GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration





Abstract
:1. Introduction
2. Structure of Human Brain Gangliosides
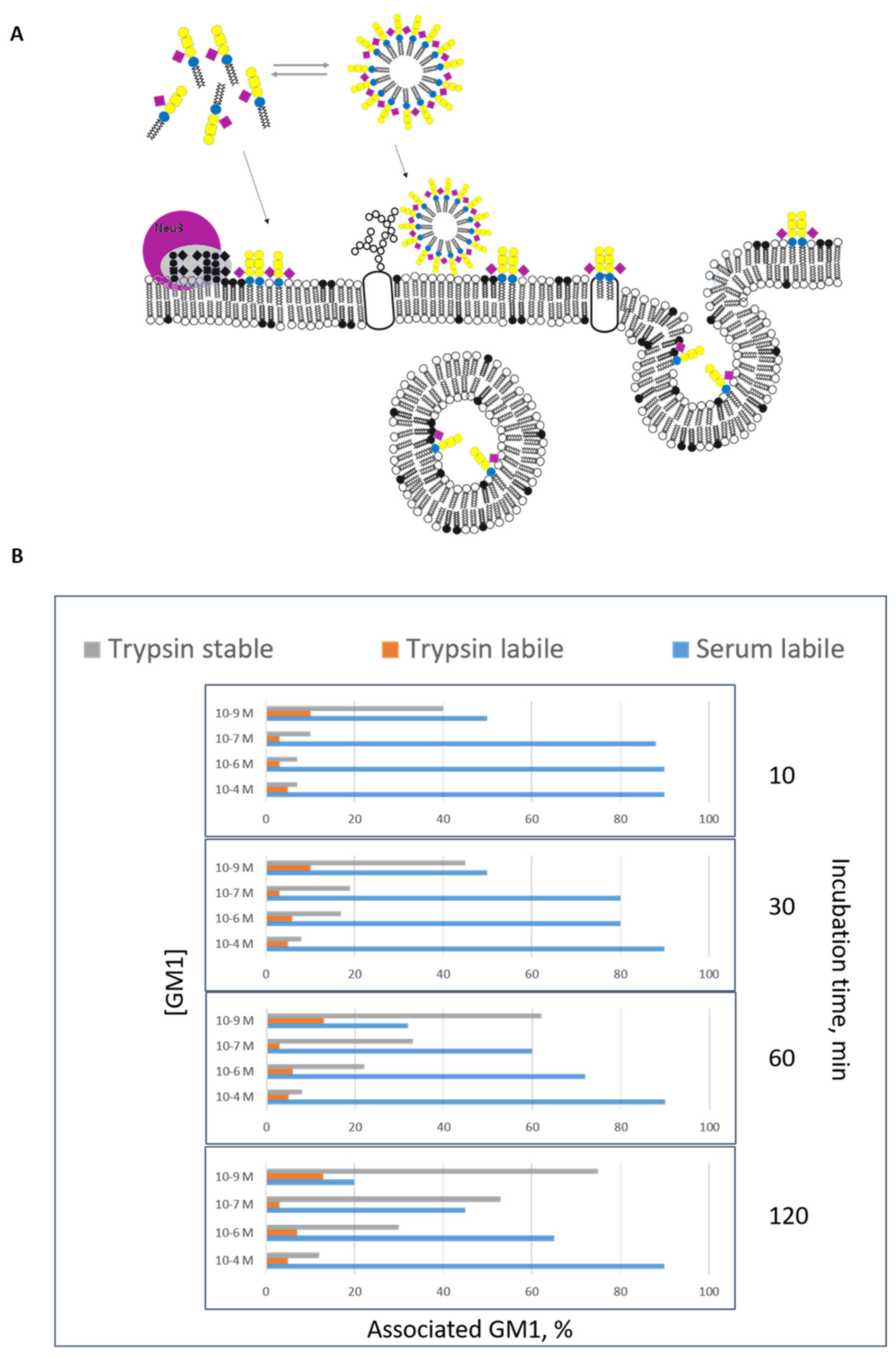
3. Gangliosides as the Main Actors in Membrane Organization: The Amphiphilic Properties of Gangliosides
4. GM1 of Neuronal Plasma Membrane in the Central Nervous System: Changes of Its Content During Aging and in Neurodegenerative Diseases
5. GM1 Preparations and GM1 Analysis
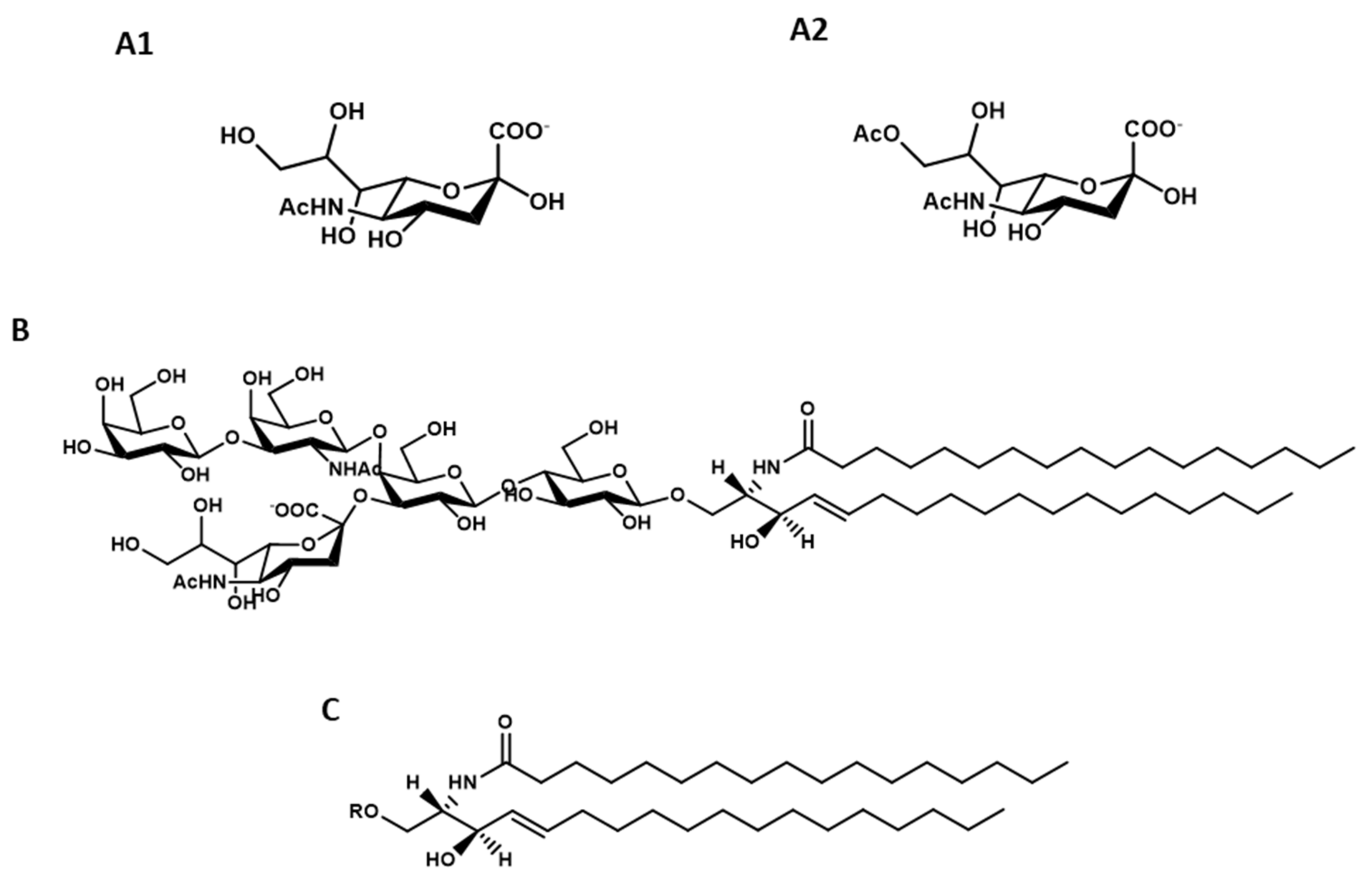
5.1. Preparation of GM1 with Heterogeneous Ceramide Moiety
5.2. Preparation of GM1 with Homogenous Ceramide Moiety
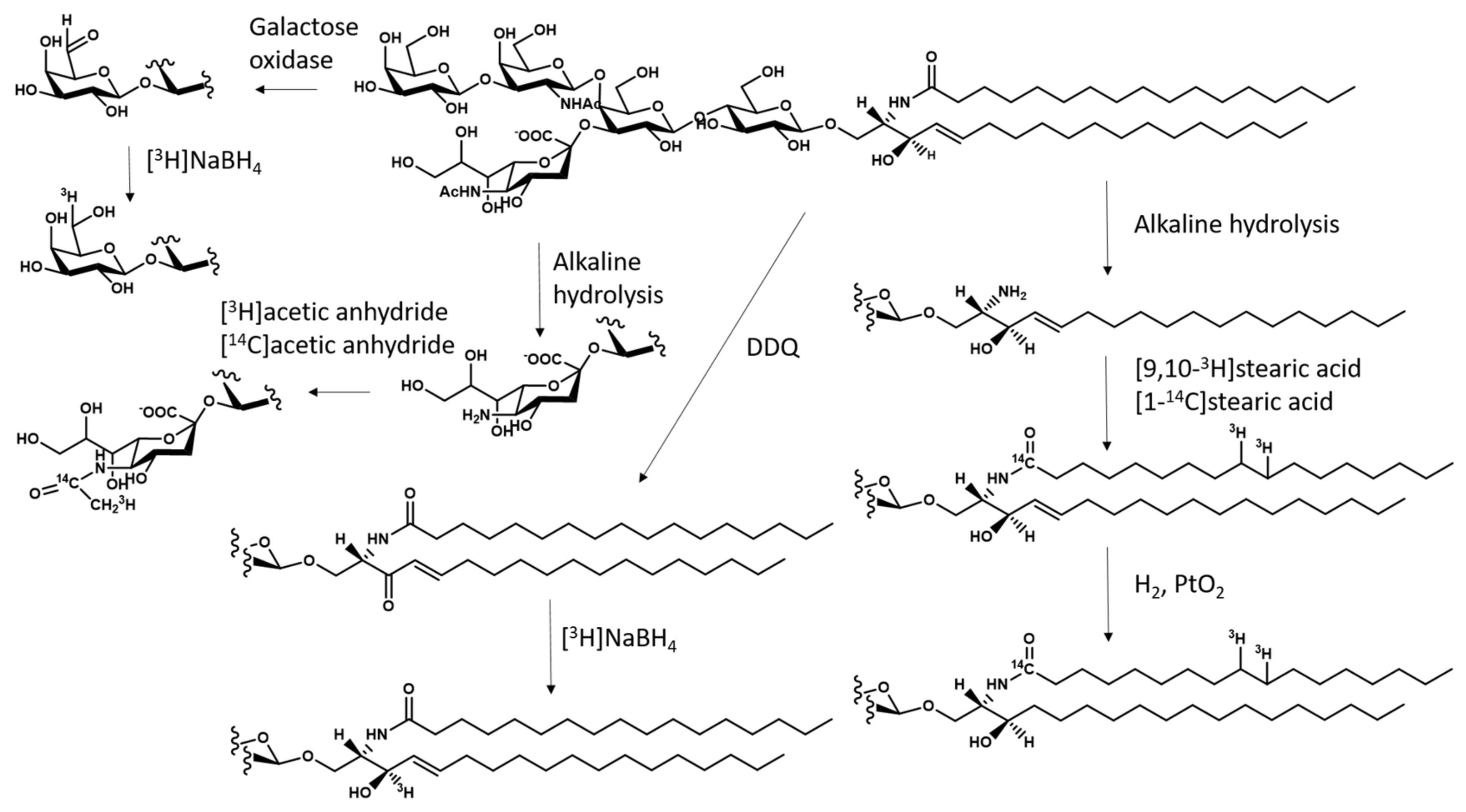
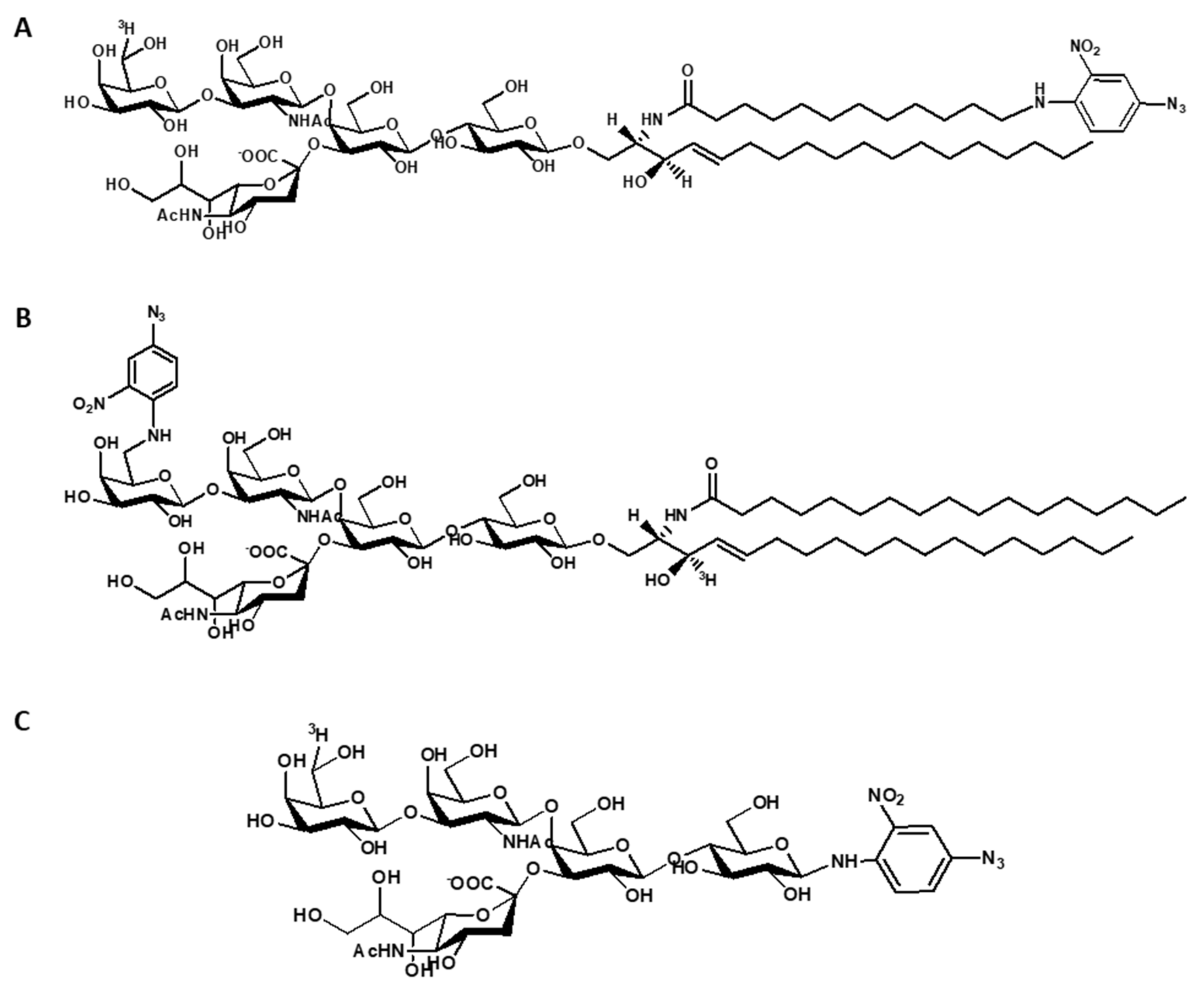
5.3. Preparation of Radioactive GM1 and its Derivatives
6. GM1 and Cholera Toxin
7. The Fate of GM1 Administered to Cells in Culture
8. Neurotrophic and Neuroprotective Properties of GM1
9. The Fate of GM1 Administered In Vivo within the Brain
10. GM1 as a Therapeutic Drug
10.1. GM1 and Parkinson’s Disease
10.2. GM1 and Alzheimer’s Disease
10.3. GM1 and Spinal Cord Injury
10.4. GM1 and Peripheral Neuroprotection by Oxaliplatin Treatment
10.5. GM1 in Brain Damage of Premature Infants
10.6. GM1 in Ischemic Damage of Adults
11. Administration of GM1 and Modified GM1 to B4galnt1−/+ Mice
12. The Oligosaccharide Chain is the Functional Portion of GM1
12.1. Administration of OligoGM1 to Cells in Vitro
12.2. OligoGM1 and TrkA Interaction Study in Silico
12.3. Administration of OligoGM1 to B4galnt1−/+ Mice
Funding
Conflicts of Interest
Abbreviations
| Ganglioside nomenclature is in accordance with IUPAC-IUB recommendations (IUPAC-IUMBMB 1998) | |
| BBB | blood brain barrier |
| c.m.c. | critical micellar concentration |
| GBS | Guillain-Barre syndrome |
| GM1 | II3Neu5Ac-Gg4Cer, β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-3)]-β-Gal-(1-4)-β-Glc-Cer; |
| N2a | Neuro2a cells |
| NGF | nerve growth factor |
| OligoGM1 | GM1 oligosaccharide, II3Neu5Ac-Gg4, β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-3)]-β-Gal-(1-4)-Glc |
| Trk | neurotrophin tyrosine kinase receptor |
References
- Kuhn, R.; Wiegandt, H. The Constitution of Gangliosides G-Ii, G-Iii and G-Iv. Z. Naturforsch B 1963, 18, 541–543. [Google Scholar] [CrossRef] [Green Version]
- Aureli, M.; Mauri, L.; Ciampa, M.G.; Prinetti, A.; Toffano, G.; Secchieri, C.; Sonnino, S. GM1 Ganglioside: Past Studies and Future Potential. Mol. Neurobiol. 2016, 53, 1824–1842. [Google Scholar] [CrossRef]
- Svennerholm, L. The Gangliosides. J. Lipid Res. 1964, 5, 145–155. [Google Scholar]
- Wiegandt, H. The structure and the function of gangliosides. Angew Chem. Int. Ed Engl. 1968, 7, 87–96. [Google Scholar] [CrossRef]
- Holmgren, J.; Lonnroth, I.; Mansson, J.; Svennerholm, L. Interaction of cholera toxin and membrane GM1 ganglioside of small intestine. Proc. Natl. Acad. Sci. USA 1975, 72, 2520–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledeen, R.W.; Wu, G. The multi-tasked life of GM1 ganglioside, a true factotum of nature. Trends Biochem. Sci. 2015, 40, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J.; Geisler, F.H.; Schneider, J.S.; Li, P.A.; Fiumelli, H.; Sipione, S. Gangliosides: Treatment Avenues in Neurodegenerative Disease. Front Neurol. 2019, 10, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappagantula, S.; Andrews, M.R.; Cheah, M.; Abad-Rodriguez, J.; Dotti, C.G.; Fawcett, J.W. Neu3 sialidase-mediated ganglioside conversion is necessary for axon regeneration and is blocked in CNS axons. J. Neurosci. 2014, 34, 2477–2492. [Google Scholar] [CrossRef] [Green Version]
- Ledeen, R.; Wu, G. Gangliosides of the Nervous System. Methods Mol. Biol. 2018, 1804, 19–55. [Google Scholar] [CrossRef]
- Svennerholm, L.; Bostrom, K.; Jungbjer, B.; Olsson, L. Membrane lipids of adult human brain: lipid composition of frontal and temporal lobe in subjects of age 20 to 100 years. J. Neurochem. 1994, 63, 1802–1811. [Google Scholar] [CrossRef]
- Svennerholm, L. Ganglioside designation. Adv. Exp. Med. Biol. 1980, 125, 11. [Google Scholar] [CrossRef]
- Chester, M.A. IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature of glycolipids--recommendations 1997. Eur. J. Biochem. 1998, 257, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Schauer, R. Chemistry, metabolism, and biological functions of sialic acids. Adv. Carbohydr. Chem. Biochem. 1982, 40, 131–234. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.A. On the chemistry and occurrence of sphingolipid long-chain bases. Chem. Phys. Lipids 1970, 5, 6–43. [Google Scholar] [CrossRef]
- Chou, H.H.; Takematsu, H.; Diaz, S.; Iber, J.; Nickerson, E.; Wright, K.L.; Muchmore, E.A.; Nelson, D.L.; Warren, S.T.; Varki, A. A mutation in human CMP-sialic acid hydroxylase occurred after the Homo-Pan divergence. Proc. Natl. Acad. Sci. USA 1998, 95, 11751–11756. [Google Scholar] [CrossRef] [Green Version]
- Stoffyn, A.; Stoffyn, P.; Yip, M.C. Chemical structure of monosialoganglioside GM1b biosynthesized in vitro. Biochim. Biophys. Acta 1975, 409, 97–103. [Google Scholar] [CrossRef]
- Sonnino, S.; Chigorno, V. Ganglioside molecular species containing C18- and C20-sphingosine in mammalian nervous tissues and neuronal cell cultures. Biochim. Biophys. Acta 2000, 1469, 63–77. [Google Scholar] [CrossRef]
- Sonnino, S.; Cantu, L.; Corti, M.; Acquotti, D.; Venerando, B. Aggregative properties of gangliosides in solution. Chem. Phys. Lipids 1994, 71, 21–45. [Google Scholar] [CrossRef]
- Corti, M.; Degiorgio, V.; Ghidoni, R.; Sonnino, S.; Tettamanti, G. Laser-light scattering investigation of the micellar properties of gangliosides. Chem. Phys. Lipids 1980, 26, 225–238. [Google Scholar] [CrossRef]
- Ulrich-Bott, B.; Wiegandt, H. Micellar properties of glycosphingolipids in aqueous media. J. Lipid Res. 1984, 25, 1233–1245. [Google Scholar]
- Cantu, L.; Corti, M.; Sonnino, S.; Tettamanti, G. Light scattering measurements on gangliosides: dependence of micellar properties on molecular structure and temperature. Chem. Phys. Lipids 1986, 41, 315–328. [Google Scholar] [CrossRef]
- Loberto, N.; Lunghi, G.; Schiumarini, D.; Samarani, M.; Chiricozzi, E.; Aureli, M. Methods for Assay of Ganglioside Catabolic Enzymes. Methods Mol. Biol. 2018, 1804, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Roseman, S. The synthesis of complex carbohydrates by multiglycosyltransferase systems and their potential function in intercellular adhesion. Chem. Phys. Lipids 1970, 5, 270–297. [Google Scholar] [CrossRef]
- Sonnino, S.; Prinetti, A.; Mauri, L.; Chigorno, V.; Tettamanti, G. Dynamic and structural properties of sphingolipids as driving forces for the formation of membrane domains. Chem. Rev. 2006, 106, 2111–2125. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Sampaio, J.L. Membrane organization and lipid rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef]
- Grassi, S.; Chiricozzi, E.; Mauri, L.; Sonnino, S.; Prinetti, A. Sphingolipids and neuronal degeneration in lysosomal storage disorders. J. Neurochem. 2019, 148, 600–611. [Google Scholar] [CrossRef]
- Aureli, M.; Samarani, M.; Loberto, N.; Chiricozzi, E.; Mauri, L.; Grassi, S.; Schiumarini, D.; Prinetti, A.; Sonnino, S. Neuronal membrane dynamics as fine regulator of sphingolipid composition. Glycoconj J. 2018, 35, 397–402. [Google Scholar] [CrossRef]
- Sonnino, S.; Chigorno, V.; Aureli, M.; Masilamani, A.P.; Valsecchi, M.; Loberto, N.; Prioni, S.; Mauri, L.; Prinetti, A. Role of gangliosides and plasma membrane-associated sialidase in the process of cell membrane organization. Adv. Exp. Med. Biol. 2011, 705, 297–316. [Google Scholar] [CrossRef]
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef] [Green Version]
- Seyfried, T.N.; Choi, H.; Chevalier, A.; Hogan, D.; Akgoc, Z.; Schneider, J.S. Sex-Related Abnormalities in Substantia Nigra Lipids in Parkinson’s Disease. ASN Neuro. 2018, 10, 1759091418781889. [Google Scholar] [CrossRef] [Green Version]
- Svennerholm, L.; Gottfries, C.G. Membrane lipids, selectively diminished in Alzheimer brains, suggest synapse loss as a primary event in early-onset form (type I) and demyelination in late-onset form (type II). J. Neurochem. 1994, 62, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, K.; Tan, L.; Chen, Y.H.; Cao, Y.P. Alterations in cholesterol and ganglioside GM1 content of lipid rafts in platelets from patients with Alzheimer disease. Alzheimer Dis. Assoc. Disord. 2015, 29, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Desplats, P.A.; Denny, C.A.; Kass, K.E.; Gilmartin, T.; Head, S.R.; Sutcliffe, J.G.; Seyfried, T.N.; Thomas, E.A. Glycolipid and ganglioside metabolism imbalances in Huntington’s disease. Neurobiol. Dis. 2007, 27, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Ghidoni, R.; Sonnino, S.; Tettamanti, G.; Baumann, N.; Reuter, G.; Schauer, R. Isolation and characterization of a trisialoganglioside from mouse brain, containing 9-O-acetyl-N-acetylneuraminic acid. J. Biol. Chem. 1980, 255, 6990–6995. [Google Scholar]
- Tettamanti, G.; Bonali, F.; Marchesini, S.; Zambotti, V. A new procedure for the extraction, purification and fractionation of brain gangliosides. Biochim. Biophys. Acta 1973, 296, 160–170. [Google Scholar] [CrossRef]
- Svennerholm, L.; Fredman, P. A procedure for the quantitative isolation of brain gangliosides. Biochim. Biophys. Acta 1980, 617, 97–109. [Google Scholar] [CrossRef]
- Iwamori, M.; Nagai, Y. A new chromatographic approach to the resolution of individual gangliosides. Ganglioside mapping. Biochim. Biophys. Acta 1978, 528, 257–267. [Google Scholar]
- Koul, O.; Prada-Maluf, M.; McCluer, R.H.; Ullman, M.D. Rapid isolation of monosialogangliosides from bovine brain gangliosides by selective-overload chromatography. J. Lipid Res. 1991, 32, 1712–1715. [Google Scholar]
- Fronza, G.; Kirschner, G.; Acquotti, D.; Bassi, R.; Tagliavacca, L.; Sonnino, S. Synthesis and structural characterization of the dilactone derivative of GD1a ganglioside. Carbohydr. Res. 1988, 182, 31–40. [Google Scholar] [CrossRef]
- Evans, J.E.; McCluer, R.H. High pressure liquid chromatography of neutral glycosphingolipids. Biochim. Biophys. Acta 1972, 270, 565–569. [Google Scholar] [CrossRef]
- Acquotti, D.; Poppe, L.; Dabrowski, J.; Vonderlieth, C.W.; Sonnino, S.; Tettamanti, G. 3-Dimensional Structure of the Oligosaccharide Chain of Gm1 Ganglioside Revealed by a Distance-Mapping Procedure—A Rotating and Laboratory Frame Nuclear Overhauser Enhancement Investigation of Native Glycolipid in Dimethyl-Sulfoxide and in Water Dodecylphosphocholine Solutions. J. Am. Chem. Soc. 1990, 112, 7772–7778. [Google Scholar] [CrossRef] [Green Version]
- Acquotti, D.; Cantu, L.; Ragg, E.; Sonnino, S. Geometrical and Conformational Properties of Ganglioside Galnac-G(D1a), Iv(4)Galnaciv(3)Neu5acii(3)Neu5acggose(4)Cer. Eur. J. Biochem. 1994, 225, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Svennerholm, L.; Mansson, J.E.; Li, Y.T. Isolation and Structural Determination of a Novel Ganglioside, a Disialosylpentahexosylceramide from Human Brain. J. Biol. Chem. 1973, 248, 740–742. [Google Scholar] [PubMed]
- Sonnino, S.; Kirschner, G.; Ghidoni, R.; Acquotti, D.; Tettamanti, G. Preparation of Gm1 Ganglioside Molecular-Species Having Homogeneous Fatty-Acid and Long-Chain Base Moieties. J. Lipid Res. 1985, 26, 248–257. [Google Scholar] [PubMed]
- Sonnino, S.; Chigorno, V.; Tettamanti, G. Preparation of radioactive gangliosides, 3H or 14C isotopically labeled at oligosaccharide or ceramide moieties. Methods Enzymol. 2000, 311, 639–656. [Google Scholar] [CrossRef]
- Sonnino, S.; Nicolini, M.; Chigorno, V. Preparation of radiolabeled gangliosides. Glycobiology 1996, 6, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Sonnino, S.; Chigorno, V.; Acquotti, D.; Pitto, M.; Kirschner, G.; Tettamanti, G. A Photoreactive Derivative of Radiolabeled Gm1 Ganglioside—Preparation and Use to Establish the Involvement of Specific Proteins in Gm1 Uptake by Human-Fibroblasts in Culture. Biochemistry-Us. 1989, 28, 77–84. [Google Scholar] [CrossRef]
- Mauri, L.; Prioni, S.; Loberto, N.; Chigorno, V.; Prinetti, A.; Sonnino, S. Synthesis of radioactive and photoactivable ganglioside derivatives for the study of ganglioside-protein interactions. Glycoconj J. 2004, 20, 11–23. [Google Scholar] [CrossRef]
- Xia, Y.; Peng, L. Photoactivatable Lipid Probes for Studying Biomembranes by Photoaffinity Labeling. Chem. Rev. 2013, 113, 7880–7929. [Google Scholar] [CrossRef]
- Schwarzmann, G. Labeled gangliosides: their synthesis and use in biological studies. FEBS Lett. 2018, 592, 3992–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sixma, T.K.; Kalk, K.H.; Van Zanten, B.A.; Dauter, Z.; Kingma, J.; Witholt, B.; Hol, W.G. Refined structure of Escherichia coli heat-labile enterotoxin, a close relative of cholera toxin. J. Mol. Biol. 1993, 230, 890–918. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.; Lonnroth, I.; Svennerholm, L. Tissue receptor for cholera exotoxin: postulated structure from studies with GM1 ganglioside and related glycolipids. Infect. Immun. 1973, 8, 208–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuziemko, G.M.; Stroh, M.; Stevens, R.C. Cholera toxin binding affinity and specificity for gangliosides determined by surface plasmon resonance. Biochemistry-Us. 1996, 35, 6375–6384. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Mauri, L.; Ciampa, M.G.; Prinetti, A.; Sonnino, S. On the use of cholera toxin. Glycoconj J. 2018, 35, 161–163. [Google Scholar] [CrossRef]
- Cambron, L.D.; Leskawa, K.C. A sensitive method to quantitate gangliosides of the gangliotetraose series directly on chromatograms using peroxidase conjugated cholera toxin. Stain Technol. 1990, 65, 293–297. [Google Scholar] [CrossRef]
- Davidsson, P.; Fredman, P.; Svennerholm, L. Gangliosides and sulphatide in human cerebrospinal fluid: quantitation with immunoaffinity techniques. J. Chromatogr. 1989, 496, 279–289. [Google Scholar] [CrossRef]
- Hansson, H.A.; Holmgren, J.; Svennerholm, L. Ultrastructural localization of cell membrane GM1 ganglioside by cholera toxin. Proc. Natl. Acad. Sci. USA 1977, 74, 3782–3786. [Google Scholar] [CrossRef] [Green Version]
- Schwarzmann, G.; Hoffmann-Bleihauer, P.; Schubert, J.; Sandhoff, K.; Marsh, D. Incorporation of ganglioside analogues into fibroblast cell membranes. A spin-label study. Biochemistry-Us. 1983, 22, 5041–5048. [Google Scholar] [CrossRef]
- Chigorno, V.; Pitto, M.; Cardace, G.; Acquotti, D.; Kirschner, G.; Sonnino, S.; Ghidoni, R.; Tettamanti, G. Association of ganglio-sides to fibroblasts in culture: a study performed with GM1 [14C]-labelled at the sialic acid acetyl group. Glycoco J. 1985, 2, 279–291. [Google Scholar] [CrossRef]
- Riboni, L.; Prinetti, A.; Pitto, M.; Tettamanti, G. Patterns of endogenous gangliosides and metabolic processing of exogenous gangliosides in cerebellar granule cells during differentiation in culture. Neurochem. Res. 1990, 15, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Ghidoni, R.; Fiorilli, A.; Trinchera, M.; Venerando, B.; Chigorno, V.; Tettamanti, G. Uptake, cell penetration and metabolic processing of exogenously administered GM1 ganglioside in rat brain. Neurochem. Int. 1989, 15, 455–465. [Google Scholar] [CrossRef]
- Sonnino, S.; Chigorno, V.; Valsecchi, M.; Pitto, M.; Tettamanti, G. Specific ganglioside-cell protein interactions: A study performed with GM1 ganglioside derivative containing photoactivable azide and rat cerebellar granule cells in culture. Neurochem. Int. 1992, 20, 315–321. [Google Scholar] [CrossRef]
- Chigorno, V.; Valsecchi, M.; Sonnino, S. Biosynthesis of gangliosides containing C18:1 and C20:1 [3-14C]sphingosine after administrating [1-14C]palmitic acid and [1-14C]stearic acid to rat cerebellar granule cells in culture. Eur. J. Biochem. 1994, 221, 1095–1101. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.; Darvill, A.; Kinoshita, T.; et al. Symbol Nomenclature for Graphical Representations of Glycans. Glycobiology 2015, 25, 1323–1324. [Google Scholar] [CrossRef] [Green Version]
- Zoli, M.; Benfenati, F.; Pich, E.M.; Toffano, G.; Fuxe, K.; Agnati, L.F. Aspects of neural plasticity in the central nervous system-IV. Chemical anatomical studies on the aging brain. Neurochem. Int. 1990, 16, 437–449. [Google Scholar] [CrossRef]
- Mocchetti, I. Exogenous gangliosides, neuronal plasticity and repair, and the neurotrophins. Cell Mol. Life Sci. 2005, 62, 2283–2294. [Google Scholar] [CrossRef]
- Gorio, A.; Carmignoto, G.; Ferrari, G.; Marini, P.; Nunzi, M.G. Plasticity in neuronal regeneration: implications for the role of exogenous gangliosides. Birth Defects Orig. Artic. Ser. 1983, 19, 157–174. [Google Scholar]
- Lipartiti, M.; Lazzaro, A.; Zanoni, R.; Mazzari, S.; Toffano, G.; Leon, A. Monosialoganglioside GM1 reduces NMDA neurotoxicity in neonatal rat brain. Exp. Neurol. 1991, 113, 301–305. [Google Scholar] [CrossRef]
- Ceccarelli, B.; Aporti, F.; Finesso, M. Effects of brain gangliosides on functional recovery in experimental regeneration and reinnervation. Adv. Exp. Med. Biol. 1976, 71, 275–293. [Google Scholar] [CrossRef]
- Bose, B.; Osterholm, J.L.; Kalia, M. Ganglioside-induced regeneration and reestablishment of axonal continuity in spinal cord-transected rats. Neurosci. Lett. 1986, 63, 165–169. [Google Scholar] [CrossRef]
- Facci, L.; Leon, A.; Toffano, G.; Sonnino, S.; Ghidoni, R.; Tettamanti, G. Promotion of neuritogenesis in mouse neuroblastoma cells by exogenous gangliosides. Relationship between the effect and the cell association of ganglioside GM1. J. Neurochem. 1984, 42, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Ledeen, R.W. Biology of gangliosides: neuritogenic and neuronotrophic properties. J. Neurosci. Res. 1984, 12, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Katoh-Semba, R.; Varon, S. GM1 ganglioside accelerates neurite outgrowth from primary peripheral and central neurons under selected culture conditions. Brain Res. 1985, 355, 19–26. [Google Scholar] [CrossRef]
- Garofalo, L.; Ribeiro-da-Silva, A.; Cuello, A.C. Potentiation of nerve growth factor-induced alterations in cholinergic fibre length and presynaptic terminal size in cortex of lesioned rats by the monosialoganglioside GM1. Neuroscience 1993, 57, 21–40. [Google Scholar] [CrossRef]
- Liberini, P.; Pioro, E.P.; Maysinger, D.; Ervin, F.R.; Cuello, A.C. Long-term protective effects of human recombinant nerve growth factor and monosialoganglioside GM1 treatment on primate nucleus basalis cholinergic neurons after neocortical infarction. Neuroscience 1993, 53, 625–637. [Google Scholar] [CrossRef]
- Hadjiconstantinou, M.; Neff, N.H. GM1 ganglioside: in vivo and in vitro trophic actions on central neurotransmitter systems. J. Neurochem. 1998, 70, 1335–1345. [Google Scholar] [CrossRef]
- Newburn, E.N.; Duchemin, A.M.; Neff, N.H.; Hadjiconstantinou, M. GM1 ganglioside enhances Ret signaling in striatum. J. Neurochem. 2014, 130, 541–554. [Google Scholar] [CrossRef]
- Bachis, A.; Rabin, S.J.; Del Fiacco, M.; Mocchetti, I. Gangliosides prevent excitotoxicity through activation of TrkB receptor. Neurotox Res. 2002, 4, 225–234. [Google Scholar] [CrossRef]
- Rabin, S.J.; Bachis, A.; Mocchetti, I. Gangliosides activate Trk receptors by inducing the release of neurotrophins. J. Biol. Chem. 2002, 277, 49466–49472. [Google Scholar] [CrossRef] [Green Version]
- Rabin, S.J.; Mocchetti, I. GM1 ganglioside activates the high-affinity nerve growth factor receptor trkA. J. Neurochem. 1995, 65, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, T.; Franklin, T.; Pearl, D.K.; Yates, A.J. Ganglioside GM1 enhances induction by nerve growth factor of a putative dimer of TrkA. J. Neurochem. 1997, 68, 2348–2355. [Google Scholar] [CrossRef] [PubMed]
- Mutoh, T.; Tokuda, A.; Miyadai, T.; Hamaguchi, M.; Fujiki, N. Ganglioside GM1 binds to the Trk protein and regulates receptor function. Proc. Natl. Acad. Sci. USA 1995, 92, 5087–5091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitto, M.; Mutoh, T.; Kuriyama, M.; Ferraretto, A.; Palestini, P.; Masserini, M. Influence of endogenous GM1 ganglioside on TrkB activity, in cultured neurons. FEBS Lett. 1998, 439, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.S.; Hasegawa, T.; Miyagi, T.; Dotti, C.G.; Abad-Rodriguez, J. Asymmetric membrane ganglioside sialidase activity specifies axonal fate. Nat. Neurosci. 2005, 8, 606–615. [Google Scholar] [CrossRef]
- Duchemin, A.M.; Ren, Q.; Mo, L.; Neff, N.H.; Hadjiconstantinou, M. GM1 ganglioside induces phosphorylation and activation of Trk and Erk in brain. J. Neurochem. 2002, 81, 696–707. [Google Scholar] [CrossRef]
- Guirland, C.; Suzuki, S.; Kojima, M.; Lu, B.; Zheng, J.Q. Lipid rafts mediate chemotropic guidance of nerve growth cones. Neuron 2004, 42, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Mihara, T.; Ueda, A.; Hirayama, M.; Takeuchi, T.; Yoshida, S.; Naito, K.; Yamamoto, H.; Mutoh, T. Detection of new anti-neutral glycosphingolipids antibodies and their effects on Trk neurotrophin receptors. FEBS Lett. 2006, 580, 4991–4995. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Numakawa, T.; Shimazu, K.; Koshimizu, H.; Hara, T.; Hatanaka, H.; Mei, L.; Lu, B.; Kojima, M. BDNF-induced recruitment of TrkB receptor into neuronal lipid rafts: roles in synaptic modulation. J. Cell Biol. 2004, 167, 1205–1215. [Google Scholar] [CrossRef] [Green Version]
- Hibbert, A.P.; Kramer, B.M.; Miller, F.D.; Kaplan, D.R. The localization, trafficking and retrograde transport of BDNF bound to p75NTR in sympathetic neurons. Mol. Cell Neurosci. 2006, 32, 387–402. [Google Scholar] [CrossRef]
- Mojsilovic-Petrovic, J.; Jeong, G.B.; Crocker, A.; Arneja, A.; David, S.; Russell, D.S.; Kalb, R.G. Protecting motor neurons from toxic insult by antagonism of adenosine A2a and Trk receptors. J. Neurosci. 2006, 26, 9250–9263. [Google Scholar] [CrossRef]
- Pereira, D.B.; Chao, M.V. The tyrosine kinase Fyn determines the localization of TrkB receptors in lipid rafts. J. Neurosci. 2007, 27, 4859–4869. [Google Scholar] [CrossRef]
- Duchemin, A.M.; Ren, Q.; Neff, N.H.; Hadjiconstantinou, M. GM1-induced activation of phosphatidylinositol 3-kinase: involvement of Trk receptors. J. Neurochem. 2008, 104, 1466–1477. [Google Scholar] [CrossRef] [PubMed]
- Mo, L.; Ren, Q.; Duchemin, A.M.; Neff, N.H.; Hadjiconstantinou, M. GM1 and ERK signaling in the aged brain. Brain Res. 2005, 1054, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Mutoh, T.; Hamano, T.; Tokuda, A.; Kuriyama, M. Unglycosylated Trk protein does not co-localize nor associate with ganglioside GM1 in stable clone of PC12 cells overexpressing Trk (PCtrk cells). Glycoconj J. 2000, 17, 233–237. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Biase, E.D.; Maggioni, M.; Lunghi, G.; Fazzari, M.; Pome, D.Y.; Casellato, R.; Loberto, N.; Mauri, L.; Sonnino, S. GM1 promotes TrkA-mediated neuroblastoma cell differentiation by occupying a plasma membrane domain different from TrkA. J. Neurochem. 2019, 149, 231–241. [Google Scholar] [CrossRef]
- Mutoh, T.; Hamano, T.; Yano, S.; Koga, H.; Yamamoto, H.; Furukawa, K.; Ledeen, R.W. Stable transfection of GM1 synthase gene into GM1-deficient NG108-15 cells, CR-72 cells, rescues the responsiveness of Trk-neurotrophin receptor to its ligand, NGF. Neurochem. Res. 2002, 27, 801–806. [Google Scholar] [CrossRef]
- Orlando, P.; Cocciante, G.; Ippolito, G.; Massari, P.; Roberti, S.; Tettamanti, G. The fate of tritium labeled GM1 ganglioside injected in mice. Pharmacol. Res. Commun. 1979, 11, 759–773. [Google Scholar] [CrossRef]
- Tettamanti, G.; Venerando, B.; Roberti, S.; Chigorno, V.; Sonnino, S.; Ghidoni, R.; Orlando, P.; Massari, P. The fate of exogenously administered brain gangliosides. In Gangliosides in Neurological and Neuromuscular Function, Development and Repair; Rapport, M.M., Gorio, A., Eds.; Raven Press: New York, NY, USA, 1981; pp. 225–240. [Google Scholar]
- Lang, W. Pharmakinetic studies with 3H-labeled exogenous gangliosides injected intramuscularly into rats. In Gangliosides in Neuromuscular Function, Development and Repair; Rapport, M.M., Gorio, A., Eds.; Raven Press: New York, NY, USA, 1981; pp. 241–251. [Google Scholar]
- Zalutsky, M.; Magistretti, P.; Gallagher, P.; Ghidoni, R. Radioiodinated ganglioside GMI: a potential tool for the investigation of ganglioside function in vivo. Pharmac. Res. Commun. 1986, 17, 897410. [Google Scholar]
- Willibald, C.J.; Rosner, H.; Schwarzmann, G.; Sandhoff, K.; Rahmann, H. Axonal transport of intraocularly injected [3H-Sph]-GD1a in the chicken optic system and the fate of the exogenous ganglioside distributed by blood. Neurosci. Res. 1988, 5, 361–379. [Google Scholar] [CrossRef]
- Masco, D.; Seifert, W. Incorporation of ganglioside GM1 into rat brain after intraventricular administration. Neurosci. Res. Commun. 1988, 3, 141–149. [Google Scholar]
- Forsayeth, J.; Hadaczek, P. Ganglioside Metabolism and Parkinson’s Disease. Front. Neurosci. 2018, 12, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svennerholm, L.; Brane, G.; Karlsson, I.; Lekman, A.; Ramstrom, I.; Wikkelso, C. Alzheimer disease—Effect of continuous intracerebroventricular treatment with GM1 ganglioside and a systematic activation programme. Dement Geriatr. Cogn Disord 2002, 14, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ledeen, R.W.; Wu, G. Gangliosides, alpha-Synuclein, and Parkinson’s Disease. Prog. Mol. Biol. Transl. Sci. 2018, 156, 435–454. [Google Scholar] [CrossRef]
- Wu, G.; Lu, Z.H.; Kulkarni, N.; Ledeen, R.W. Deficiency of ganglioside GM1 correlates with Parkinson’s disease in mice and humans. J. Neurosci. Res. 2012, 90, 1997–2008. [Google Scholar] [CrossRef]
- Svennerholm, L.; Bostrom, K.; Fredman, P.; Mansson, J.E.; Rosengren, B.; Rynmark, B.M. Human brain gangliosides: developmental changes from early fetal stage to advanced age. Biochim. Biophys. Acta 1989, 1005, 109–117. [Google Scholar] [CrossRef]
- Schneider, J.S.; Seyfried, T.N.; Choi, H.S.; Kidd, S.K. Intraventricular Sialidase Administration Enhances GM1 Ganglioside Expression and Is Partially Neuroprotective in a Mouse Model of Parkinson’s Disease. PLoS ONE 2015, 10, e0143351. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Pome, D.Y.; Maggioni, M.; Di Biase, E.; Parravicini, C.; Palazzolo, L.; Loberto, N.; Eberini, I.; Sonnino, S. Role of the GM1 ganglioside oligosaccharide portion in the TrkA-dependent neurite sprouting in neuroblastoma cells. J. Neurochem. 2017, 143, 645–659. [Google Scholar] [CrossRef] [Green Version]
- Chiricozzi, E.; Maggioni, M.; Di Biase, E.; Lunghi, G.; Fazzari, M.; Loberto, N.; Elisa, M.; Scalvini, F.G.; Tedeschi, G.; Sonnino, S. The Neuroprotective Role of the GM1 Oligosaccharide, II(3)Neu5Ac-Gg4, in Neuroblastoma Cells. Mol. Neurobiol. 2019, 56, 6673–6702. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Mauri, L.; Lunghi, G.; Di Biase, E.; Fazzari, M.; Maggioni, M.; Valsecchi, M.; Prioni, S.; Loberto, N.; Pomè, D.Y.; et al. Parkinson’s disease recovery by GM1 oligosaccharide treatment in the B4galnt1+/− mouse model. Sci. Rep. 2019, 9, 19330. [Google Scholar] [CrossRef] [Green Version]
- Maggioni, M.; Di Biase, E.; Lunghi, G.; Sevin, E.; Gosselet, F.; Chiricozzi, E.; Sonnino, S. Characterization of GM1 oligosaccharide transport across the blood-brain-barrier. FEBS Open Bio. 2018, 8, 14–17. [Google Scholar]
- Horowitz, S.H. Ganglioside (Cronassial) therapy in diabetic neuropathy. Adv. Exp. Med. Biol. 1984, 174, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M.; Flood, T.; Slater, N.; Dambrosia, J. Trial of ganglioside therapy for diabetic neuropathy. Muscle Nerve 1987, 10, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Bradley, W.G.; Badger, G.J.; Tandan, R.; Fillyaw, M.J.; Young, J.; Fries, T.J.; Krusinski, P.B.; Witarsa, M.; Boerman, J.; Blair, C.J. Double-blind controlled trials of Cronassial in chronic neuromuscular diseases and ataxia. Neurology 1988, 38, 1731–1739. [Google Scholar] [CrossRef]
- Sobolewski, P. Cronassial in the treatment of neuropathies and atrophy of the optic nerve. Klin Oczna 1992, 94, 57–58. [Google Scholar]
- Siagoside. GM1, Sygen. Drugs R D 1999, 1, 36–37. [Google Scholar] [CrossRef]
- Candelise, L.; Ciccone, A. Gangliosides for acute ischaemic stroke. Cochrane Database Syst. Rev. 2011, CD000094. [Google Scholar] [CrossRef]
- Sonnino, S.; Chiricozzi, E.; Ciampa, M.G.; Mauri, L.; Prinetti, A.; Toffano, G.; Aureli, M. Serum Antibodies to Glycans in Peripheral Neuropathies. Mol. Neurobiol. 2017, 54, 1564–1567. [Google Scholar] [CrossRef]
- Yu, R.K.; Usuki, S.; Ariga, T. Ganglioside molecular mimicry and its pathological roles in Guillain-Barre syndrome and related diseases. Infect Immun. 2006, 74, 6517–6527. [Google Scholar] [CrossRef] [Green Version]
- Gottfries, C.G. Therapy options in Alzheimer’s disease. Br. J. Clin Pract. 1994, 48, 327–330. [Google Scholar]
- Augustinsson, L.E.; Blennow, K.; Blomstrand, C.; Brane, G.; Ekman, R.; Fredman, P.; Karlsson, I.; Kihlgren, M.; Lehmann, W.; Lekman, A.; et al. Intracerebroventricular administration of GM1 ganglioside to presenile Alzheimer patients. Dement Geriatr. Cogn Disord. 1997, 8, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Sendek, S.; Daskalakis, C.; Cambi, F. GM1 ganglioside in Parkinson’s disease: Results of a five year open study. J. Neurol. Sci. 2010, 292, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Gollomp, S.M.; Sendek, S.; Colcher, A.; Cambi, F.; Du, W. A randomized, controlled, delayed start trial of GM1 ganglioside in treated Parkinson’s disease patients. J. Neurol. Sci. 2013, 324, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Geisler, F.H.; Coleman, W.P.; Grieco, G.; Poonian, D.; Sygen Study Group. The Sygen multicenter acute spinal cord injury study. Spine 2001, 26, S87–S98. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Roeltgen, D.P.; Rothblat, D.S.; Chapas-Crilly, J.; Seraydarian, L.; Rao, J. GM1 ganglioside treatment of Parkinson’s disease: an open pilot study of safety and efficacy. Neurology 1995, 45, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.B.; Lang, A.; Poewe, W. Toward a redefinition of Parkinson’s disease. Mov. Disord 2012, 27, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Hadaczek, P.; Wu, G.; Sharma, N.; Ciesielska, A.; Bankiewicz, K.; Davidow, A.L.; Lu, Z.H.; Forsayeth, J.; Ledeen, R.W. GDNF signaling implemented by GM1 ganglioside; failure in Parkinson’s disease and GM1-deficient murine model. Exp. Neurol. 2015, 263, 177–189. [Google Scholar] [CrossRef]
- Schneider, J.S. Altered expression of genes involved in ganglioside biosynthesis in substantia nigra neurons in Parkinson’s disease. PLoS ONE 2018, 13, e0199189. [Google Scholar] [CrossRef]
- Schengrund, C.L. Gangliosides: Glycosphingolipids essential for normal neural development and function. Trends Biochem. Sci. 2015, 40, 397–406. [Google Scholar] [CrossRef]
- Bartels, T.; Kim, N.C.; Luth, E.S.; Selkoe, D.J. N-alpha-acetylation of alpha-synuclein increases its helical folding propensity, GM1 binding specificity and resistance to aggregation. PLoS ONE 2014, 9, e103727. [Google Scholar] [CrossRef] [Green Version]
- Martinez, Z.; Zhu, M.; Han, S.; Fink, A.L. GM1 specifically interacts with alpha-synuclein and inhibits fibrillation. Biochemistry-Us. 2007, 46, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Aras, R.; Williams, C.K.; Koprich, J.B.; Brotchie, J.M.; Singh, V. GM1 Ganglioside Modifies alpha-Synuclein Toxicity and is Neuroprotective in a Rat alpha-Synuclein Model of Parkinson’s Disease. Sci. Rep. 2019, 9, 8362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, J.S.; Cambi, F.; Gollomp, S.M.; Kuwabara, H.; Brasic, J.R.; Leiby, B.; Sendek, S.; Wong, D.F. GM1 ganglioside in Parkinson’s disease: Pilot study of effects on dopamine transporter binding. J. Neurol. Sci. 2015, 356, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, B.S.; Monahan, A.J.; Carvey, P.M.; Hendey, B. Blood-brain barrier pathology in Alzheimer’s and Parkinson’s disease: Implications for drug therapy. Cell Transpl. 2007, 16, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Erdo, F.; Denes, L.; De Lange, E. Age-associated physiological and pathological changes at the blood-brain barrier: A review. J. Cereb. Blood Flow Metab. 2017, 37, 4–24. [Google Scholar] [CrossRef] [Green Version]
- Saulino, M.F.; Schengrund, C.L. Differential accumulation of gangliosides by the brains of MPTP-lesioned mice. J. Neurosci. Res. 1994, 37, 384–391. [Google Scholar] [CrossRef]
- Polo, A.; Kirschner, G.; Guidotti, A.; Costa, E. Brain content of glycosphingolipids after oral administration of monosialogangliosides GM1 and LIGA20 to rats. Mol. Chem. Neuropathol. 1994, 21, 41–53. [Google Scholar] [CrossRef]
- Ghidoni, R.; Trinchera, M.; Venerando, B.; Fiorilli, A.; Sonnino, S.; Tettamanti, G. Incorporation and metabolism of exogenous GM1 ganglioside in rat liver. Biochem. J. 1986, 237, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Amaro, M.; Sachl, R.; Aydogan, G.; Mikhalyov, I.I.; Vacha, R.; Hof, M. GM1 Ganglioside Inhibits beta-Amyloid Oligomerization Induced by Sphingomyelin. Angew Chem. Int. Ed Engl. 2016, 55, 9411–9415. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, A.; Harrison, F.E.; McCord, M.; Zhao, J.; Bruchey, A.; Davies, S.S.; Jackson Roberts, L., 2nd; Mathews, P.M.; Matsuoka, Y.; Ariga, T.; et al. Elimination of GD3 synthase improves memory and reduces amyloid-beta plaque load in transgenic mice. Neurobiol. Aging 2009, 30, 1777–1791. [Google Scholar] [CrossRef]
- Yang, R.; Wang, Q.; Min, L.; Sui, R.; Li, J.; Liu, X. Monosialoanglioside improves memory deficits and relieves oxidative stress in the hippocampus of rat model of Alzheimer’s disease. Neurol. Sci. 2013, 34, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Zhang, X.; Li, L.; Chen, H.; Chai, Y. Autophagy Inhibition Contributes to ROS-Producing NLRP3-Dependent Inflammasome Activation and Cytokine Secretion in High Glucose-Induced Macrophages. Cell Physiol. Biochem. 2017, 43, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; Furukawa, J.; Fujitani, N.; Shinohara, Y.; Igarashi, Y. Decreased amyloid-beta pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J. Biol. Chem. 2014, 289, 24488–24498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagisawa, K. GM1 ganglioside and Alzheimer’s disease. Glycoconj J. 2015, 32, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Yamamoto, N.; Yanagisawa, K. Accelerated release of exosome-associated GM1 ganglioside (GM1) by endocytic pathway abnormality: another putative pathway for GM1-induced amyloid fibril formation. J. Neurochem. 2008, 105, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Matsubara, T.; Sato, T.; Yanagisawa, K. Age-dependent high-density clustering of GM1 ganglioside at presynaptic neuritic terminals promotes amyloid beta-protein fibrillogenesis. Biochim. Biophys. Acta 2008, 1778, 2717–2726. [Google Scholar] [CrossRef] [Green Version]
- Sandhoff, R.; Schulze, H.; Sandhoff, K. Ganglioside Metabolism in Health and Disease. Prog. Mol. Biol. Transl. Sci. 2018, 156, 1–62. [Google Scholar] [CrossRef]
- Sonnino, S.; Chiricozzi, E.; Grassi, S.; Mauri, L.; Prioni, S.; Prinetti, A. Gangliosides in Membrane Organization. Prog. Mol. Biol. Transl. Sci. 2018, 156, 83–120. [Google Scholar] [CrossRef]
- Prinetti, A.; Chigorno, V.; Prioni, S.; Loberto, N.; Marano, N.; Tettamanti, G.; Sonnino, S. Changes in the lipid turnover, composition, and organization, as sphingolipid-enriched membrane domains, in rat cerebellar granule cells developing in vitro. J. Biol. Chem. 2001, 276, 21136–21145. [Google Scholar] [CrossRef] [Green Version]
- Prinetti, A.; Prioni, S.; Chiricozzi, E.; Schuchman, E.H.; Chigorno, V.; Sonnino, S. Secondary alterations of sphingolipid metabolism in lysosomal storage diseases. Neurochem. Res. 2011, 36, 1654–1668. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Ciampa, M.G.; Brasile, G.; Compostella, F.; Prinetti, A.; Nakayama, H.; Ekyalongo, R.C.; Iwabuchi, K.; Sonnino, S.; Mauri, L. Direct interaction, instrumental for signaling processes, between LacCer and Lyn in the lipid rafts of neutrophil-like cells. J. Lipid Res. 2015, 56, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariga, T.; McDonald, M.P.; Yu, R.K. Role of ganglioside metabolism in the pathogenesis of Alzheimer’s disease—A review. J. Lipid Res. 2008, 49, 1157–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molander-Melin, M.; Blennow, K.; Bogdanovic, N.; Dellheden, B.; Mansson, J.E.; Fredman, P. Structural membrane alterations in Alzheimer brains found to be associated with regional disease development; increased density of gangliosides GM1 and GM2 and loss of cholesterol in detergent-resistant membrane domains. J. Neurochem. 2005, 92, 171–182. [Google Scholar] [CrossRef]
- Svennerholm, L.; Gottfries, C.G.; Blennow, K.; Fredman, P.; Karlsson, I.; Mansson, J.E.; Toffano, G.; Wallin, A. Parenteral administration of GM1 ganglioside to presenile Alzheimer patients. Acta Neurol. Scand 1990, 81, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Geisler, F.H.; Dorsey, F.C.; Coleman, W.P. Recovery of motor function after spinal-cord injury—A randomized, placebo-controlled trial with GM-1 ganglioside. N. Engl. J. Med. 1991, 324, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Geisler, F.H.; Dorsey, F.C.; Coleman, W.P. Correction: Recovery of motor function after spinal-cord injury--a randomized, placebo-controlled trial with GM-1 ganglioside. N. Engl. J. Med. 1991, 325, 1659–1660. [Google Scholar]
- Hadley, M.N.; Walters, B.C.; Grabb, P.A.; Oyesiku, N.M.; Przybylski, G.J.; Resnick, D.K.; Ryken, T.C.; Mielke, D.H. Guidelines for the management of acute cervical spine and spinal cord injuries. Clin Neurosurg. 2002, 49, 407–498. [Google Scholar] [CrossRef]
- Walters, B.C.; Hadley, M.N. Guidelines for GM-1 ganglioside in acute spinal cord injury. Neurosurgery 2013, 73, E752. [Google Scholar] [CrossRef]
- Walters, B.C.; Hadley, M.N.; Hurlbert, R.J.; Aarabi, B.; Dhall, S.S.; Gelb, D.E.; Harrigan, M.R.; Rozelle, C.J.; Ryken, T.C.; Theodore, N.; et al. Guidelines for the management of acute cervical spine and spinal cord injuries: 2013 update. Neurosurgery 2013, 60, 82–91. [Google Scholar] [CrossRef]
- Walker, J.B.; Harris, M. GM-1 ganglioside administration combined with physical therapy restores ambulation in humans with chronic spinal cord injury. Neurosci. Lett. 1993, 161, 174–178. [Google Scholar] [CrossRef]
- Papo, I.; Benedetti, A.; Carteri, A.; Merli, G.A.; Mingrino, S.; Bruno, R. Monosialoganglioside in subarachnoid hemorrhage. Stroke 1991, 22, 22–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoermann, M. Efficacy and safety of ganglioside GM1 treatment in the rehabilitation of patients following closed traumatic head injury. Results of an interim evaluation. New Trends Ganglioside Res. Neurochem. Neurodegener. Asp. 1988, 14, 595–604. [Google Scholar]
- Li, L.; Tian, J.; Long, M.K.; Chen, Y.; Lu, J.; Zhou, C.; Wang, T. Protection against Experimental Stroke by Ganglioside GM1 Is Associated with the Inhibition of Autophagy. PLoS ONE 2016, 11, e0144219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Krafft, P.R.; Wang, T.; Zhang, J.H.; Li, L.; Tang, J. Pathophysiology of Ganglioside GM1 in Ischemic Stroke: Ganglioside GM1: A Critical Review. Cell Transplant. 2019, 28, 657–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khwaja, O.; Volpe, J.J. Pathogenesis of cerebral white matter injury of prematurity. Arch Dis. Child-Fetal 2008, 93, F153–F161. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.C.V.; De Silva, D.A.; Macleod, M.R.; Coutts, S.B.; Schwamm, L.H.; Davis, S.M.; Donnan, G.A. Ischaemic stroke. Nat. Rev. Dis. Primers 2019, 5, 70. [Google Scholar] [CrossRef]
- Wang, X.; Tian, X.; Ma, J.; Zheng, J. Clinical efficacy of gangliosides on premature infants suffering from white matter damage and its effect on the levels of IL-6, NSE and S100beta. Exp. Ther. Med. 2019, 18, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Hagberg, H.; David Edwards, A.; Groenendaal, F. Perinatal brain damage: The term infant. Neurobiol. Dis. 2016, 92, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.Y.; Ye, M.Y.; Zhang, A.M.; Wang, W.D.; Zeng, F.; Li, J.L.; Fang, F. Influence of one-year neurologic outcome of treatment on newborns with moderate and severe hypoxic-ischemic encephalopathy by rhuEP0 combined with ganglioside (GM1). Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3955–3960. [Google Scholar]
- Whitehead, S.N.; Chan, K.H.; Gangaraju, S.; Slinn, J.; Li, J.; Hou, S.T. Imaging mass spectrometry detection of gangliosides species in the mouse brain following transient focal cerebral ischemia and long-term recovery. PLoS ONE 2011, 6, e20808. [Google Scholar] [CrossRef]
- Kwak, D.H.; Kim, S.M.; Lee, D.H.; Kim, J.S.; Kim, S.M.; Lee, S.U.; Jung, K.Y.; Seo, B.B.; Choo, Y.K. Differential expression patterns of gangliosides in the ischemic cerebral cortex produced by middle cerebral artery occlusion. Mol. Cells 2005, 20, 354–360. [Google Scholar] [PubMed]
- Su, D.; Ma, J.; Yang, J.; Kang, Y.; Lv, M.; Li, Y. Monosialotetrahexosy-1 ganglioside attenuates diabetes-associated cerebral ischemia/reperfusion injury through suppression of the endoplasmic reticulum stress-induced apoptosis. J. Clin. Neurosci. 2017, 41, 54–59. [Google Scholar] [CrossRef]
- Zhang, J.; Fang, X.; Zhou, Y.; Deng, X.; Lu, Y.; Li, J.; Li, S.; Wang, B.; Xu, R. The Possible Damaged Mechanism and the Preventive Effect of Monosialotetrahexosylganglioside in a Rat Model of Cerebral Ischemia-Reperfusion Injury. J. Stroke Cerebrovasc. Dis. 2015, 24, 1471–1478. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Jing, L.; Ma, Y.; Guo, F.Y.; Chang, Y.; Li, P.A. Monosialotetrahexosy-1 ganglioside attenuates diabetes-enhanced brain damage after transient forebrain ischemia and suppresses phosphorylation of ERK1/2 in the rat brain. Brain Res. 2010, 1344, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.R.; Ding, M.P.; Wei, E.Q.; Luo, J.H.; Song, Y.; Huang, J.Z.; Ge, Q.F.; Hu, H.; Zhu, L.J. GM1 stabilizes expression of NMDA receptor subunit 1 in the ischemic hemisphere of MCAo/reperfusion rat. J. Zhejiang Univ. Sci. B 2005, 6, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.P.; Chen, J.; Graham, S.H. GM1 ganglioside treatment of focal ischemia: A dose-response and microdialysis study. J. Pharmacol. Exp. Ther. 1993, 265, 24–29. [Google Scholar] [PubMed]
- Bharucha, V.A.; Wakade, C.G.; Mahadik, S.P.; Karpiak, S.E. GM1 ganglioside treatment reduces functional deficits associated with cortical focal ischemia. Exp. Neurol. 1991, 114, 136–139. [Google Scholar] [CrossRef]
- Alter, M. GM1 ganglioside for acute ischemic stroke. Trial design issues. Ann N. Y. Acad. Sci. 1998, 845, 391–401. [Google Scholar] [CrossRef]
- Lenzi, G.L.; Grigoletto, F.; Gent, M.; Roberts, R.S.; Walker, M.D.; Easton, J.D.; Carolei, A.; Dorsey, F.C.; Rocca, W.A.; Bruno, R.; et al. Early treatment of stroke with monosialoganglioside GM-1. Efficacy and safety results of the Early Stroke Trial. Stroke 1994, 25, 1552–1558. [Google Scholar] [CrossRef] [Green Version]
- Argentino, C.; Sacchetti, M.L.; Toni, D.; Savoini, G.; D’Arcangelo, E.; Erminio, F.; Federico, F.; Milone, F.F.; Gallai, V.; Gambi, D.; et al. GM1 ganglioside therapy in acute ischemic stroke. Italian Acute Stroke Study—Hemodilution + Drug. Stroke 1989, 20, 1143–1149. [Google Scholar] [CrossRef] [Green Version]
- Alter, M.; Bell, R.; Brass, L.; Gaines, K.; Goldstein, L.; Hollander, J.; Jozefczyk, P.; Kelley, R.; Mayman, C.; Miller, A.; et al. Ganglioside Gm(1) in Acute Ischemic Stroke—The Sass Trial. Stroke 1994, 25, 1141–1148. [Google Scholar]
- Scarpino, O.; Martinazzo, C.; Magi, M.; Bruno, R. GM1 Ganglioside therapy in acute ischemic stroke. In Cerebral Ischemia and Dementia; Hartmann, A., Kuschinsky, W., Hoyer, S., Eds.; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Choucry, A.M.; Al-Shorbagy, M.Y.; Attia, A.S.; El-Abhar, H.S. Pharmacological Manipulation of Trk, p75NTR, and NGF Balance Restores Memory Deficit in Global Ischemia/Reperfusion Model in Rats. J. Mol. Neurosci. 2019, 68, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Sonnino, S.; Prinetti, A. Membrane domains and the “lipid raft” concept. Curr. Med. Chem. 2013, 20, 4–21. [Google Scholar] [PubMed]
- Wu, G.; Lu, Z.H.; Kulkarni, N.; Amin, R.; Ledeen, R.W. Mice lacking major brain gangliosides develop parkinsonism. Neurochem. Res. 2011, 36, 1706–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Tommee, C.; Vidal-Martinez, G.; Annette Reyes, C.; Vargas-Medrano, J.; Herrera, G.V.; Martin, S.M.; Chaparro, S.A.; Perez, R.G. Parkinsonian GM2 synthase knockout mice lacking mature gangliosides develop urinary dysfunction and neurogenic bladder. Exp. Neurol. 2019, 311, 265–273. [Google Scholar] [CrossRef]
- Vidal-Martinez, G.; Najera, K.; Miranda, J.D.; Gil-Tommee, C.; Yang, B.; Vargas-Medrano, J.; Diaz-Pacheco, V.; Perez, R.G. FTY720 Improves Behavior, Increases Brain Derived Neurotrophic Factor Levels and Reduces alpha-Synuclein Pathology in Parkinsonian GM2+/- Mice. Neuroscience 2019, 411, 1–10. [Google Scholar] [CrossRef]
- Sonnino, S.; Mauri, L.; Ciampa, M.G.; Prinetti, A. Gangliosides as regulators of cell signaling: Ganglioside-protein interactions or ganglioside-driven membrane organization? J. Neurochem. 2013, 124, 432–435. [Google Scholar] [CrossRef]
- Sonnino, S.; Aureli, M.; Grassi, S.; Mauri, L.; Prioni, S.; Prinetti, A. Lipid rafts in neurodegeneration and neuroprotection. Mol. Neurobiol. 2014, 50, 130–148. [Google Scholar] [CrossRef]
- Schengrund, C.L.; Prouty, C. Oligosaccharide portion of GM1 enhances process formation by S20Y neuroblastoma cells. J. Neurochem. 1988, 51, 277–282. [Google Scholar] [CrossRef]
- Di Biase, E.; Lunghi, G.; Fazzari, M.; Prioni, S.; Chiricozzi, E.; Sonnino, S. Neurotrophic properties of GM1 oligosaccharide: Evidence on the development of the primary neurons in culture. In Special Issue on Glycoconj J., Proceedings of the 25th International Symposium on Glycoconjugates, Milano, Italy, 25–31 August 2019; Springer: New York, NY, USA, 2019; Volume 36, p. 267. [Google Scholar]
- Ferrari, G.; Anderson, B.L.; Stephens, R.M.; Kaplan, D.R.; Greene, L.A. Prevention of apoptotic neuronal death by GM1 ganglioside. Involvement of Trk neurotrophin receptors. J. Biol. Chem. 1995, 270, 3074–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.A.; Piddini, E.; Hasegawa, T.; Miyagi, T.; Dotti, C.G. Plasma membrane ganglioside sialidase regulates axonal growth and regeneration in hippocampal neurons in culture. J. Neurosci. 2001, 21, 8387–8395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantini, J.; Yahi, N. Lipid Regulation of Receptor Function. Brain Lipids Synaptic Funct. Neurol. Dis. 2015, 163–181. [Google Scholar]
- Abeliovich, A.; Schmitz, Y.; Farinas, I.; Choi-Lundberg, D.; Ho, W.H.; Castillo, P.E.; Shinsky, N.; Verdugo, J.M.; Armanini, M.; Ryan, A.; et al. Mice lacking alpha-synuclein display functional deficits in the nigrostriatal dopamine system. Neuron 2000, 25, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Nemani, V.M.; Lu, W.; Berge, V.; Nakamura, K.; Onoa, B.; Lee, M.K.; Chaudhry, F.A.; Nicoll, R.A.; Edwards, R.H. Increased expression of alpha-synuclein reduces neurotransmitter release by inhibiting synaptic vesicle reclustering after endocytosis. Neuron 2010, 65, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Das, U.; Scott, D.A.; Tang, Y.; McLean, P.J.; Roy, S. Alpha-synuclein multimers cluster synaptic vesicles and attenuate recycling. Curr. Biol. 2014, 24, 2319–2326. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Reitbock, P.; Anichtchik, O.; Bellucci, A.; Iovino, M.; Ballini, C.; Fineberg, E.; Ghetti, B.; Della Corte, L.; Spano, P.; Tofaris, G.K.; et al. SNARE protein redistribution and synaptic failure in a transgenic mouse model of Parkinson’s disease. Brain 2010, 133, 2032–2044. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trivial Accepted/Used Name | IUPAC-IUB Nomenclature | Chemical Structure |
|---|---|---|
| AGM1 (to discourage) | Gg4Cer | β-Gal-(1-3)-β-GalNAc-(1-4)-β-Gal-(1-4)-β-Glc-(1-1)-Cer |
| GM1a | II3Neu5Ac-Gg4Cer | β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-3)-]β-Gal-(1-4)-β-Glc-(1-1)-Cer |
| GD1a | II3Neu5Ac,IV3Neu5Ac-Gg4Cer | α-Neu5Ac-(2-3)-β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-3)-]β-Gal-(1-4)-β-Glc-(1-1)-Cer |
| GD1b | II3Neu5Ac2-Gg4Cer | β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-8)-α-Neu5Ac-(2-3)-]β-Gal-(1-4)-β-Glc-(1-1)-Cer |
| GT1b | II3Neu5Ac2,IV3Neu5Ac-Gg4Cer | α-Neu5Ac-(2-3)-β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-α-Neu5Ac-(2-3)-]β-Gal-(1-4)-β-Glc-(1-1)-Cer |
| GQ1b | II3Neu5Ac2,IV3Neu5Ac2-Gg4Cer | α-Neu5Ac-(2-8)-α-Neu5Ac-(2-3)-β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-α-Neu5Ac-(2-3)-]β-Gal-(1-4)-β-Glc-(1-1)-Cer |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiricozzi, E.; Lunghi, G.; Di Biase, E.; Fazzari, M.; Sonnino, S.; Mauri, L. GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration. Int. J. Mol. Sci. 2020, 21, 868. https://doi.org/10.3390/ijms21030868
Chiricozzi E, Lunghi G, Di Biase E, Fazzari M, Sonnino S, Mauri L. GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration. International Journal of Molecular Sciences. 2020; 21(3):868. https://doi.org/10.3390/ijms21030868
Chicago/Turabian StyleChiricozzi, Elena, Giulia Lunghi, Erika Di Biase, Maria Fazzari, Sandro Sonnino, and Laura Mauri. 2020. "GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration" International Journal of Molecular Sciences 21, no. 3: 868. https://doi.org/10.3390/ijms21030868
APA StyleChiricozzi, E., Lunghi, G., Di Biase, E., Fazzari, M., Sonnino, S., & Mauri, L. (2020). GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration. International Journal of Molecular Sciences, 21(3), 868. https://doi.org/10.3390/ijms21030868