Effects of Vitamin D Treatment on Skeletal Muscle Histology and Ultrastructural Changes in a Rodent Model

{kind=link}
{kind=link}
Abstract
:1. Introduction
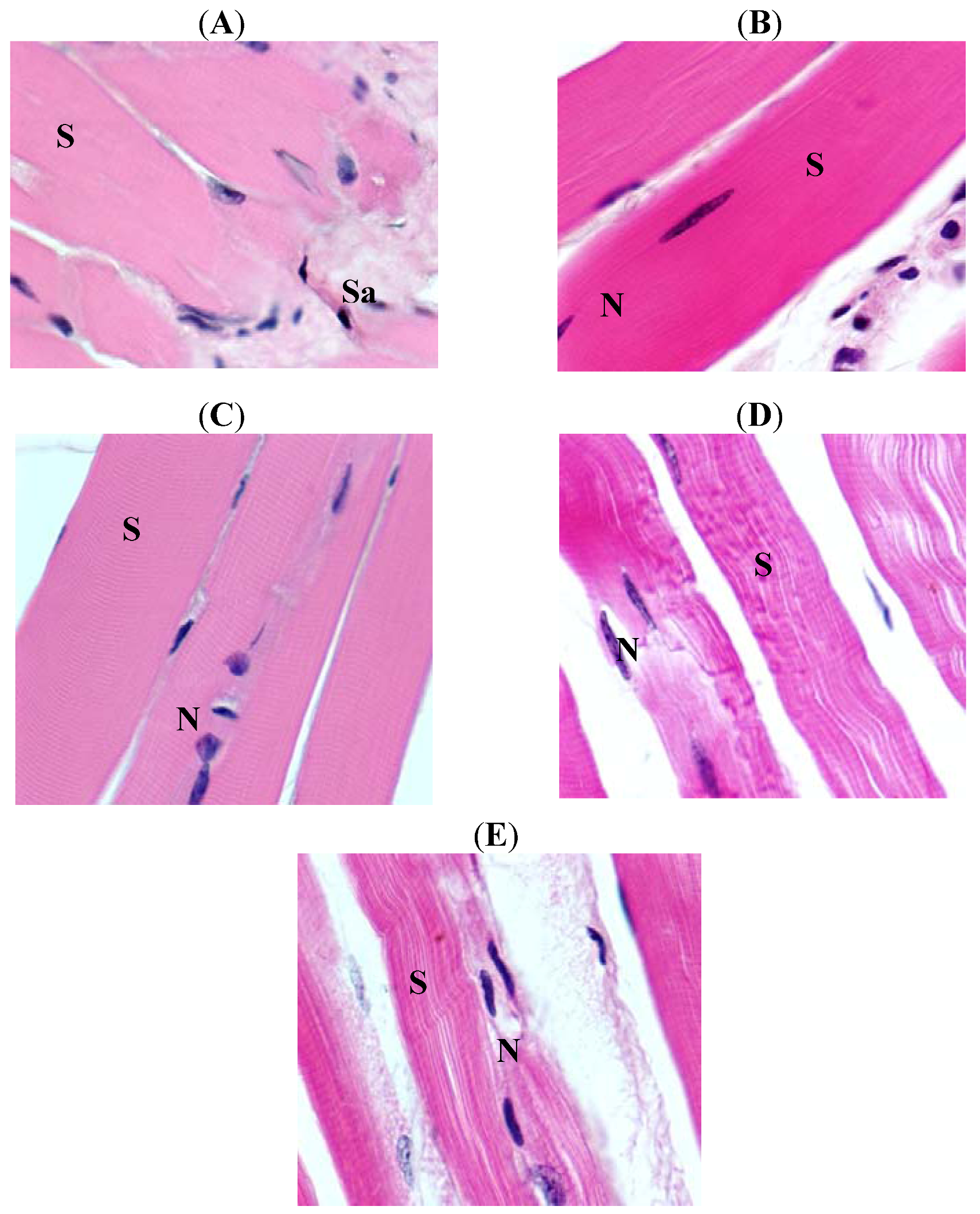
2. Results
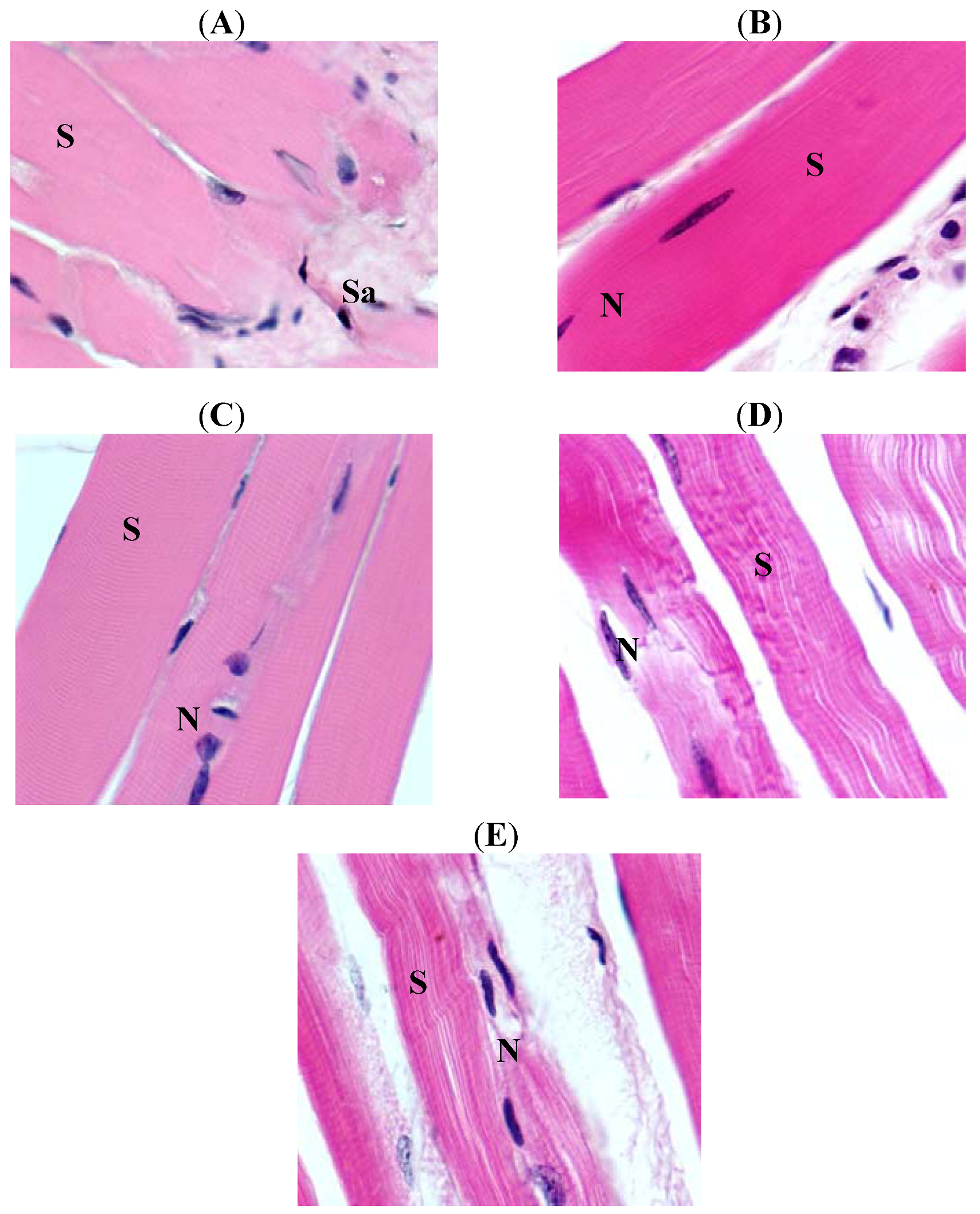

3. Discussion
4. Experimental
4.1. Animals and Study Protocol
4.2. Histology and Transmission Electronic Microscope Studies
4.3. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Gomez-Perez, Y.; Capllonch-Amer, G.; Gianotti, M.; Llado, I.; Proenza, A.M. Long-term high-fat-diet feeding induces skeletal muscle mitochondrial biogenesis in rats in a sex-dependent and muscle-type specific manner. Nutr. Metab. (Lond) 2012, 9, 15. [Google Scholar] [CrossRef]
- Chanseaume, E.; Morio, B. Potential mechanisms of muscle mitochondrial dysfunction in aging and obesity and cellular consequences. Int. J. Mol. Sci. 2009, 10, 306–324. [Google Scholar] [CrossRef]
- Ostronoff, L.K.; Izquierdo, J.M.; Enriquez, J.A.; Montoya, J.; Cuezva, J.M. Transient activation of mitochondrial translation regulates the expression of the mitochondrial genome during mammalian mitochondrial differentiation. Biochem. J. 1996, 316, 183–191. [Google Scholar]
- Johannsen, D.L.; Ravussin, E. The role of mitochondria in health and disease. Curr. Opin. Pharmacol. 2009, 9, 780–786. [Google Scholar] [CrossRef]
- van den Broek, N.M.; Ciapaite, J.; De Feyter, H.M.; Houten, S.M.; Wanders, R.J.; Jeneson, J.A.; Nicolay, K.; Prompers, J.J. Increased mitochondrial content rescues in vivo muscle oxidative capacity in long-term high-fat-diet-fed rats. FASEB J. 2010, 24, 1354–1364. [Google Scholar] [CrossRef]
- Abdul-Ghani, M.A.; DeFronzo, R.A. Pathogenesis of insulin resistance in skeletal muscle. J. Biomed. Biotechnol. 2010, 19. Article ID 476279.. [Google Scholar]
- Barazzoni, R.; Zanetti, M.; Semolic, A.; Cattin, M.R.; Pirulli, A.; Cattin, L.; Guarnieri, G. High-fat diet with acyl-ghrelin treatment leads to weight gain with low inflammation, high oxidative capacity and normal triglycerides in rat muscle. PLoS One 2011, 6, e26224. [Google Scholar]
- Choi, K.; Kim, Y.B. Molecular mechanism of insulin resistance in obesity and type 2 diabetes. Korean J. Intern. Med. 2010, 25, 119–129. [Google Scholar] [CrossRef]
- Ridker, P.M.; Hennekens, C.H.; Buring, J.E.; Rifai, N. C-reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women. N. Engl. J. Med. 2000, 342, 836–843. [Google Scholar] [CrossRef]
- Prineas, J.; Mason, A.; Henson, R. Myopathy in metabolic bone disease. Br. Med. J. 1965, 1, 1034–1036. [Google Scholar]
- Smith, R.; Stern, G. Myopathy, osteomalacia and hyperparathyroidism. Brain 1967, 90, 593–602. [Google Scholar] [CrossRef]
- Astrup, A.; Buemann, B.; Western, P.; Toubro, S.; Raben, A.; Christensen, N.J. Obesity as an adaptation to a high-fat diet: Evidence from a cross-sectional study. Am. J. Clin. Nutr. 1994, 59, 350–355. [Google Scholar]
- Kim, J.K.; Kim, Y.J.; Fillmore, J.J.; Chen, Y.; Moore, I.; Lee, J.; Yuan, M.; Li, Z.W.; Karin, M.; Perret, P.; Shoelson, S.E.; Shulman, G.I. Prevention of fat-induced insulin resistance by salicylate. J. Clin. Invest. 2001, 108, 437–446. [Google Scholar]
- Sims, E.A. Storage and expenditure of energy in obesity and their implications for management. Med. Clin. N. Am. 1989, 73, 97–110. [Google Scholar]
- Ahren, B. Plasma leptin and insulin in C57BI/6J mice on a high-fat diet: Relation to subsequent changes in body weight. Acta Physiol. Scand. 1999, 165, 233–240. [Google Scholar] [CrossRef]
- Safdar, A.; Hamadeh, M.J.; Kaczor, J.J.; Raha, S.; Debeer, J.; Tarnopolsky, M.A. Aberrant mitochondrial homeostasis in the skeletal muscle of sedentary older adults. PLoS One 2010, 5, e10778. [Google Scholar]
- Kim, J.; Wei, Y.; Sowers, J.R. Role of mitochondrial dysfunction in insulin resistance. Circ. Res. 2008, 102, 401–414. [Google Scholar] [CrossRef]
- Ceglia, L. Vitamin D and skeletal muscle tissue and function. Mol. Aspects Med. 2008, 29, 407–414. [Google Scholar] [CrossRef]
- Russell, J.A. Osteomalacic myopathy. Muscle Nerve 1994, 17, 578–580. [Google Scholar] [CrossRef]
- Alkharfy, K.M.; Al-Daghri, N.M.; yakout, S.M.; Ahmed, M. Vitamin D3 Attenuates Weight-Related Systemic Inflammation and Ultrastructural Changes of the Liver in a Rodent Model. Basic Clin. Pharmacol. Toxicol. 2012, unpublished work.. [Google Scholar]
- Ceglia, L. Vitamin D and its role in skeletal muscle. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 628–633. [Google Scholar] [CrossRef]
- Civitarese, A.E.; Smith, S.R.; Ravussin, E. Diet, energy metabolism and mitochondrial biogenesis. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 679–687. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kadowaki, T. Physiological and pathophysiological roles of adiponectin and adiponectin receptors in the integrated regulation of metabolic and cardiovascular diseases. Int. J. Obes. (Lond) 2008, 32, S13–S18. [Google Scholar] [CrossRef]
- Calle, C.; Maestro, B.; Garcia-Arencibia, M. Genomic actions of 1,25-dihydroxyvitamin D3 on insulin receptor gene expression, insulin receptor number and insulin activity in the kidney, liver and adipose tissue of streptozotocin-induced diabetic rats. BMC Mol. Biol. 2008, 9, 65. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Alkharfy, K.M.; Al-Daghri, N.M.; Ahmed, M.; Yakout, S.M. Effects of Vitamin D Treatment on Skeletal Muscle Histology and Ultrastructural Changes in a Rodent Model. Molecules 2012, 17, 9081-9089. https://doi.org/10.3390/molecules17089081
Alkharfy KM, Al-Daghri NM, Ahmed M, Yakout SM. Effects of Vitamin D Treatment on Skeletal Muscle Histology and Ultrastructural Changes in a Rodent Model. Molecules. 2012; 17(8):9081-9089. https://doi.org/10.3390/molecules17089081
Chicago/Turabian StyleAlkharfy, Khalid M., Nasser M. Al-Daghri, Mukhtar Ahmed, and Sobhy M. Yakout. 2012. "Effects of Vitamin D Treatment on Skeletal Muscle Histology and Ultrastructural Changes in a Rodent Model" Molecules 17, no. 8: 9081-9089. https://doi.org/10.3390/molecules17089081