. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Hernández, J.A.; Diaz-Vivancos, P. Role of H2O2 in Pea Seed Germination. Plant Signal. Behav. 2012, 7, 193–195. [Google Scholar] [CrossRef]
- Farooq, M.; Irfan, M.; Aziz, T.; Ahmad, I.; Cheema, S.A. Seed Priming with Ascorbic Acid Improves Drought Resistance of Wheat. J. Agron. Crop Sci. 2013, 199, 12–22. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic Acid Priming Enhances Seed Germination and Seedling Growth of Winter Wheat under Low Temperature Due to Late Sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef]
- Khan, I.; Lubna; Asaf, S.; Bilal, S.; Alamri, S.S.; Jan, R.; Asif, S.; Kim, K.M.; AL-Harrasi, A. Enhanced Growth and Stress Tolerance in Barley (Hordeum vulgare) through Biopriming with Aspergillus Niger CSR3: A Promising Approach for Sustainable Agriculture in Saline Environments. Cereal Res. Commun. 2024, 52, 1201–1213. [Google Scholar] [CrossRef]
- Rawat, L.; Bisht, T.S.; Kukreti, A. Potential of Seed Biopriming with Trichoderma in Ameliorating Salinity Stress and Providing Resistance against Leaf Blast Disease in Finger Millet (Eleusine coracana L.). Indian Phytopathol. 2022, 75, 147–164. [Google Scholar] [CrossRef]
- Dief, H.E.S.; Hashem, E.S.A.; Fawzan, S.; El-Sayed, A.S.A. Alleviation of Salt Stress in Triticum aestivum by Biopriming with Phanerochaete chrysosporium. J. Crop Sci. Biotechnol. 2021, 24, 103–116. [Google Scholar] [CrossRef]
- Hadj Brahim, A.; Ben Ali, M.; Daoud, L.; Jlidi, M.; Akremi, I.; Hmani, H.; Feto, N.A.; Ben Ali, M. Biopriming of Durum Wheat Seeds with Endophytic Diazotrophic Bacteria Enhances Tolerance to Fusarium Head Blight and Salinity. Microorganisms 2022, 10, 970. [Google Scholar] [CrossRef]
- Singh, S.; Singh, U.B.; Trivedi, M.; Sahu, P.K.; Paul, S.; Paul, D.; Saxena, A.K. Seed Biopriming with Salt-Tolerant Endophytic Pseudomonas Geniculata-Modulated Biochemical Responses Provide Ecological Fitness in Maize (Zea mays L.) Grown in Saline Sodic Soil. Int. J. Environ. Res. Public Health 2020, 17, 253. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Arshad, U.; Ayub, S.; Jabeen, A.; Saeed, M. Bio-Priming of Seed with Trichoderma sp. and Bacillus sp. Leads to Improved Morpho-Physiological Parameters Related to Drought Tolerance in Maize. J. Integr. Plant Biol. 2023, 1, 43–51. [Google Scholar] [CrossRef]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-Priming as Emerging Seed Priming Technology for Sustainable Agriculture—Recent Developments and Future Perspectives. J. Nanobiotechnol. 2022, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.M.; Baksi, S.; Rani, S.; Jha, A.B.; Dubey, R.S.; Sharma, P. NanoBoost: Maximizing Crop Resilience and Yield via Nanopriming Under Salt Stress. Environ. Exp. Bot. 2024, 226, 105937. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Abdullah Alsahli, A.; Jan, S.; Ahmad, P. Seed Priming with Titanium Dioxide Nanoparticles Enhances Seed Vigor, Leaf Water Status, and Antioxidant Enzyme Activities in Maize (Zea mays L.) under Salinity Stress. J. King Saud Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- Farooq, T.; Akram, M.N.; Hameed, A.; Ahmed, T.; Hameed, A. Nanopriming-Mediated Memory Imprints Reduce Salt Toxicity in Wheat Seedlings by Modulating Physiobiochemical Attributes. BMC Plant Biol. 2022, 22, 540. [Google Scholar] [CrossRef] [PubMed]
- Soni, A.T.; Rookes, J.E.; Arya, S.S. Chitosan Nanoparticles as Seed Priming Agents to Alleviate Salinity Stress in Rice (Oryza sativa L.) Seedlings. Polysaccharides 2023, 4, 129–141. [Google Scholar] [CrossRef]
- El-Badri, A.M.A.; Batool, M.; Mohamed, I.A.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of Salinity Impact on Early Seedling Stage via Nano-Priming Application of Zinc Oxide on Rapeseed (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Ellouzi, H.; Sghayar, S.; Abdelly, C. H2O2 Seed Priming Improves Tolerance to Salinity; Drought and Their Combined Effect More than Mannitol in Cakile Maritima When Compared to Eutrema salsugineum. J. Plant Physiol. 2017, 210, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Zammali, I.; Dabbous, A.; Youssef, S.; Ben Hamed, K. Effects of Chemical Priming on the Germination of the Ornamental Halophyte Lobularia maritima under NaCl Salinity. Seeds 2022, 1, 99–109. [Google Scholar] [CrossRef]
- Ramírez, E.; Chaâbene, Z.; Hernández-Apaolaza, L.; Rekik, M.; Elleuch, A.; de la Fuente, V. Seed Priming to Optimize Germination in Arthrocnemum Moq. BMC Plant Biol. 2022, 22, 527. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Baazaoui, N.; Hammami, S.B.M.; Abdelly, C. Priming-Mediated Tolerance to Biotic and Abiotic Stresses in Halophytes. In Handbook of Halophytes: From Molecules to Ecosystems Towards Biosaline Agriculture; Grigore, M.N., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–26. [Google Scholar] [CrossRef]
- Khaleghi, G. Investigating the Effect of Hydropriming on Germination Components of Atriplex Plant (Atriplex canescens) under Salinity Stress. Greenh. Plant Prod. J. 2024, 1, 47–54. [Google Scholar] [CrossRef]
- Khaleghi, G. The Effect of Hydropriming on Germination Characteristics and Seedling Growth of Nitraria Schoberi Seeds under Salinity Stress. Greenh. Plant Prod. J. 2024, 1, 38–46. [Google Scholar] [CrossRef]
- Lallouche, B.; Kouider, B.H. Effects of Hydropriming, Halopriming, and Hormopriming Seed Treatments on the Subsequent Salt Stress Tolerance of Quinoa (Chenopodium quinoa Willd.) in Algeria. Acta Sci. Pol. Hortoru. 2024, 23, 59–70. [Google Scholar] [CrossRef]
- Ben Youssef, R.; Jelali, N.; Boukari, N.; Albacete, A.; Martinez, C.; Alfocea, F.P.; Abdelly, C. The Efficiency of Different Priming Agents for Improving Germination and Early Seedling Growth of Local Tunisian Barley under Salinity Stress. Plants 2021, 10, 2264. [Google Scholar] [CrossRef] [PubMed]
- Shiri, M.; Rabhi, M.; El Amrani, A.; Abdelly, C. Cross-Tolerance to Abiotic Stresses in Halophytes: Application for Phytoremediation of Organic Pollutants. Acta Physiol. Plant. 2015, 37, 209. [Google Scholar] [CrossRef]
- Moreno, C.; Seal, C.E.; Papenbrock, J. Seed Priming Improves Germination in Saline Conditions for Chenopodium quinoa and Amaranthus caudatus. J Agron. Crop Sci. 2018, 204, 40–48. [Google Scholar] [CrossRef]
- Messedi, D.; Hmidi, D.; Farhani, F.; Zribi, F.; Zorrig, W.; Abdelly, C.; Debez, A. Proline Priming: An Efficient Strategy to Mitigate Salinity Impact at Early Developmental Stages of the Oilseed Halophyte Cakile Maritima. Not. Bot. Horti Agrobot. Cluj-Napoca 2024, 52, 13458. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Cross-Talk between Nitric Oxide, Hydrogen Peroxide and Calcium in Salt-Stressed Chenopodium quinoa Willd. At Seed Germination Stage. Plant Physiol. Biochem. 2020, 154, 657–664. [Google Scholar] [CrossRef]
- Nimac, A.; Lazarević, B.; Petek, M.; Vidak, M.; Šatović, Z.; Carović-Stanko, K. Effects of Salinity and Seed Priming on Germination of Sea Fennel (Crithmum maritimum L.). Agric. Conspec. Sci. 2018, 83, 181–185. [Google Scholar]
- Khan, M.A.; Ahmed, M.Z.; Hameed, A. Effect of Sea Salt and L-Ascorbic Acid on the Seed Germination of Halophytes. J. Arid Environ. 2006, 67, 535–540. [Google Scholar] [CrossRef]
- Aizaz, M.; Ahmad, W.; Asaf, S.; Khan, I.; Saad Jan, S.; Salim Alamri, S.; Bilal, S.; Jan, R.; Kim, K.M.; Al-Harrasi, A. Characterization of the Seed Biopriming, Plant Growth-Promoting and Salinity-Ameliorating Potential of Halophilic Fungi Isolated from Hypersaline Habitats. Int. J. Mol. Sci. 2023, 24, 4904. [Google Scholar] [CrossRef] [PubMed]
- Dutta, B.; Datta, A.; Dey, A.; Ghosh, A.K.; Bandopadhyay, R. Establishment of Seed Biopriming in Salt Stress Mitigation of Rice Plants by Mangrove Derived Bacillus sp. Biocatal. Agric. Biotechnol. 2023, 48, 102626. [Google Scholar] [CrossRef]
- Meinzer, M.; Ahmad, N.; Nielsen, B.L. Halophilic Plant-Associated Bacteria with Plant-Growth-Promoting Potential. Microorganisms 2023, 11, 2910. [Google Scholar] [CrossRef]
- Al-Barakah, F.N.I.; Sohaib, M. Evaluating the Germination Response of Chenopodium quinoa Seeds to Bacterial Inoculation under Different Germination Media and Salinity Conditions. Seed Sci. Technol. 2019, 47, 161–169. [Google Scholar] [CrossRef]
- Lee, J.H.J.; Kasote, D.M. Nano-Priming for Inducing Salinity Tolerance, Disease Resistance, Yield Attributes, and Alleviating Heavy Metal Toxicity in Plants. Plants 2024, 13, 446. [Google Scholar] [CrossRef]
- Munir, N.; Hanif, M.; Dias, D.A.; Abideen, Z. The Role of Halophytic Nanoparticles towards the Remediation of Degraded and Saline Agricultural Lands. Environ. Sci. Pollut. Res. 2021, 28, 60383–60405. [Google Scholar] [CrossRef]
- Hanif, M.; Munir, N.; Abideen, Z.; Dias, D.A.; Hessini, K.; El-Keblawy, A. Enhancing Tomato Plant Growth in a Saline Environment through the Eco-Friendly Synthesis and Optimization of Nanoparticles Derived from Halophytic Sources. Environ. Sci. Pollut. Res. 2023, 30, 118830–118854. [Google Scholar] [CrossRef]
- Ramzan, M.; Parveen, M.; Naz, G.; Sharif, H.M.A.; Nazim, M.; Aslam, S.; Hussain, A.; Rahimi, M.; Alamer, K.H. Enhancing Physio-Biochemical Characteristics in Okra Genotypes through Seed Priming with Biogenic Zinc Oxide Nanoparticles Synthesized from Halophytic Plant Extracts. Sci. Rep. 2024, 14, 23753. [Google Scholar] [CrossRef]
- Ashraf Ganjouii, F.; Nasibi, F.; Manoochehri Kalantari, K.; Ahmadi Mousavi, E. Effect of Seed Priming with Selenium Nanoparticles and Plant Growth Promoting Rhizobacteria on Improving Quinoa Seedling Growth under Salinity Stress. J. Plant Process Funct. 2023, 11, 65–74. [Google Scholar]
- Elouaer, M.A.; Hannachi, C. Seed Priming to Improve Germination and Seedling Growth of Safflower (Carthamus tinctorius) under Salt Stress. Eurasia J. Biosci. 2012, 6, 76–84. [Google Scholar] [CrossRef]
- Toorop, P.E.; van Aelst, A.C.; Hilhorst, H.W.M. Endosperm Cap Weakening and Endo-β-Mannanase Activity during Priming of Tomato (Lycopersicon esculentum Cv. Moneymaker) Seeds Are Initiated upon Crossing a Threshold Water Potential. Seed Sci. Res. 1998, 8, 483–492. [Google Scholar] [CrossRef]
- Nagarajan, S.; Pandita, V.K.; Joshi, D.K.; Sinha, J.P.; Modi, B.S. Characterization of Water Status in Primed Seeds of Tomato (Lycopersicon esculentum Mill.) by Sorption Properties and NMR Relaxation Times. Seed Sci. Res. 2005, 15, 99–111. [Google Scholar] [CrossRef]
- Galhaut, L.; de Lespinay, A.; Walker, D.J.; Bernal, M.P.; Correal, E.; Lutts, S. Seed Priming of Trifolium Repens L. Improved Germination and Early Seedling Growth on Heavy Metal-Contaminated Soil. Water Air Soil Pollut. 2014, 225, 1905. [Google Scholar] [CrossRef]
- Liu, Y.; van der Burg, W.J.; Aartse, J.W.; van Zwol, R.A.; Jalink, H.; Bino, R.J. X-Ray Studies on Changes in Embryo and Endosperm Morphology during Priming and Imbibition of Tomato Seeds. Seed Sci. Res. 1993, 3, 171–178. [Google Scholar] [CrossRef]
- Oluoch, M.O.; Welbaum, G.E. Viability and Vigor of Osmotically Primed Muskmelon Seeds After Nine Years of Storage. JASHS 1996, 121, 408–413. [Google Scholar] [CrossRef]
- Kubala, S.; Garnczarska, M.; Wojtyla, Ł.; Clippe, A.; Kosmala, A.; Żmieńko, A.; Lutts, S.; Quinet, M. Deciphering Priming-Induced Improvement of Rapeseed (Brassica napus L.) Germination through an Integrated Transcriptomic and Proteomic Approach. Plant Sci. 2015, 231, 94–113. [Google Scholar] [CrossRef]
- Chojnowski, M.; Corbineau, F.; Côme, D. Physiological and Biochemical Changes Induced in Sunflower Seeds by Osmopriming and Subsequent Drying, Storage and Aging. Seed Sci. Res. 1997, 7, 323–332. [Google Scholar] [CrossRef]
- Dahal, P.; Kim, N.S.; Bradford, K.J. Respiration and Germination Rates of Tomato Seeds at Suboptimal Temperatures and Reduced Water Potentials. J. Exp. Bot. 1996, 47, 941–947. [Google Scholar] [CrossRef]
- Ozbingol, N.; Corbineau, F.; Côme, D. Responses of Tomato Seeds to Osmoconditioning as Related to Temperature and Oxygen. Seed Sci. Res. 1998, 8, 377–384. [Google Scholar] [CrossRef]
- Corbineau, F.; Özbingöl, N.; Vinel, D.; Côme, D. Improvement of Tomato Seed Germination by Osmopriming as Related to Energy Metabolism. In Seed Biology: Advances and Applications. Proceedings of the Sixth International Workshop on Seeds, Merida, Mexico, 1999; CABI Books: Wallingford, UK, 2000; pp. 467–476. [Google Scholar] [CrossRef]
- Ashraf, M.; Bray, C.M. DNA Synthesis in Osmoprimed Leek (Allium porrum L.) Seeds and Evidence for Repair and Replication. Seed Sci. Res. 1993, 3, 15–23. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M.; Jamil, A.; Ur-Rehman, S. Does Seed Priming Induce Changes in the Levels of Some Endogenous Plant Hormones in Hexaploid Wheat Plants Under Salt Stress? J. Integr. Plant Biol. 2006, 48, 181–189. [Google Scholar] [CrossRef]
- Nakaune, M.; Hanada, A.; Yin, Y.G.; Matsukura, C.; Yamaguchi, S.; Ezura, H. Molecular and Physiological Dissection of Enhanced Seed Germination Using Short-Term Low-Concentration Salt Seed Priming in Tomato. Plant Physiol. Biochem. 2012, 52, 28–37. [Google Scholar] [CrossRef]
- Nie, L.; Song, S.; Yin, Q.; Zhao, T.; Liu, H.; He, A.; Wang, W. Enhancement in Seed Priming-Induced Starch Degradation of Rice Seed Under Chilling Stress via GA-Mediated α-Amylase Expression. Rice 2022, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Elkholy, S.F.; Almutairi, Z.M. Effect of Priming With Gibberellin On Germination and Expression of Ga Biosynthesis Gene Of Tomato (Solanum lycopersicum L.) Seedlings Under Salt Stress. Int. J. Bio-Technol. Res. 2018, 8, 11–22. [Google Scholar] [CrossRef]
- Mukasa, Y.; Takahashi, H.; Taguchi, K.; Ogata, N.; Okazaki, K.; Tanaka, M. Accumulation of Soluble Sugar in True Seeds by Priming of Sugar Beet Seeds and the Effects of Priming on Growth and Yield of Drilled Plants. Plant Prod. Sci. 2003, 6, 74–82. [Google Scholar] [CrossRef]
- Verma, J.; Srivastava, A.K. Physiological Basis of Salt Stress Resistance in Pigeon Pea (Cajanus cajan L.). Pre-sowing seed soaking treatment in regulating early seedling metabolism during seed germination. Plant Physiol. Biochem. 1998, 25, 89–94. [Google Scholar]
- Li, X.; Zhang, L. SA and PEG-Induced Priming for Water Stress Tolerance in Rice Seedling. In Information Technology and Agricultural Engineering; Zhu, E., Sambath, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 881–887. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Lokhande, V.H.; Patade, V.Y.; Suprasanna, P.; Sjahril, R.; D’Souza, S.F. Comparative Evaluation of Hydro, Chemo, and Hormonal-Priming Methods for Imparting Salt and PEG Stress Tolerance in Indian Mustard (Brassicajuncea L.). Acta Physiol. Plant. 2010, 32, 1135–1144. [Google Scholar] [CrossRef]
- Zhang, F.; Yu, J.; Johnston, C.R.; Wang, Y.; Zhu, K.; Lu, F.; Zhang, Z.; Zou, J. Seed Priming with Polyethylene Glycol Induces Physiological Changes in Sorghum (Sorghum bicolor L. Moench) Seedlings under Suboptimal Soil Moisture Environments. PLoS ONE 2015, 10, e0140620. [Google Scholar] [CrossRef]
- Salama, K.H.A.; Mansour, M.M.F.; Hassan, N.S. Choline Priming Improves Salt Tolerance in Wheat (Triticum aestivum L.). Aust. J. Basic Appl. Sci. 2011, 5, 126–132. [Google Scholar]
- Azooz, M.M. Salt Stress Mitigation by Seed Priming with Salicylic Acid in Two Faba Bean Genotypes Differing in Salt Tolerance. Int. J. Agric. Biol. 2009, 11, 343–350. [Google Scholar]
- Alam, M.U.; Fujita, M.; Nahar, K.; Rahman, A.; Anee, T.I.; Masud, A.A.C.; Amin, A.K.M.R.; Hasanuzzaman, M. Seed Priming Upregulates Antioxidant Defense and Glyoxalase Systems to Conferring Simulated Drought Tolerance in Wheat Seedlings. Plant Stress 2022, 6, 100120. [Google Scholar] [CrossRef]
- Nawaz, A.; Amjad, M.; Jahangir, M.M.; Khan, S.M.; Cui, H.; Hu, J. Induction of Salt Tolerance in Tomato (Lycopersicon esculentum Mill.) Seeds through Sand Priming. Aust. J. Crop Sci. 2012, 6, 1199–1203. [Google Scholar]
- Ellouzi, H.; Oueslati, S.; Hessini, K.; Rabhi, M.; Abdelly, C. Seed-Priming with H2O2 Alleviates Subsequent Salt Stress by Preventing ROS Production and Amplifying Antioxidant Defense in Cauliflower Seeds and Seedlings. Sci. Hortic. 2021, 288, 110360. [Google Scholar] [CrossRef]
- Ali, Q.; Daud, M.K.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F.; et al. Seed Priming by Sodium Nitroprusside Improves Salt Tolerance in Wheat (Triticum aestivum L.) by Enhancing Physiological and Biochemical Parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of Salinity Stress on Vicia faba Plants via Seed Priming with Melatonin. Acta Biol. Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Meena, D.S. Effect of Seed Bio-Priming and N Doses under Varied Soil Type on Nitrogen Use Efficiency (NUE) of Wheat (Triticum aestivum L.) under Greenhouse Conditions. Biocatal. Agric. Biotechnol. 2016, 6, 68–75. [Google Scholar] [CrossRef]
- Sen, A.; Johnson, R.; Puthur, J.T. Seed Priming: A Cost-effective Strategy to Impart Abiotic Stress Tolerance. In Plant Performance Under Environmental Stress Hormones, Biostimulants and Sustainable Plant Growth Management; Husen, A., Ed.; Springer: Cham, Switzerland, 2021; pp. 459–480. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.; Dong, Y.; Zhang, F.; He, Q.; Chen, J.; Zhu, S.; Zhao, T. Seed Priming with Melatonin Improves Salt Tolerance in Cotton through Regulating Photosynthesis, Scavenging Reactive Oxygen Species and Coordinating with Phytohormone Signal Pathways. Ind. Crops Prod. 2021, 169, 113671. [Google Scholar] [CrossRef]
- Dos Santos Araújo, G.; de Oliveira Paula-Marinho, S.; de Paiva Pinheiro, S.K.; de Castro Miguel, E.; de Sousa Lopes, L.; Camelo Marques, E.; de Carvalho, H.H.; Gomes-Filho, E. H2O2 Priming Promotes Salt Tolerance in Maize by Protecting Chloroplasts Ultrastructure and Primary Metabolites Modulation. Plant Sci. 2021, 303, 110774. [Google Scholar] [CrossRef]
- Liu, J.; Hasanuzzaman, M.; Wen, H.; Zhang, J.; Peng, T.; Sun, H.; Zhao, Q. High Temperature and Drought Stress Cause Abscisic Acid and Reactive Oxygen Species Accumulation and Suppress Seed Germination Growth in Rice. Protoplasma 2019, 256, 1217–1227. [Google Scholar] [CrossRef]
- Lanteri, S.; Saracco, F.; Kraak, H.L.; Bino, R.J. The Effects of Priming on Nuclear Replication Activity and Germination of Pepper (Capsicum annuum) and Tomato (Lycopersicon esculentum) Seeds. Seed Sci. Res. 1994, 4, 81–87. [Google Scholar] [CrossRef]
- Ozbingöl, N.; Corbineau, F.; Groot, S.P.C.; Bino, R.J.; Côme, D. Activation of the Cell Cycle in Tomato (Lycopersicon esculentum Mill.) Seeds during Osmoconditioning as Related to Temperature and Oxygen. Ann. Bot. 1999, 84, 245–251. [Google Scholar] [CrossRef]
- Bino, R.J.; Vries, J.N.D.; Kraak, H.L.; Pijlen, J.G.V. Flow Cytometric Determination of Nuclear Replication Stages in Tomato Seeds during Priming and Germination. Ann. Bot. 1992, 69, 231–236. [Google Scholar] [CrossRef]
- Sivritepe, H. The Effect of Priming Treatments on the Viability and Accumulation of Chromosomal Damage in Aged Pea Seeds. Ann. Bot. 1995, 75, 165–171. [Google Scholar] [CrossRef]
- Varier, A.; Vari, A.K.; Dadlani, M. The Subcellular Basis of Seed Priming. Curr. Sci. 2010, 99, 450–456. [Google Scholar]
- Sharma, S.N.; Maheshwari, A. Expression Patterns of DNA Repair Genes Associated with Priming Small and Large Chickpea (Cicer arietinum) Seeds. Seed Sci. Technol. 2015, 43, 250–261. [Google Scholar] [CrossRef]
- Hussain, S.; Zhu, C.; Huang, J.; Huang, J.; Zhu, L.; Cao, X.; Nanda, S.; Khaskheli, M.A.; Liang, Q.; Kong, Y.; et al. Ethylene Response of Salt Stressed Rice Seedlings Following Ethephon and 1-Methylcyclopropene Seed Priming. Plant Growth Regul. 2020, 92, 219–231. [Google Scholar] [CrossRef]
- Matsunami, M.; Hayashi, H.; Murai-Hatano, M.; Ishikawa-Sakurai, J. Effect of Hydropriming on Germination and Aquaporin Gene Expression in Rice. Plant Growth Regul. 2022, 97, 263–270. [Google Scholar] [CrossRef]
- Chen, K.; Fessehaie, A.; Arora, R. Aquaporin Expression during Seed Osmopriming and Post-Priming Germination in Spinach. Biol. Plant 2013, 57, 193–198. [Google Scholar] [CrossRef]
- Soeda, Y.; Konings, M.C.J.M.; Vorst, O.; van Houwelingen, A.M.M.L.; Stoopen, G.M.; Maliepaard, C.A.; Kodde, J.; Bino, R.J.; Groot, S.P.C.; van der Geest, A.H.M. Gene Expression Programs during Brassica oleracea Seed Maturation, Osmopriming, and Germination Are Indicators of Progression of the Germination Process and the Stress Tolerance Level. Plant Physiol. 2005, 137, 354–368. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, H.; Yangzong, Z.; Gardea-Torresdey, J.L.; White, J.C.; Zhao, L. Seed Priming with Reactive Oxygen Species-Generating Nanoparticles Enhanced Maize Tolerance to Multiple Abiotic Stresses. Environ. Sci. Technol. 2023, 57, 19932–19941. [Google Scholar] [CrossRef] [PubMed]
- Karalija, E.; Demira, A.; Samardžić, J.; Parić, A.; Dahija, S.; Contaldi, F.; Martinelli, F. RNA Sequencing Indicates Gene Expression Changes in Silene Sendtneri Seeds after Seed Priming with Silicic Acid. Genet. Appl. 2022, 6, 79–88. [Google Scholar] [CrossRef]
- Buitink, J.; Leger, J.J.; Guisle, I.; Vu, B.L.; Wuillème, S.; Lamirault, G.; Bars, A.L.; Meur, N.L.; Becker, A.; Küster, H.; et al. Transcriptome Profiling Uncovers Metabolic and Regulatory Processes Occurring during the Transition from Desiccation-Sensitive to Desiccation-Tolerant Stages in Medicago Truncatula Seeds. Plant J. 2006, 47, 735–750. [Google Scholar] [CrossRef]
- Catusse, J.; Meinhard, J.; Job, C.; Strub, J.M.; Fischer, U.; Pestsova, E.; Westhoff, P.; Van Dorsselaer, A.; Job, D. Proteomics Reveals Potential Biomarkers of Seed Vigor in Sugarbeet. Proteomics 2011, 11, 1569–1580. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Priming Memory Invokes Seed Stress-Tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Gamir, J.; Sánchez-Bel, P.; Flors, V. Molecular and Physiological Stages of Priming: How Plants Prepare for Environmental Challenges. Plant Cell Rep. 2014, 33, 1935–1949. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Priming Crops for the Future: Rewiring Stress Memory. Trends Plant Sci. 2022, 27, 699–716. [Google Scholar] [CrossRef]
- Louis, N.; Dhankher, O.P.; Puthur, J.T. Seed Priming Can Enhance and Retain Stress Tolerance in Ensuing Generations by Inducing Epigenetic Changes and Trans-Generational Memory. Physiol. Plant. 2023, 175, e13881. [Google Scholar] [CrossRef]
- Sen, A.; Puthur, J.T. Seed Priming-Induced Physiochemical and Molecular Events in Plants Coupled to Abiotic Stress Tolerance: An Overview. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Hossain, M.A., Liu, F., Burritt, D.J., Fujita, M., Huang, B., Eds.; Elsevier International Publishing: Amsterdam, The Netherlands, 2020; pp. 303–316. [Google Scholar] [CrossRef]
- Hameed, A.; Hussain, T.; Gulzar, S.; Aziz, I.; Gul, B.; Khan, M.A. Salt Tolerance of a Cash Crop Halophyte Suaeda Fruticosa: Biochemical Responses to Salt and Exogenous Chemical Treatments. Acta Physiol. Plant. 2012, 34, 2331–2340. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Prashanth, S.R.; Sivaprakash, K.R.; Parida, A. Monitoring Expression Profiles of Antioxidant Genes to Salinity, Iron, Oxidative, Light and Hyperosmotic Stresses in the Highly Salt Tolerant Grey Mangrove, Avicennia Marina (Forsk.) Vierh. by mRNA Analysis. Plant Cell Rep. 2006, 25, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Rai-Kalal, P.; Jajoo, A. Priming with Zinc Oxide Nanoparticles Improve Germination and Photosynthetic Performance in Wheat. Plant Physiol. Biochem. 2021, 160, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Habib, N.; Ali, Q.; Ali, S.; Haider, M.Z.; Javed, M.T.; Khalid, M.; Perveen, R.; Alsahli, A.A.; Alyemeni, M.N. Seed Priming with Sodium Nitroprusside and H2O2 Confers Better Yield in Wheat Under Salinity: Water Relations, Antioxidative Defense Mechanism and Ion Homeostasis. J. Plant Growth Regul. 2021, 40, 2433–2453. [Google Scholar] [CrossRef]
- Molassiotis, A.; Tanou, G.; Diamantidis, G. NO Says More than ‘YES’ to Salt Tolerance. Plant Signal. Behav. 2010, 5, 209–212. [Google Scholar] [CrossRef]
- Sangwan, P.N.S.; Saini, N.; Joshi, G. The Role of Nitric Oxide in Plants under Salt Stress: A Review. J. Plant Biol. Crop Res. 2024, 7, 1098. [Google Scholar]
- Kataria, S.; Anand, A.; Raipuria, R.K.; Kumar, S.; Jain, M.; Watts, A.; Brestic, M. Magnetopriming Actuates Nitric Oxide Synthesis to Regulate Phytohormones for Improving Germination of Soybean Seeds under Salt Stress. Cells 2022, 11, 2174. [Google Scholar] [CrossRef]
- Kataria, S.; Jain, M.; Tripathi, D.K.; Singh, V.P. Involvement of Nitrate Reductase-Dependent Nitric Oxide Production in Magnetopriming-Induced Salt Tolerance in Soybean. Physiol. Plant. 2020, 168, 422–436. [Google Scholar] [CrossRef]
- Wright, B.; Rowse, H.; Whipps, J.M. Microbial Population Dynamics on Seeds during Drum and Steeping Priming. Plant Soil 2003, 255, 631–640. [Google Scholar] [CrossRef]
- Butler, L.H.; Hay, F.R.; Ellis, R.H.; Smith, R.D.; Murray, T.B. Priming and Re-Drying Improve the Survival of Mature Seeds of Digitalis Purpurea during Storage. Ann. Bot. 2009, 103, 1261–1270. [Google Scholar] [CrossRef]
- Chiu, K.Y.; Chen, C.L.; Sung, J.M. Effect of Priming Temperature on Storability of Primed Sh-2 Sweet Corn Seed. Crop Sci. 2002, 42, 1996–2003. [Google Scholar] [CrossRef]
- Hill, H.J.; Cunningham, J.D.; Bradford, K.J.; Taylor, A.G. Primed Lettuce Seeds Exhibit Increased Sensitivity to Moisture Content During Controlled Deterioration. HortScience 2007, 42, 1436–1439. [Google Scholar] [CrossRef]
- Hacisalihoglu, G.; Paine, D.H.; Hilderbrand, M.B.; Khan, A.A.; Taylor, A.G. Embryo Elongation and Germination Rates as Sensitive Indicators of Lettuce Seed Quality: Priming and Aging Studies. HortScience 1999, 34, 1240–1243. [Google Scholar] [CrossRef]
- Ma, H.Y.; Zhao, D.D.; Ning, Q.R.; Wei, J.P.; Li, Y.; Wang, M.M.; Liu, X.L.; Jiang, C.J.; Liang, Z.W. A Multi-Year Beneficial Effect of Seed Priming with Gibberellic Acid-3 (GA3) on Plant Growth and Production in a Perennial Grass, Leymus chinensis. Sci. Rep. 2018, 8, 13214. [Google Scholar] [CrossRef]
- Hussain, S.; Zheng, M.; Khan, F.; Khaliq, A.; Fahad, S.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Benefits of Rice Seed Priming are Offset Permanently by Prolonged Storage and the Storage Conditions. Sci. Rep. 2015, 5, 8101. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hameed, A.; Hussain, S.; Nisar, F.; Rasheed, A.; Shah, S.Z. Seed Priming as an Effective Technique for Enhancing Salinity Tolerance in Plants: Mechanistic Insights and Prospects for Saline Agriculture with a Special Emphasis on Halophytes. Seeds 2025, 4, 14. https://doi.org/10.3390/seeds4010014
Hameed A, Hussain S, Nisar F, Rasheed A, Shah SZ. Seed Priming as an Effective Technique for Enhancing Salinity Tolerance in Plants: Mechanistic Insights and Prospects for Saline Agriculture with a Special Emphasis on Halophytes. Seeds. 2025; 4(1):14. https://doi.org/10.3390/seeds4010014
Chicago/Turabian StyleHameed, Abdul, Sadiq Hussain, Farah Nisar, Aysha Rasheed, and Syed Zaheer Shah. 2025. "Seed Priming as an Effective Technique for Enhancing Salinity Tolerance in Plants: Mechanistic Insights and Prospects for Saline Agriculture with a Special Emphasis on Halophytes" Seeds 4, no. 1: 14. https://doi.org/10.3390/seeds4010014
APA StyleHameed, A., Hussain, S., Nisar, F., Rasheed, A., & Shah, S. Z. (2025). Seed Priming as an Effective Technique for Enhancing Salinity Tolerance in Plants: Mechanistic Insights and Prospects for Saline Agriculture with a Special Emphasis on Halophytes. Seeds, 4(1), 14. https://doi.org/10.3390/seeds4010014