Short-Term Dietary Intervention with Cooked but Not Raw Brassica Leafy Vegetables Increases Telomerase Activity in CD8+ Lymphocytes in a Randomized Human Trial


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
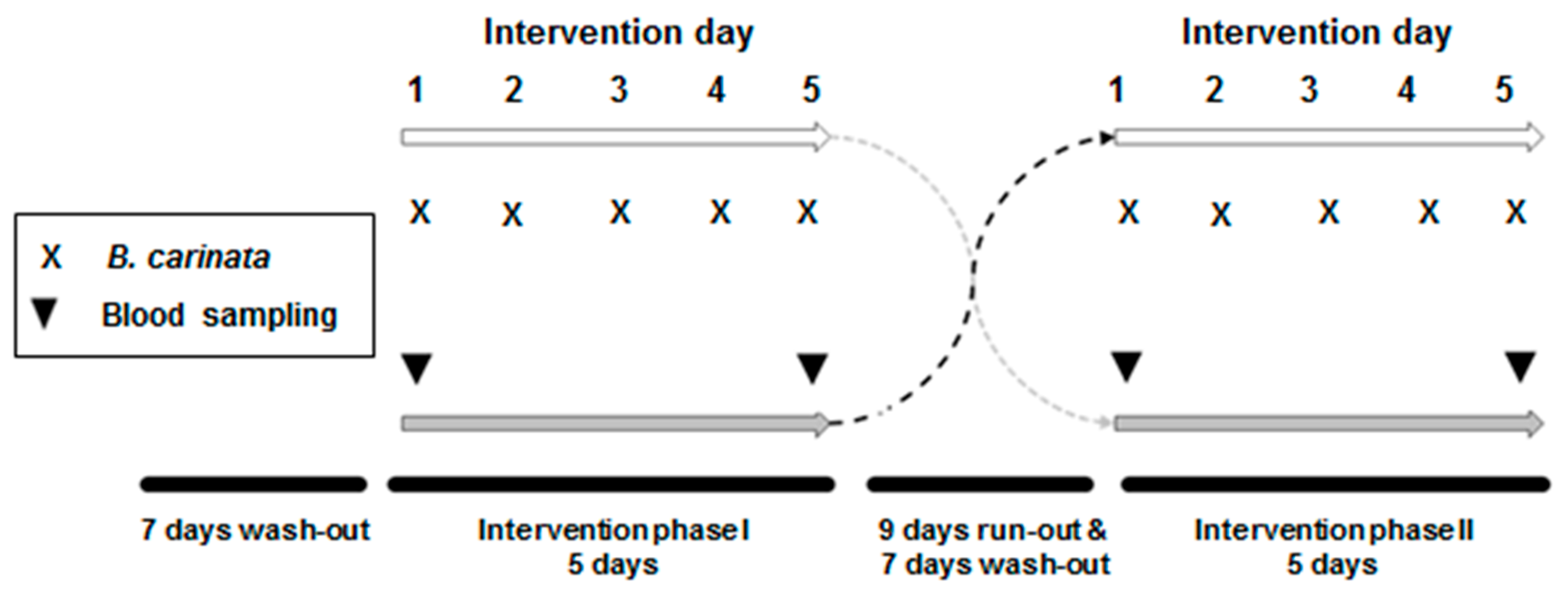
2.2. Subjects and Study Protocol
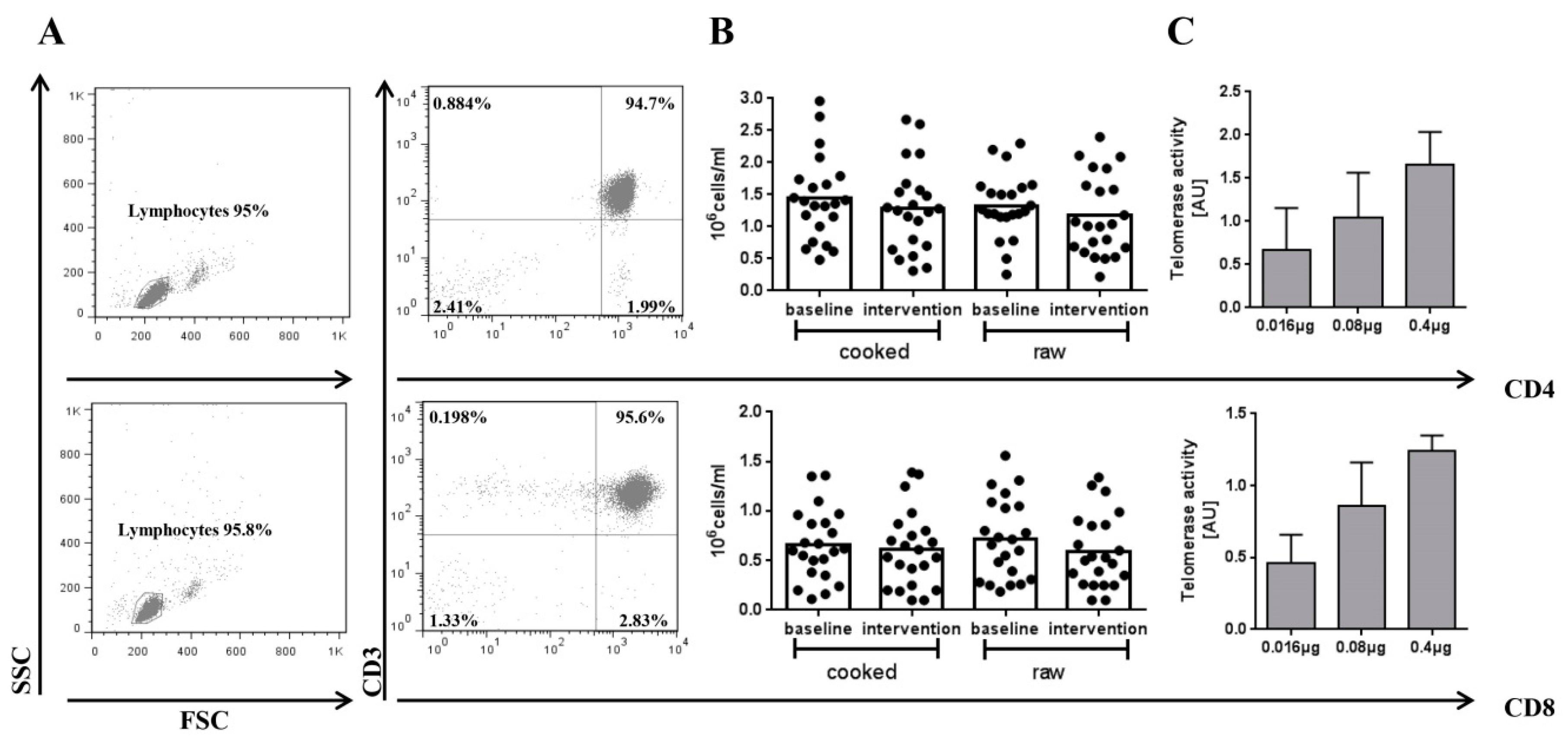
2.3. Blood Collection and Isolation of CD4+ or CD8+ T cells
2.4. Phytochemical Analysis in the Plant Material
2.5. Telomerase Activity Analysis by TRAP-ELISA Assay
2.6. Flow Cytometry
2.7. Statistical Analysis
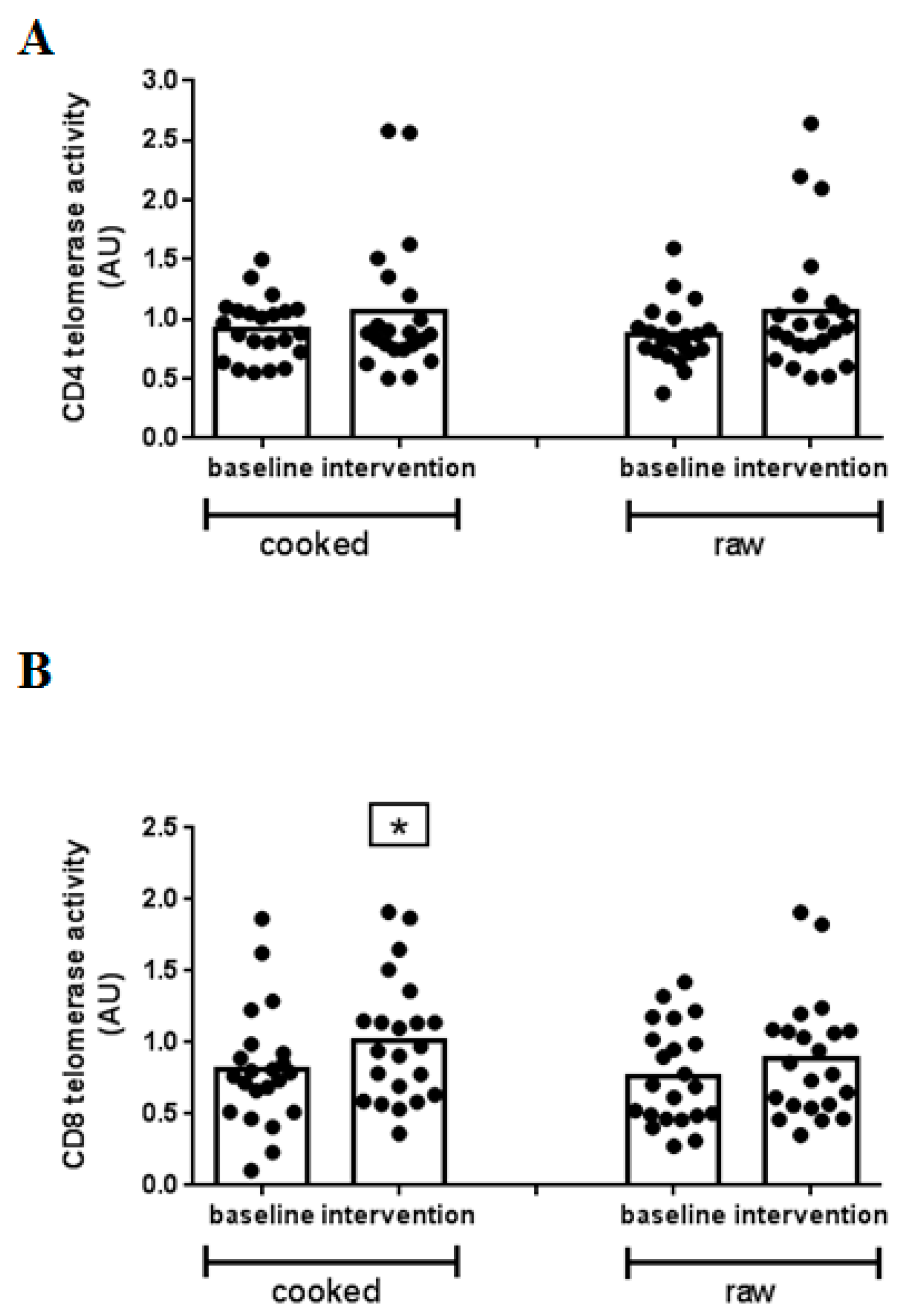
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lamy, E.; Goetz, V.; Erlacher, M.; Herz, C.; Mersch-Sundermann, V. Htert: Another brick in the wall of cancer cells. Mutat. Res. 2013, 752, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, H.; Schachter, F.; Uchida, I.; Wei, L.; Zhu, X.; Effros, R.; Cohen, D.; Harley, C.B. Loss of telomeric DNA during aging of normal and trisomy 21 human lymphocytes. Am. J. Hum. Genet. 1993, 52, 661–667. [Google Scholar] [PubMed]
- Effros, R.B. Telomere/telomerase dynamics within the human immune system: Effect of chronic infection and stress. Exp. Gerontol. 2011, 46, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Linton, P.J.; Dorshkind, K. Age-related changes in lymphocyte development and function. Nat. Immunol. 2004, 5, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Epel, E.S.; Lin, J.; Wilhelm, F.H.; Wolkowitz, O.M.; Cawthon, R.; Adler, N.E.; Dolbier, C.; Mendes, W.B.; Blackburn, E.H. Cell aging in relation to stress arousal and cardiovascular disease risk factors. Psychoneuroendocrinology 2006, 31, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Guo, D.; Li, K.; Pedersen-White, J.; Stallmann-Jorgensen, I.S.; Huang, Y.; Parikh, S.; Liu, K.; Dong, Y. Increased telomerase activity and vitamin d supplementation in overweight african americans. Int. J. Obes. 2012, 36, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Ornish, D.; Lin, J.; Chan, J.M.; Epel, E.; Kemp, C.; Weidner, G.; Marlin, R.; Frenda, S.J.; Magbanua, M.J.; Daubenmier, J.; et al. Effect of comprehensive lifestyle changes on telomerase activity and telomere length in men with biopsy-proven low-risk prostate cancer: 5-year follow-up of a descriptive pilot study. Lancet Oncol. 2013, 14, 1112–1120. [Google Scholar] [CrossRef]
- Zhang, X.; Shu, X.O.; Xiang, Y.B.; Yang, G.; Li, H.; Gao, J.; Cai, H.; Gao, Y.T.; Zheng, W. Cruciferous vegetable consumption is associated with a reduced risk of total and cardiovascular disease mortality. Am. J. Clin. Nutr. 2011, 94, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Blekkenhorst, L.C.; Bondonno, C.P.; Lewis, J.R.; Devine, A.; Zhu, K.; Lim, W.H.; Woodman, R.J.; Beilin, L.J.; Prince, R.L.; Hodgson, J.M. Cruciferous and allium vegetable intakes are inversely associated with 15-year atherosclerotic vascular disease deaths in older adult women. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Hosseinpanah, F.; Hedayati, M.; Hosseinpour-Niazi, S.; Azizi, F. Broccoli sprouts reduce oxidative stress in type 2 diabetes: A randomized double-blind clinical trial. Eur. J. Clin. Nutr. 2011, 65, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Tohidi, M.; Nazeri, P.; Mehran, M.; Azizi, F.; Mirmiran, P. Effect of broccoli sprouts on insulin resistance in type 2 diabetic patients: A randomized double-blind clinical trial. Int. J. Food Sci. Nutr. 2012, 63, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.H.; Kristal, A.R.; Stanford, J.L. Fruit and vegetable intakes and prostate cancer risk. J. Natl. Cancer Inst. 2000, 92, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E.; Rimm, E.B.; Liu, Y.; Stampfer, M.J.; Willett, W.C. A prospective study of cruciferous vegetables and prostate cancer. Cancer epidemiology, biomarkers & prevention: A publication of the American Association for Cancer Research. Am. Soc. Prev. Oncol. 2003, 12, 1403–1409. [Google Scholar]
- Ngwene, B.; Neugart, S.; Baldermann, S.; Ravi, B.; Schreiner, M. Intercropping induces changes in specific secondary metabolite concentration in ethiopian kale (brassica carinata) and african nightshade (solanum scabrum) under controlled conditions. Front. Plant Sci. 2017, 8, 1700. [Google Scholar] [CrossRef] [PubMed]
- Odongo, G.A.; Schlotz, N.; Herz, C.; Hanschen, F.S.; Baldermann, S.; Neugart, S.; Trierweiler, B.; Frommherz, L.; Franz, C.M.; Ngwene, B.; et al. The role of plant processing for the cancer preventive potential of ethiopian kale (brassica carinata). Food Nutr. Res. 2017, 61, 1271527. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Wagstaff, C. Enhancement of glucosinolate and isothiocyanate profiles in brassicaceae crops: Addressing challenges in breeding for cultivation, storage, and consumer-related traits. J. Agric. Food Chem. 2017, 65, 9379–9403. [Google Scholar] [CrossRef] [PubMed]
- Herz, C.; Márton, M.R.; Tran, H.T.T.; Gründemann, C.; Schell, J.; Lamy, E. Benzyl isothiocyanate but not benzyl nitrile from brassicales plants dually blocks the cox and lox pathway in primary human immune cells. J. Funct. Foods 2016, 23, 135–143. [Google Scholar] [CrossRef]
- Singh, S.V.; Singh, K. Cancer chemoprevention with dietary isothiocyanates mature for clinical translational research. Carcinogenesis 2012, 33, 1833–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giambanelli, E.; Verkerk, R.; D’Antuono, L.F.; Oliviero, T. The kinetic of key phytochemical compounds of non-heading and heading leafy brassica oleracea landraces as affected by traditional cooking methods. J. Sci. Food Agric. 2016, 96, 4772–4784. [Google Scholar] [CrossRef]
- Schlotz, N.; Odongo, G.A.; Herz, C.; Wassmer, H.; Kuhn, C.; Hanschen, F.S. Are raw brassica vegetables healthier than cooked ones? A randomized, controlled crossover intervention trial on the health-promoting potential of ethiopian kale. Nutrients 2018, 10, 1622. [Google Scholar] [CrossRef]
- Lamy, E.; Herz, C.; Lutz-Bonengel, S.; Hertrampf, A.; Marton, M.R.; Mersch-Sundermann, V. The mapk pathway signals telomerase modulation in response to isothiocyanate-induced DNA damage of human liver cancer cells. PLoS ONE 2013, 8, e53240. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Balcerczyk, A.; Gajewska, A.; Macierzynska-Piotrowska, E.; Pawelczyk, T.; Bartosz, G.; Szemraj, J. Enhanced antioxidant capacity and anti-ageing biomarkers after diet micronutrient supplementation. Molecules 2014, 19, 14794–14808. [Google Scholar] [CrossRef] [PubMed]
- Boccardi, V.; Esposito, A.; Rizzo, M.R.; Marfella, R.; Barbieri, M.; Paolisso, G. Mediterranean diet, telomere maintenance and health status among elderly. PLoS ONE 2013, 8, e62781. [Google Scholar] [CrossRef] [PubMed]
- Ornish, D.; Lin, J.; Daubenmier, J.; Weidner, G.; Epel, E.; Kemp, C.; Magbanua, M.J.M.; Marlin, R.; Yglecias, L.; Carroll, P.R.; et al. Increased telomerase activity and comprehensive lifestyle changes: A pilot study. Lancet Oncol. 2008, 9, 1048–1057. [Google Scholar] [CrossRef]
- Weng, N.P. Telomere and adaptive immunity. Mech. Ageing Dev. 2008, 129, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Mariani, E.; Meneghetti, A.; Formentini, I.; Neri, S.; Cattini, L.; Ravaglia, G.; Forti, P.; Facchini, A. Different rates of telomere shortening and telomerase activity reduction in cd8 t and cd16 nk lymphocytes with ageing. Exp. Gerontol. 2003, 38, 653–659. [Google Scholar] [CrossRef]
- Mariani, E.; Meneghetti, A.; Formentini, I.; Neri, S.; Cattini, L.; Ravaglia, G.; Forti, P.; Facchini, A. Telomere length and telomerase activity: Effect of ageing on human NK cells. Mech. Ageing Dev. 2003, 124, 403–408. [Google Scholar] [CrossRef]
- Lin, J.; Epel, E.; Cheon, J.; Kroenke, C.; Sinclair, E.; Bigos, M.; Wolkowitz, O.; Mellon, S.; Blackburn, E. Analyses and comparisons of telomerase activity and telomere length in human T and B cells: Insights for epidemiology of telomere maintenance. J. Immunol. Methods 2010, 352, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Reichert, T.; DeBruyere, M.; Deneys, V.; Totterman, T.; Lydyard, P.; Yuksel, F.; Chapel, H.; Jewell, D.; Van Hove, L.; Linden, J.; et al. Lymphocyte subset reference ranges in adult caucasians. Clin. Immunol. Immunopathol. 1991, 60, 190–208. [Google Scholar] [CrossRef]
- Leung, V.; Gillis, J.; Raboud, J.; Cooper, C.; Hogg, R.S.; Loutfy, M.R.; Machouf, N.; Montaner, J.S.G.; Rourke, S.B.; Tsoukas, C.; et al. Predictors of cd4:Cd8 ratio normalization and its effect on health outcomes in the era of combination antiretroviral therapy. PLoS ONE 2013, 8, e77665. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Fauce, S.R.; Effros, R.B. Reduced telomerase activity in human T lymphocytes exposed to cortisol. Brain Behav. Immun. 2008, 22, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Schutte, N.S.; Malouff, J.M. A meta-analytic review of the effects of mindfulness meditation on telomerase activity. Psychoneuroendocrinology 2014, 42, 45–48. [Google Scholar] [CrossRef] [PubMed]
- O’Bryan, J.M.; Woda, M.; Co, M.; Mathew, A.; Rothman, A.L. Telomere length dynamics in human memory t cells specific for viruses causing acute or latent infections. Immun. Ageing 2013, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Rufer, N.; Migliaccio, M.; Antonchuk, J.; Humphries, R.K.; Roosnek, E.; Lansdorp, P.M. Transfer of the human telomerase reverse transcriptase (TERT) gene into t lymphocytes results in extension of replicative potential. Blood 2001, 98, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Svenson, U.; Nordfjall, K.; Baird, D.; Roger, L.; Osterman, P.; Hellenius, M.L.; Roos, G. Blood cell telomere length is a dynamic feature. PLoS ONE 2011, 6, e21485. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G.; Lansdorp, P.M. Telomeres and aging. Physiol. Rev. 2008, 88, 557–579. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.D.; Wen, M.S.; Shie, S.S.; Lo, Y.L.; Wo, H.T.; Wang, C.C.; Hsieh, I.C.; Lee, T.H.; Wang, C.Y. The circadian rhythm controls telomeres and telomerase activity. Biochem. Biophys. Res. Commun. 2014, 451, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Odongo, G.A.; Schlotz, N.; Baldermann, S.; Neugart, S.; Ngwene, B.; Schreiner, M.; Lamy, E. Effects of amaranthus cruentus l. On aflatoxin b1- and oxidative stress-induced DNA damage in human liver (hepg2) cells. Food Biosci. 2018, 26, 42–48. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Typek, R. Thermal stability of 5-o-caffeoylquinic acid in aqueous solutions at different heating conditions. J. Agric. Food Chem. 2010, 58, 12578–12584. [Google Scholar] [CrossRef]
- Herz, C.; Hertrampf, A.; Zimmermann, S.; Stetter, N.; Wagner, M.; Kleinhans, C.; Erlacher, M.; Schuler, J.; Platz, S.; Rohn, S.; et al. The isothiocyanate erucin abrogates telomerase in hepatocellular carcinoma cells in vitro and in an orthotopic xenograft tumour model of Hcc. J. Cell. Mol. Med. 2014, 18, 2393–2403. [Google Scholar] [CrossRef]
- Sadava, D.; Whitlock, E.; Kane, S.E. The green tea polyphenol, epigallocatechin-3-gallate inhibits telomerase and induces apoptosis in drug-resistant lung cancer cells. Biochem. Biophys. Res. Commun. 2007, 360, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Eitsuka, T.; Nakagawa, K.; Kato, S.; Ito, J.; Otoki, Y.; Takasu, S.; Shimizu, N.; Takahashi, T.; Miyazawa, T. Modulation of telomerase activity in cancer cells by dietary compounds: A review. Int. J. Mol. Sci. 2018, 19, 478. [Google Scholar] [CrossRef] [PubMed]
- Odongo, G.A.; Skatchkov, I.; Herz, C.; Lamy, E. Optimization of the alkaline comet assay for easy repair capacity quantification of oxidative DNA damage in pbmc from human volunteers using aphidicolin block. DNA Repair 2019, 77, 58–64. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, H.T.T.; Schlotz, N.; Schreiner, M.; Lamy, E. Short-Term Dietary Intervention with Cooked but Not Raw Brassica Leafy Vegetables Increases Telomerase Activity in CD8+ Lymphocytes in a Randomized Human Trial. Nutrients 2019, 11, 786. https://doi.org/10.3390/nu11040786
Tran HTT, Schlotz N, Schreiner M, Lamy E. Short-Term Dietary Intervention with Cooked but Not Raw Brassica Leafy Vegetables Increases Telomerase Activity in CD8+ Lymphocytes in a Randomized Human Trial. Nutrients. 2019; 11(4):786. https://doi.org/10.3390/nu11040786
Chicago/Turabian StyleTran, Hoai Thi Thu, Nina Schlotz, Monika Schreiner, and Evelyn Lamy. 2019. "Short-Term Dietary Intervention with Cooked but Not Raw Brassica Leafy Vegetables Increases Telomerase Activity in CD8+ Lymphocytes in a Randomized Human Trial" Nutrients 11, no. 4: 786. https://doi.org/10.3390/nu11040786
APA StyleTran, H. T. T., Schlotz, N., Schreiner, M., & Lamy, E. (2019). Short-Term Dietary Intervention with Cooked but Not Raw Brassica Leafy Vegetables Increases Telomerase Activity in CD8+ Lymphocytes in a Randomized Human Trial. Nutrients, 11(4), 786. https://doi.org/10.3390/nu11040786