Methyljasmonate Elicitation Increases Terpenoid Indole Alkaloid Accumulation in Rhazya stricta Hairy Root Cultures


 , ,
, ,  and
and 
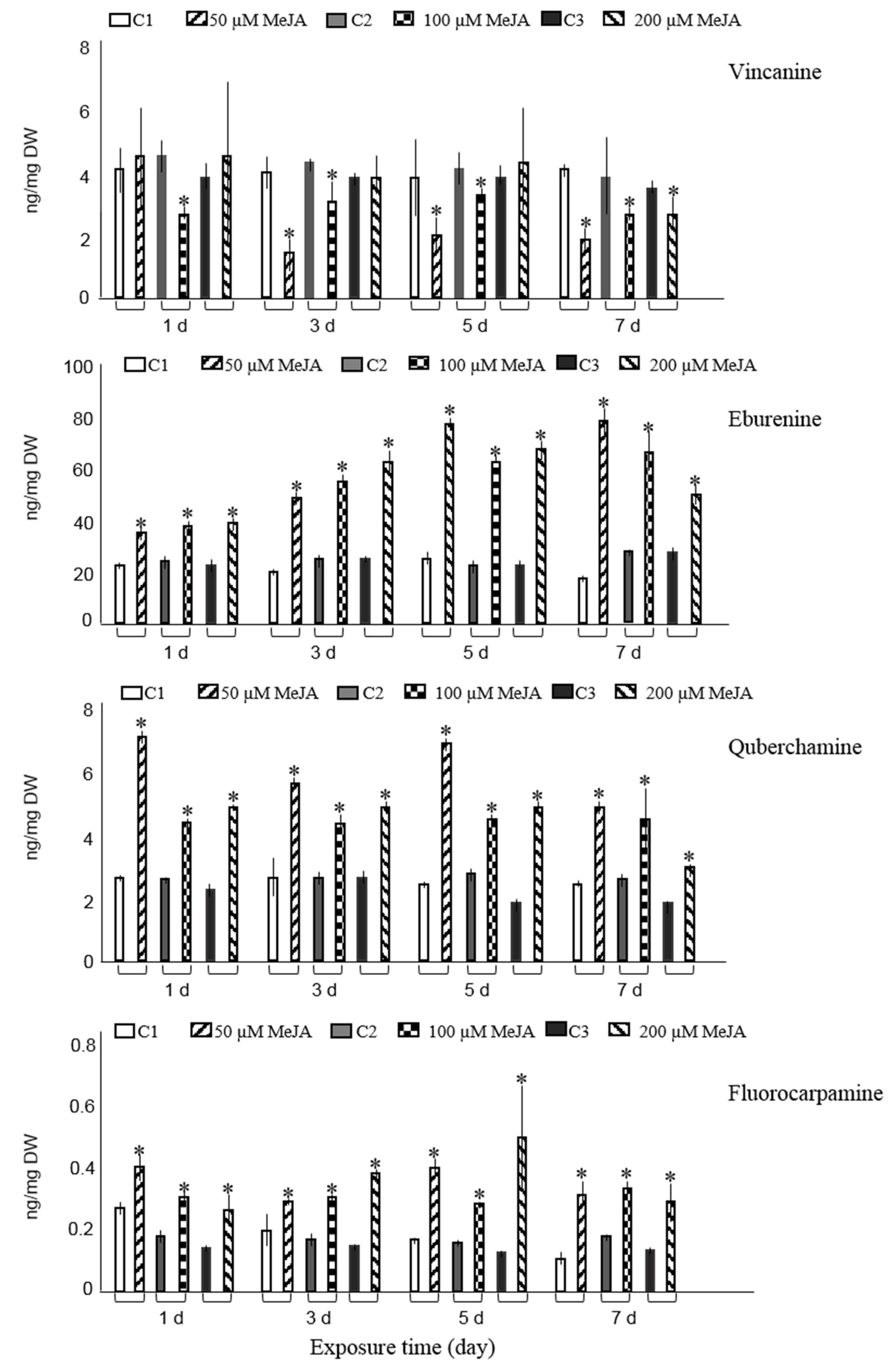{kind=link}
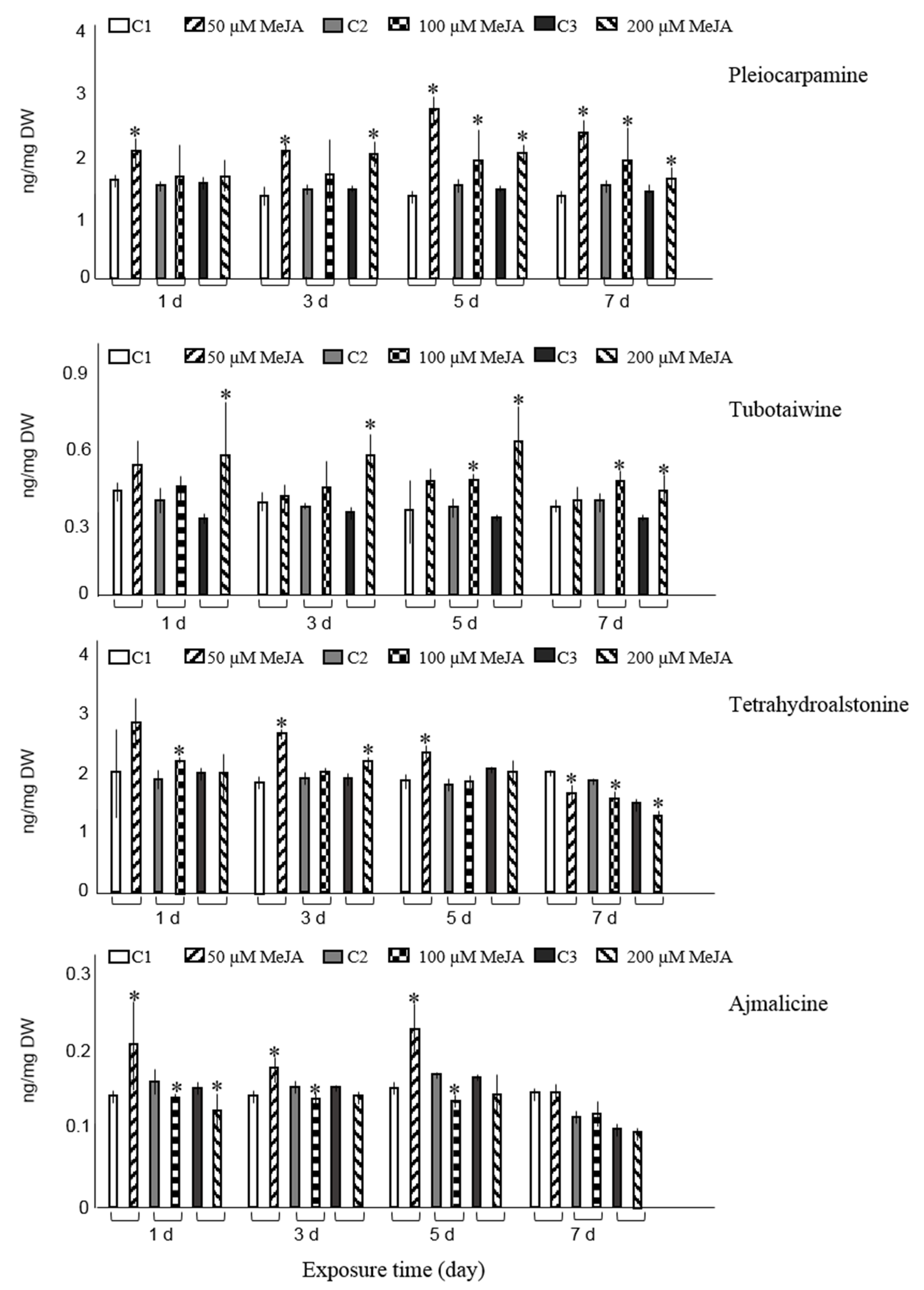{kind=link}
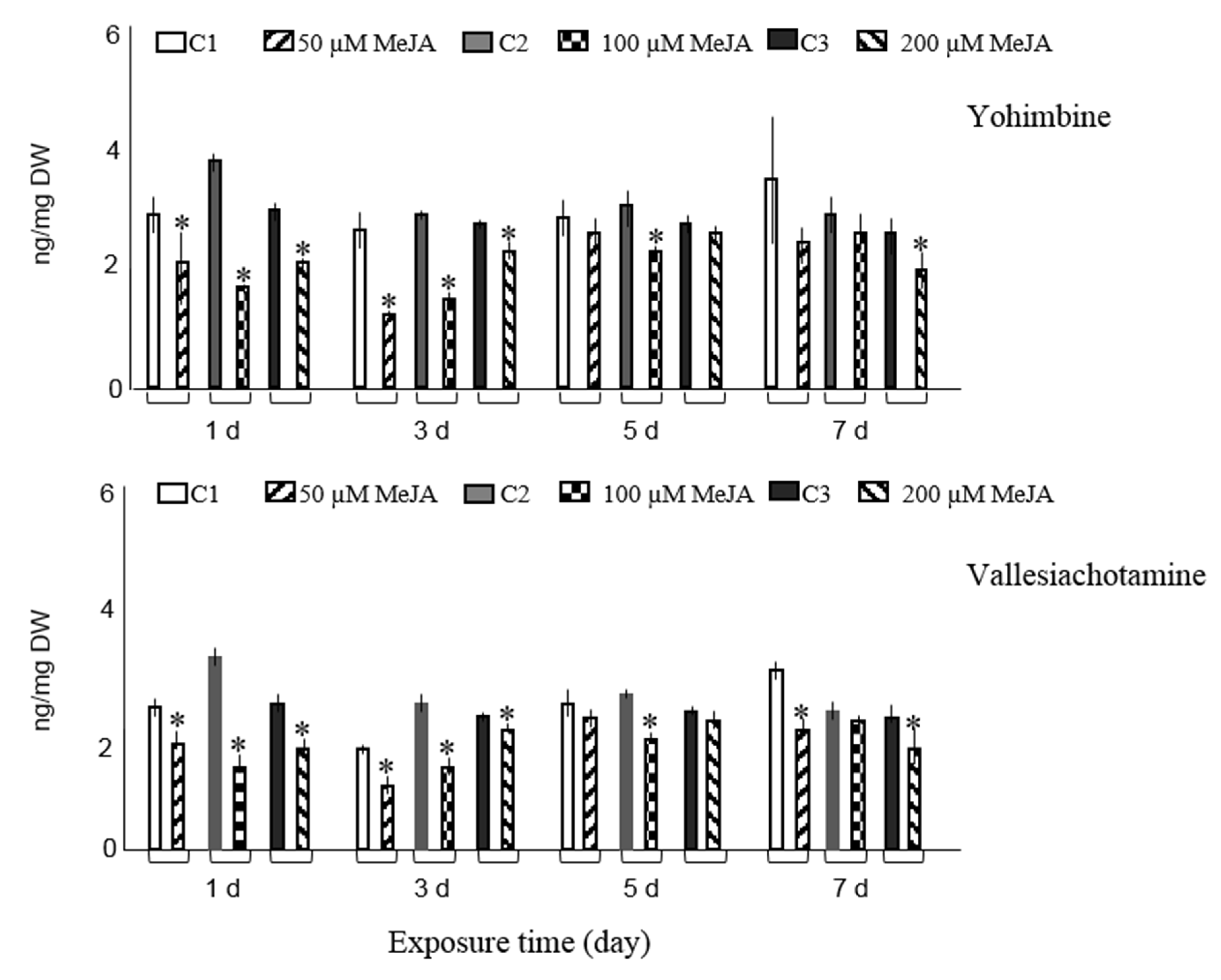{kind=link}
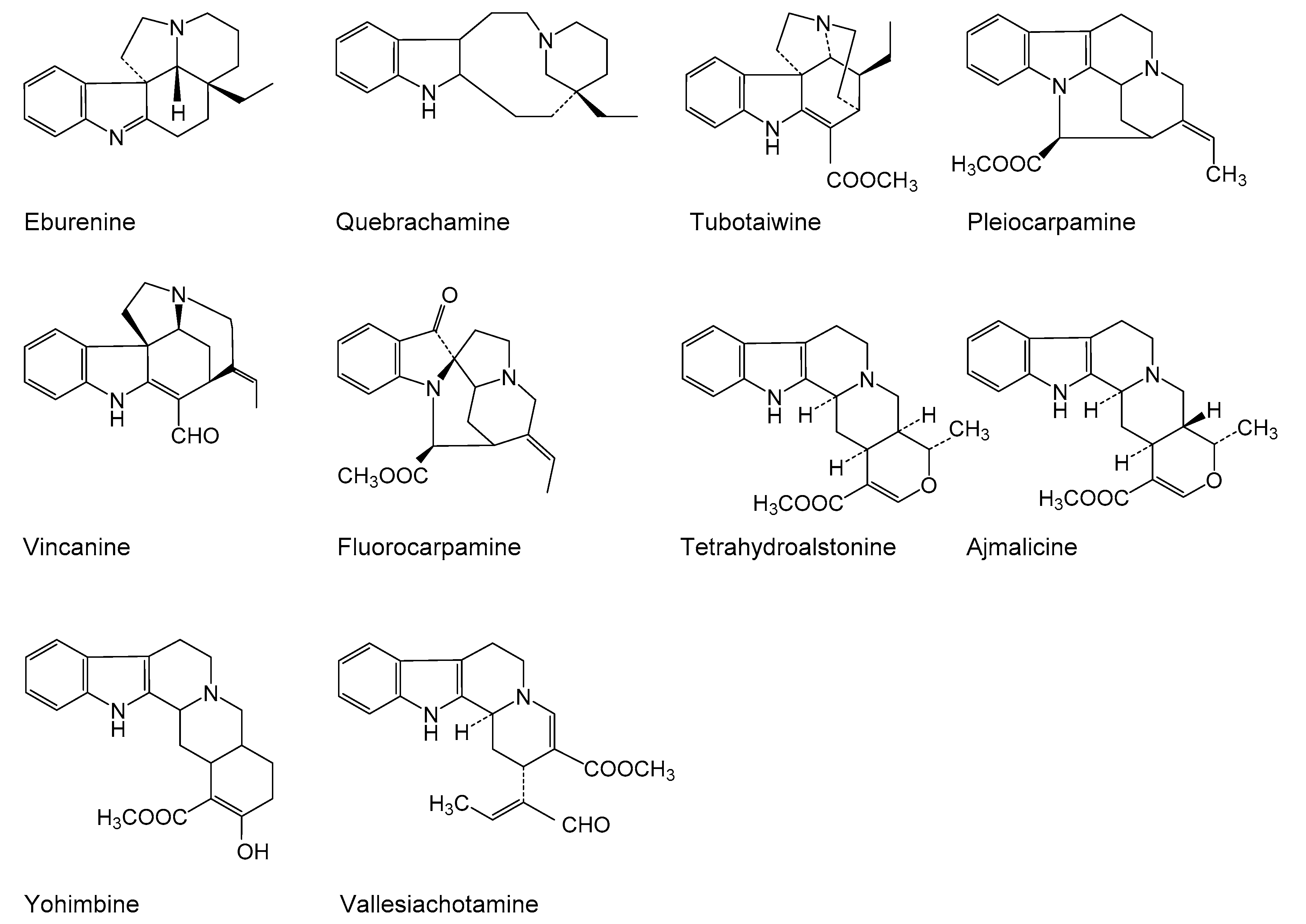{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hairy Root Induction
2.2. Elicitation
2.3. Extraction of Alkaloids and GC-MS Analysis
2.4. Extraction of Compounds and Metabolic Profiling by 1H NMR
2.5. Statistical Analysis
3. Results and Discussion
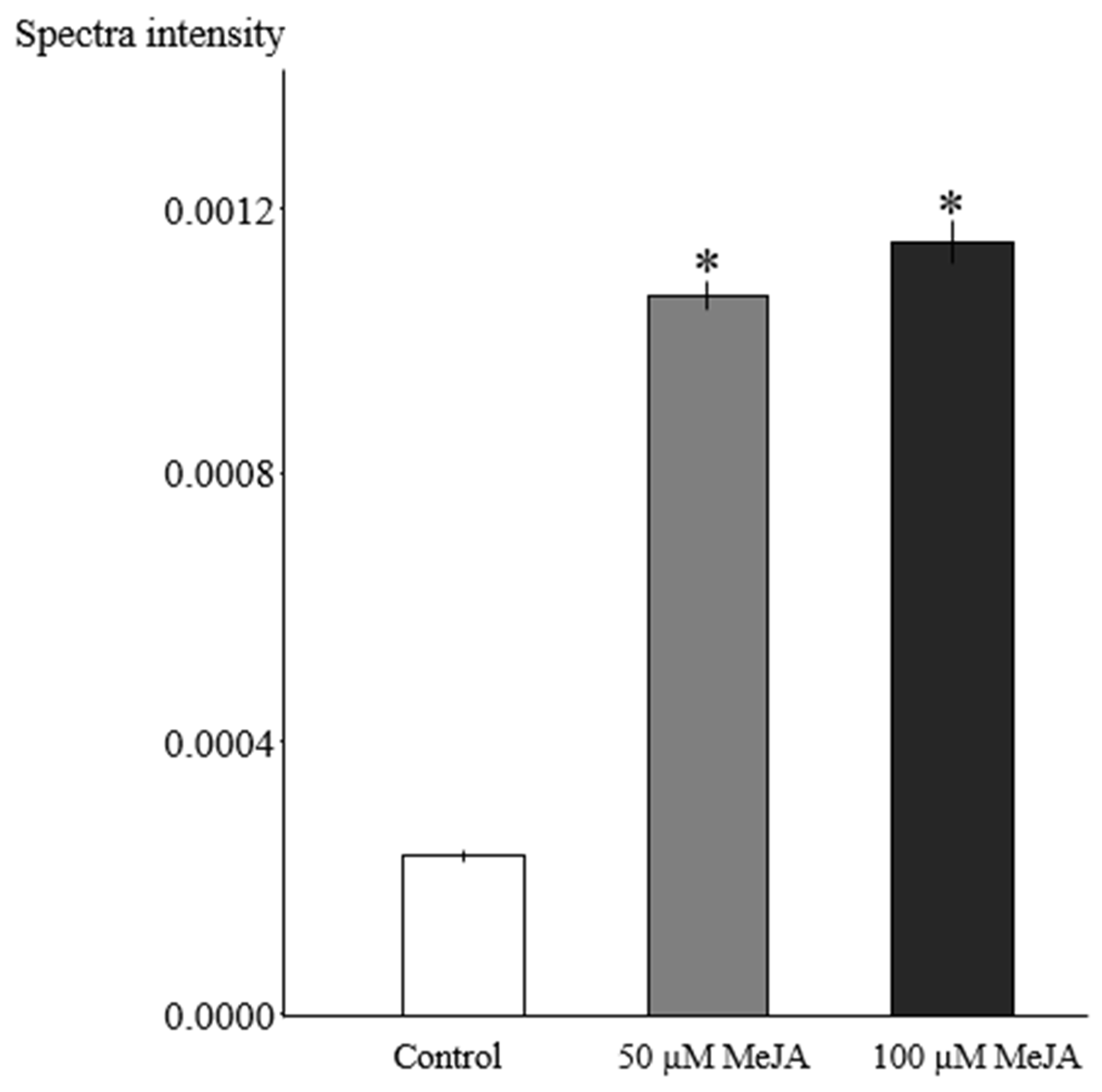
3.1. Effect of Elicitation on Alkaloid Accumulation
3.2. Metabolite Profile Changes after Elicitation Monitored by NMR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilani, S.A.; Kikuchi, A.; Shinwari, Z.K.; Khattak, Z.I.; Watanabe, K.N. Phytochemical, pharmacological and ethnobotanical studies of Rhazya stricta Decne. Phyther. Res. 2007, 21, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.U.; Qureshi, M.M.; Zaman, K.; Malik, S.; Ali, S.S. The alkaloids of Rhazya stricta and R. orientalis. A review. Fitoterapia 1989, 60, 291–322. [Google Scholar]
- Elkady, A.I. Crude alkaloid extract of Rhazya stricta inhibits cell growth and sensitizes human lung cancer cells to cisplatin through induction of apoptosis. Genet. Mol. Biol. 2013, 36, 12–21. [Google Scholar] [CrossRef] [PubMed]
- David, B.; Sévenet, T.; Morgat, M.; Guénard, D.; Moisand, A.; Tollon, Y.; Thoison, O.; Wright, M. Rhazinilam mimics the cellular effects of taxol by different mechanisms of action. Cell Motil. Cytoskelet. 1994, 28, 317–326. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Handy, G.A.; Funayama, S.; Cordell, G.A. Anticancer indole alkaloids of Rhazya stricta. J. Nat. Prod. 1981, 44, 696–700. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Weber, J.; MacIuk, A. Bioprocessing of plant cell cultures for mass production of targeted compounds. Appl. Microbiol. Biotechnol. 2009, 83, 809–823. [Google Scholar] [CrossRef]
- Pauwels, L.; Inzé, D.; Goossens, A. Jasmonate-inducible gene: What does it mean? Trends Plant Sci. 2009, 14, 87–91. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef]
- Wilson, S.A.; Roberts, S.C. Recent advances towards development and commercialization of plant cell culture processes for the synthesis of biomolecules. Plant Biotechnol. J. 2012, 10, 249–268. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Mishra, A.K.; Sharma, K.; Misra, R.S. Elicitor recognition, signal transduction and induced resistance in plants. J. Plant Interact. 2012, 7, 95–120. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S. Natural plant genetic engineer Agrobacterium rhizogenes: Role of T-DNA in plant secondary metabolism. Biotechnol. Lett. 2012, 34, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A tool for enriching the bioactive composition of foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef] [PubMed]
- Memelink, J.; Kijne, J.W.; van der Heijden, R.; Verpoorte, R. Genetic modification of plant secondary metabolite pathways using transcriptional regulators. Adv. Biochem. Eng. Biotechnol. 2001, 72, 103–125. [Google Scholar] [PubMed]
- Rischer, H.; Orešič, M.; Seppänen-Laakso, T.; Katajamaa, M.; Lammertyn, F.; Ardiles-Diaz, W.; Van Montagu, M.C.E.; Inze, D.; Oksman-Caldentey, K.M.; Goossens, A. Gene-to-metabolite networks for terpenoid indole alkaloid biosynthesis in Catharanthus roseus cells. Proc. Natl. Acad. Sci. USA 2006, 103, 5614–5619. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, K.; Dong, L.; Navrot, N.; Schneider, T.; Burlat, V.; Pollier, J.; Woittiez, L.; Van Der Krol, S.; Lugan, R.; Ilc, T.; et al. The seco-iridoid pathway from Catharanthus roseus. Nat. Commun. 2014, 5, 3606. [Google Scholar] [CrossRef]
- Zhou, M.L.; Zhu, X.M.; Shao, J.R.; Tang, Y.X.; Wu, Y.M. Production and metabolic engineering of bioactive substances in plant hairy root culture. Appl. Microbiol. Biotechnol. 2011, 90, 1229–1239. [Google Scholar] [CrossRef]
- Cusidó, R.M.; Onrubia, M.; Sabater-Jara, A.B.; Moyano, E.; Bonfill, M.; Goossens, A.; Angeles Pedreño, M.; Palazon, J. A rational approach to improving the biotechnological production of taxanes in plant cell cultures of Taxus spp. Biotechnol. Adv. 2014, 32, 1157–1167. [Google Scholar] [CrossRef]
- Pawelka, K.H.; Stöckigt, J. Major indole alkaloids produced in cell suspension cultures of Rhazya stricta Decaisne. Z. Naturforsch.–Sect. C J. Biosci. 1986, 41, 385–390. [Google Scholar] [CrossRef]
- Sheludko, Y.; Gerasimenko, I.; Unger, M.; Kostenyuk, I.; Stöckigt, J. Induction of alkaloid diversity in hybrid plant cell cultures. Plant Cell Rep. 1999, 18, 911–918. [Google Scholar] [CrossRef]
- Sheludko, Y.; Gerasimenko, I.; Platonova, O. Divergence of the indole alkaloid pattern in two somatic hybrid plant cell subcultures of Rauvolfia serpentina × Rhazya stricta. Planta Med. 2000, 66, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Akhgari, A.; Yrjönen, T.; Laakso, I.; Vuorela, H.; Oksman-Caldentey, K.M.; Rischer, H. Establishment of transgenic Rhazya stricta hairy roots to modulate terpenoid indole alkaloid production. Plant. Cell Rep. 2015, 34, 1939–1952. [Google Scholar] [CrossRef] [PubMed]
- Akhgari, A.; Laakso, I.; Seppänen-Laakso, T.; Yrjönen, T.; Vuorela, H.; Oksman-Caldentey, K.M.; Rischer, H. Determination of terpenoid indole alkaloids in hairy roots of Rhazya stricta (Apocynaceae) by GC-MS. Phytochem. Anal. 2015, 26, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Akhgari, A.; Laakso, I.; Seppänen-Laakso, T.; Yrjönen, T.; Vuorela, H.; Oksman-Caldentey, K.M.; Rischer, H. Analysis of indole alkaloids from Rhazya stricta hairy roots by ultra-performance liquid chromatography-mass spectrometry. Molecules 2015, 20, 22621–22634. [Google Scholar] [CrossRef]
- Oksman-Caldentey, K.M.; Kivelä, O.; Hiltunen, R. Spontaneous shoot organogenesis and plant regeneration from hairy root cultures of Hyoscyamus muticus. Plant Sci. 1991, 78, 129–136. [Google Scholar] [CrossRef]
- O’Connor, S.E.; Maresh, J.J. Chemistry and biology of monoterpene indole alkaloid biosynthesis. Nat. Prod. Rep. 2006, 23, 532–547. [Google Scholar] [CrossRef]
- Stöckigt, J. Indirect involvement of geissoschizine in the biosynthesis of ajmalicine and related alkaloids. J. Chem. Soc. Chem. Commun. 1978, 1097–1099. [Google Scholar] [CrossRef]
- Stavrinides, A.; Tatsis, E.C.; Foureau, E.; Caputi, L.; Kellner, F.; Courdavault, V.; O’Connor, S.E. Unlocking the diversity of alkaloids in Catharanthus roseus: Nuclear localization suggests metabolic channeling in secondary metabolism. Chem. Biol. 2015, 22, 336–341. [Google Scholar] [CrossRef]
- Stavrinides, A.; Tatsis, E.C.; Caputi, L.; Foureau, E.; Stevenson, C.E.M.; Lawson, D.M.; Courdavault, V.; O’Connor, S.E. Structural investigation of heteroyohimbine alkaloid synthesis reveals active site elements that control stereoselectivity. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Buckingham, J.; Baggaley, K.H.; Roberts, A.D.; Szabó, L.F. Dictionary of Alkaloids, 2nd ed.; CRC Press/Taylor & Francis Group: London, UK, 2010. [Google Scholar]
- Saxton, J.E. Recent progress in the chemistry of the monoterpenoid lndole alkaloids. Nat. Prod. Rep. 1995, 4, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Szabó, L.F. Rigorous biogenetic network for a group of indole alkaloids derived from strictosidine. Molecules 2008, 13, 1875–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Eisenreich, W.; Kutchan, T.M. Bacterial biotransformation of 3α(S)-strictosidine to the monoterpenoid indole alkaloid vallesiachotamine. Phytochemistry 1998, 48, 293–296. [Google Scholar] [CrossRef]
- Kan-Fan, C.; Husson, H.P. Biomimetic synthesis of yohimbine and heteroyohimbine alkaloids from 4,21-dehydrogeissoschizine. Tetrahedron Lett. 1980, 21, 1463–1466. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed.; Wiley: New York, NY, USA, 2002; pp. 350–358. [Google Scholar]
- Gerasimenko, I.; Sheludko, Y.; Unger, M.; Stöckigt, J. Development of an efficient system for the separation of indole alkaloids by high performance liquid chromatography and its applications. Phytochem. Anal. 2001, 12, 96–103. [Google Scholar] [CrossRef]
- Kai, G.; Yang, S.; Zhang, Y.; Luo, X.; Fu, X.; Zhang, A.; Xiao, J. Effects of different elicitors on yield of tropane alkaloids in hairy roots of Anisodus acutangulus. Mol. Biol. Rep. 2012, 39, 1721–1729. [Google Scholar] [CrossRef]
- Dehghan, E.; Häkkinen, S.T.; Oksman-Caldentey, K.M.; Ahmadi, F.S. Production of tropane alkaloids in diploid and tetraploid plants and in vitro hairy root cultures of Egyptian henbane (Hyoscyamus muticus L.). Plant Cell. Tissue Organ. Cult. 2012, 110, 35–44. [Google Scholar] [CrossRef]
- Häkkinen, S.T.; Moyano, E.; Cusidó, R.M.; Oksman-Caldentey, K.M. Exploring the metabolic stability of engineered hairy roots after 16 years maintenance. Front. Plant Sci. 2016, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mano, Y.; Nabeshima, S.; Matsui, C.; Ohkawa, H. production of tropane alkaloids by hairy root cultures of Scopolia japonica. Agric. Biol. Chem. 1986, 50, 2715–2722. [Google Scholar] [CrossRef]
- Guivarc’h, A.; Boccara, M.; Prouteau, M.; Chriqui, D. Instability of phenotype and gene expression in long-term culture of carrot hairy root clones. Plant Cell Rep. 1999, 19, 43–50. [Google Scholar] [CrossRef]
- Goklany, S.; Loring, R.H.; Glick, J.; Lee-Parsons, C.W.T. Assessing the limitations to terpenoid indole alkaloid biosynthesis in Catharanthus roseus hairy root cultures through gene expression profiling and precursor feeding. Biotechnol. Prog. 2009, 25, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Lee-Parsons, C.W.T.; Ertürk, S.; Tengtrakool, J. Enhancement of ajmalicine production in Catharanthus roseus cell cultures with methyl jasmonate is dependent on timing and dosage of elicitation. Biotechnol. Lett. 2004, 26, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-May, E.; Galaz-Ávalos, R.M.; Loyola-Vargas, V.M. Differential secretion and accumulation of terpene indole alkaloids in hairy roots of Catharanthus roseus treated with methyl jasmonate. Mol. Biotechnol. 2009, 41, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Goldhaber-Pasillas, G.D.; Mustafa, N.R.; Verpoorte, R. Jasmonic acid effect on the fatty acid and terpenoid indole alkaloid accumulation in cell suspension cultures of Catharanthus roseus. Molecules 2014, 19, 10242–10260. [Google Scholar] [CrossRef] [Green Version]
- Saiman, M.Z.; Mustafa, N.R.; Choi, Y.H.; Verpoorte, R.; Schulte, A.E. Metabolic alterations and distribution of five-carbon precursors in jasmonic acid-elicited Catharanthus roseus cell suspension cultures. Plant Cell. Tissue Organ. Cult. 2015, 122, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, N.R.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Metabolic changes of salicylic acid-elicited Catharanthus roseus cell suspension cultures monitored by NMR-based metabolomics. Biotechnol. Lett. 2009, 31, 1967–1974. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.S.; Choi, Y.H.; Kim, H.K.; Linthorst, H.J.M.; Verpoorte, R. Metabolomic analysis of methyl jasmonate treated Brassica rapa leaves by 2-dimensional NMR spectroscopy. Phytochemistry 2006, 67, 2503–2511. [Google Scholar] [CrossRef]
- Zulak, K.G.; Weljie, A.M.; Vogel, H.J.; Facchini, P.J. Quantitative 1H NMR metabolomics reveals extensive metabolic reprogramming of primary and secondary metabolism in elicitor-treated opium poppy cell cultures. BMC Plant Biol. 2008, 8, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Flores-Sanchez, I.J.; Peč, J.; Fei, J.; Choi, Y.H.; Dušek, J.; Verpoorte, R. Elicitation studies in cell suspension cultures of Cannabis sativa L. J. Biotechnol. 2009, 143, 157–168. [Google Scholar] [CrossRef]
- Saiman, M.Z.; Mustafa, N.R.; Pomahočová, B.; Verberne, M.; Verpoorte, R.; Choi, Y.H.; Schulte, A.E. Analysis of metabolites in the terpenoid pathway of Catharanthus roseus cell suspensions. Plant Cell Tissue Organ. Cult. 2014, 117, 225–239. [Google Scholar] [CrossRef]
- Akhgari, A. Alkaloids of In Vitro Cultures of Rhazya Stricta. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2015. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhgari, A.; Laakso, I.; Maaheimo, H.; Choi, Y.H.; Seppänen-Laakso, T.; Oksman-Caldentey, K.-M.; Rischer, H. Methyljasmonate Elicitation Increases Terpenoid Indole Alkaloid Accumulation in Rhazya stricta Hairy Root Cultures. Plants 2019, 8, 534. https://doi.org/10.3390/plants8120534
Akhgari A, Laakso I, Maaheimo H, Choi YH, Seppänen-Laakso T, Oksman-Caldentey K-M, Rischer H. Methyljasmonate Elicitation Increases Terpenoid Indole Alkaloid Accumulation in Rhazya stricta Hairy Root Cultures. Plants. 2019; 8(12):534. https://doi.org/10.3390/plants8120534
Chicago/Turabian StyleAkhgari, Amir, Into Laakso, Hannu Maaheimo, Young Hae Choi, Tuulikki Seppänen-Laakso, Kirsi-Marja Oksman-Caldentey, and Heiko Rischer. 2019. "Methyljasmonate Elicitation Increases Terpenoid Indole Alkaloid Accumulation in Rhazya stricta Hairy Root Cultures" Plants 8, no. 12: 534. https://doi.org/10.3390/plants8120534
APA StyleAkhgari, A., Laakso, I., Maaheimo, H., Choi, Y. H., Seppänen-Laakso, T., Oksman-Caldentey, K.-M., & Rischer, H. (2019). Methyljasmonate Elicitation Increases Terpenoid Indole Alkaloid Accumulation in Rhazya stricta Hairy Root Cultures. Plants, 8(12), 534. https://doi.org/10.3390/plants8120534