Anti-Inflammatory and Antipruritic Effects of Luteolin from Perilla (P. frutescens L.) Leaves

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
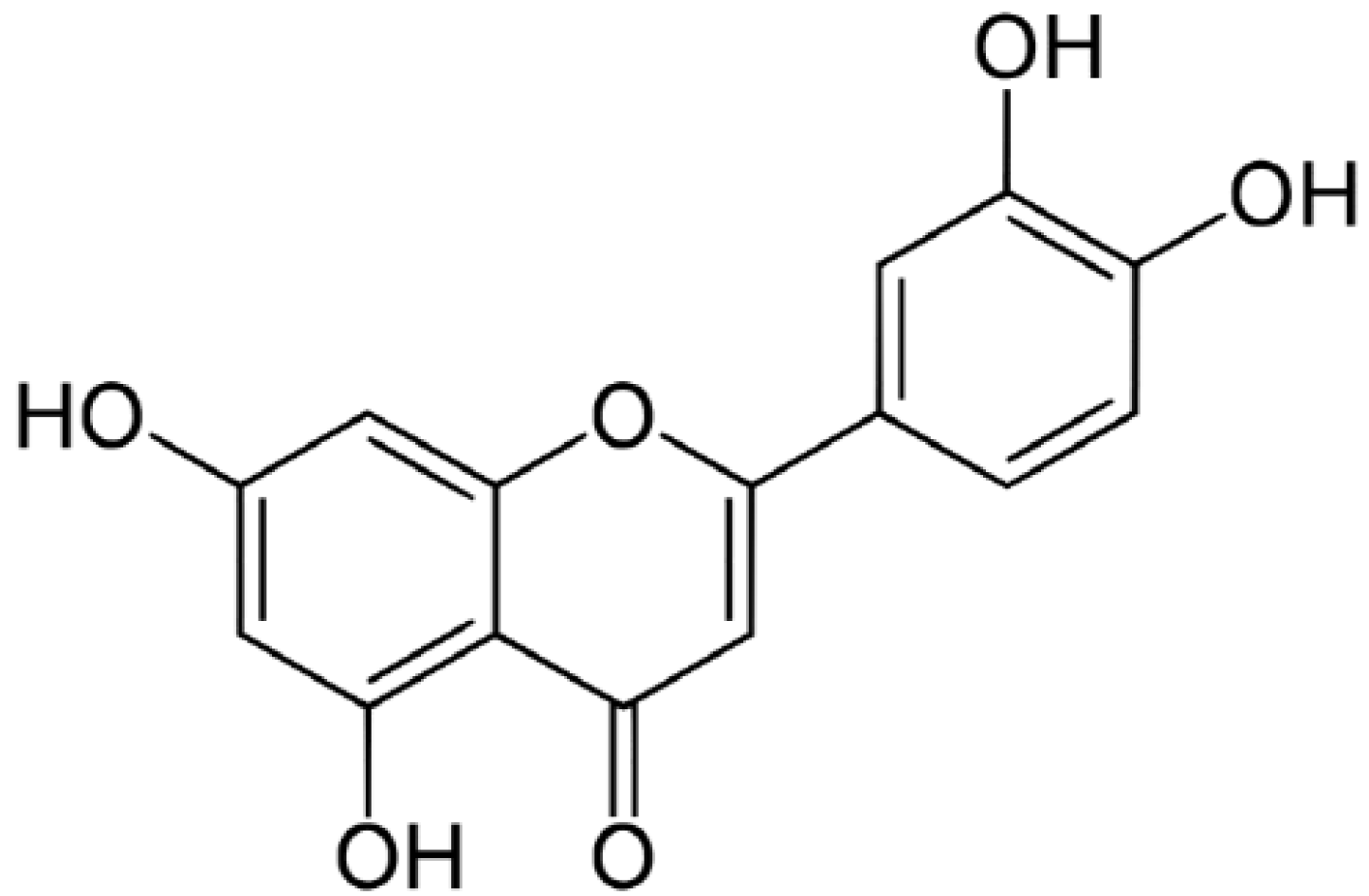
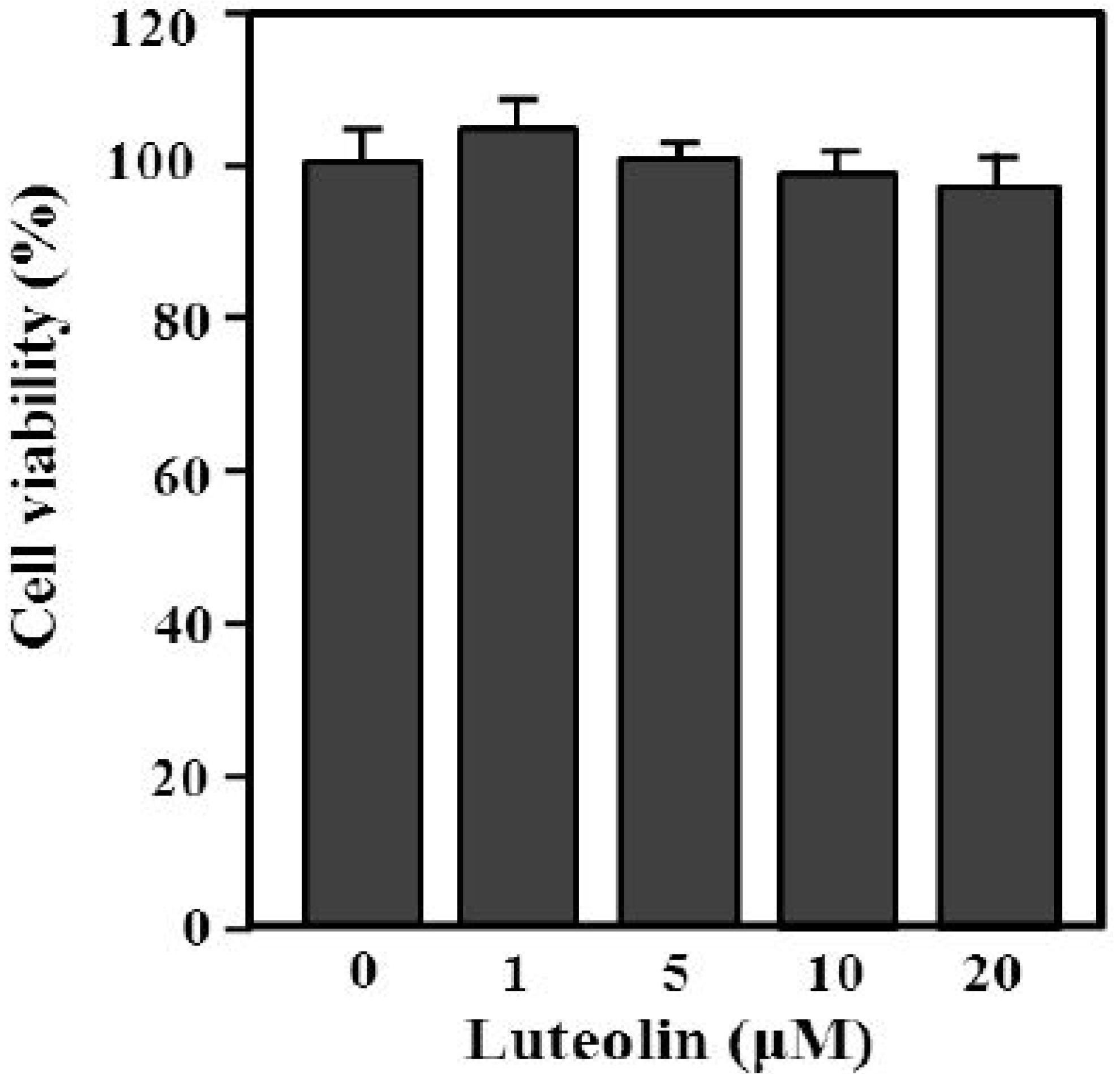
2.1. The Chemical Structure and Effect of Luteolin on Cell Survival


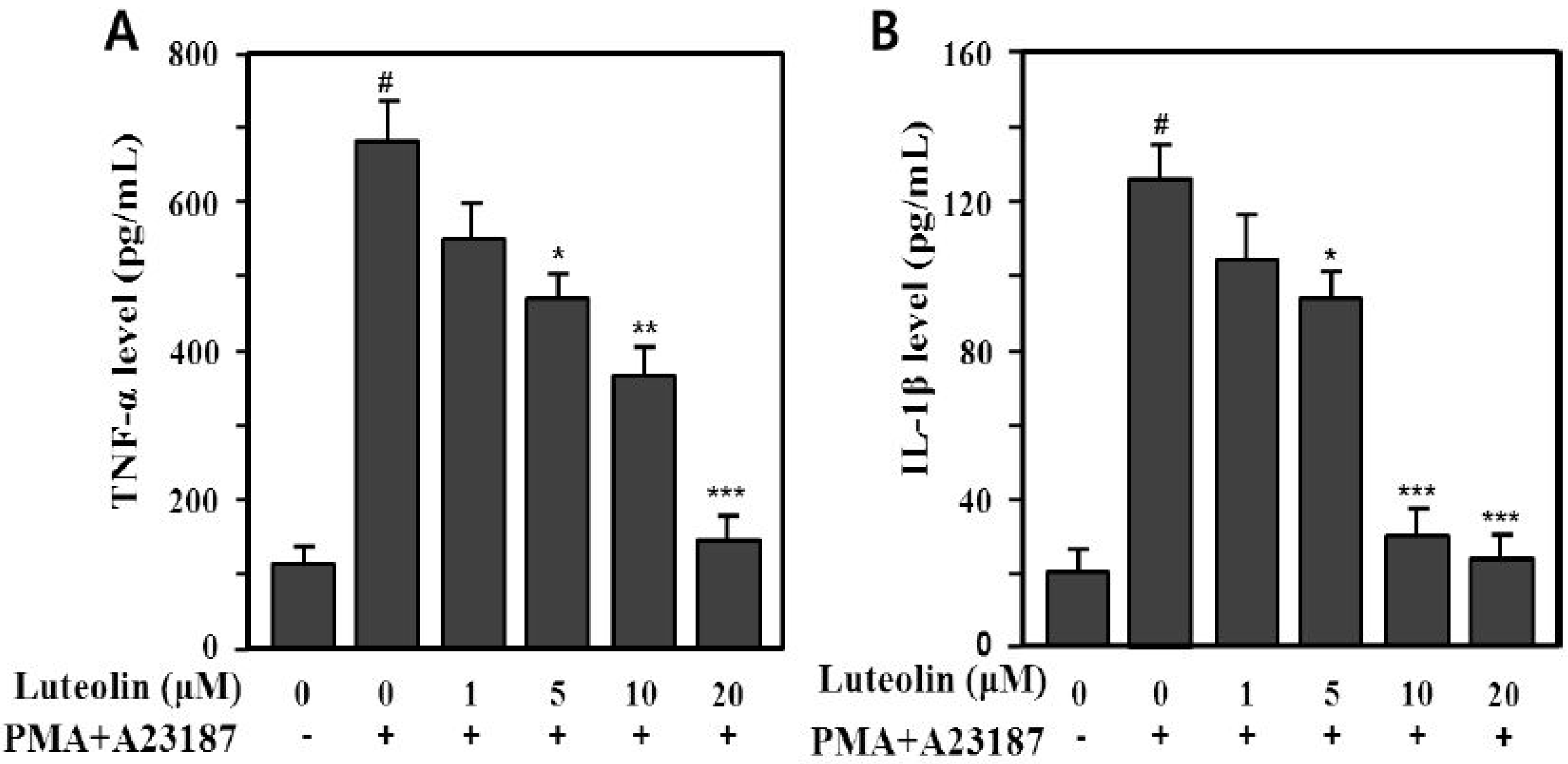
2.2. Inhibition of TNF-α and IL-1β Production by Luteolin

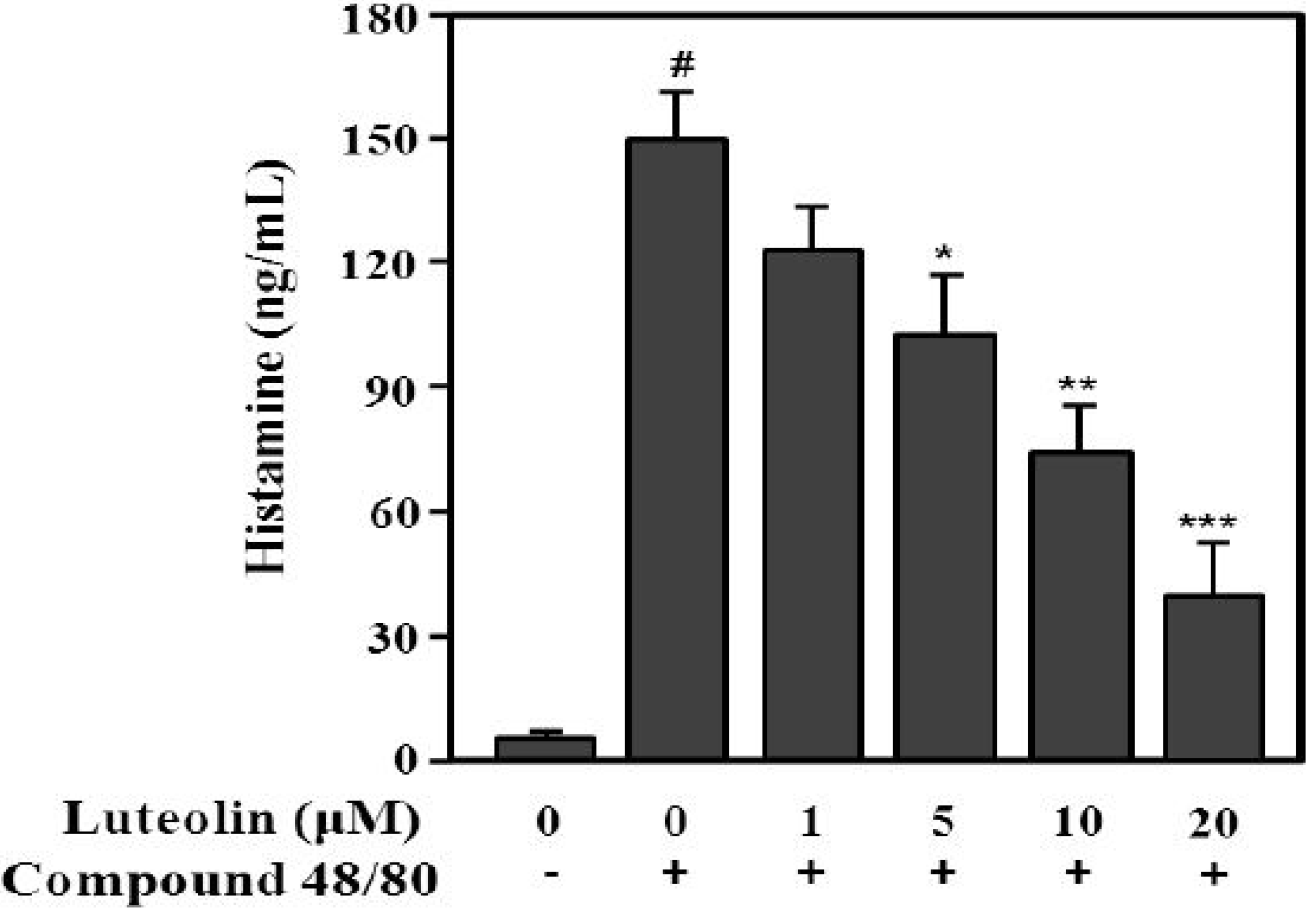
2.3. Inhibition of Histamine Release by Luteolin

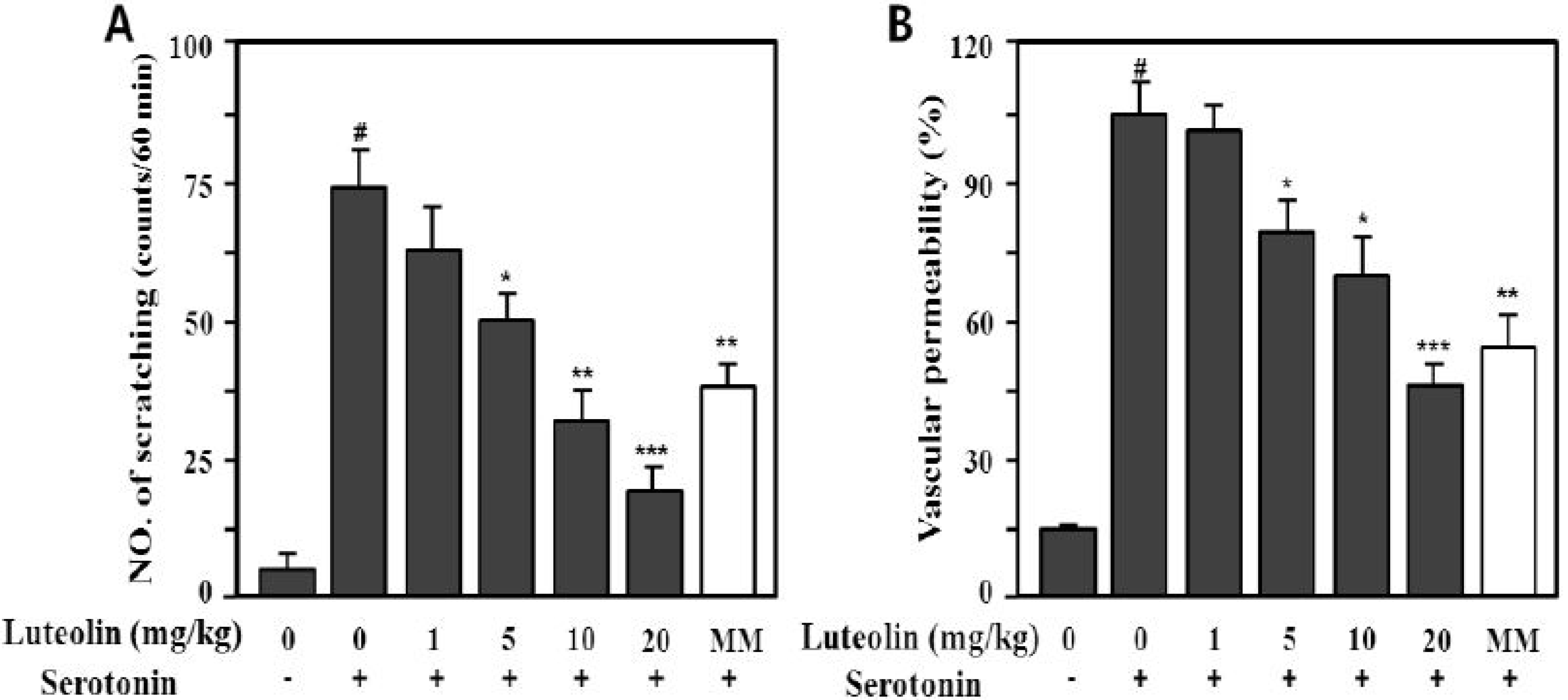
2.4. Effects of Luteolin on Scratching Behavior and Vascular Permeability


3. Experimental
3.1. Chemicals
3.2. Perilla Leaves and Luteolin Isolation
3.3. Animals
3.4. Cell Culture and Stimulation
3.5. MTT Assay
3.6. Rat Peritoneal Mast Cell (RPMC) Isolation and Stimulation
3.7. Histamine Assay
3.8. Cytokine ELISA
3.9. Scratching Behavior
3.10. Skin Vascular Permeability
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hougee, S.; Sanders, A.; Faber, J.; Graus, Y.M.; van den Berg, W.B.; Garssen, J.; Smit, H.F.; Hoijer, M.A. Decreased pro-inflammatory cytokine production by LPS-stimulated PBMC upon in vitro incubation with the flavonoids apigenin, luteolin or chrysin, due to selective elimination of monocytes/macrophages. Biochem. Pharmacol. 2005, 69, 241–248. [Google Scholar]
- Suárez, A.L.; Feramisco, J.D.; Koo, J.; Steinhoff, M. Psychoneuroimmunology of psychological stress and atopic dermatitis: Pathophysiologic and therapeutic updates. Acta Derm. Venereol. 2012, 92, 7–15. [Google Scholar]
- Gurish, M.F.; Austen, K.F. The diverse roles of mast cells. J. Exp. Med. 2001, 94, F1–F5. [Google Scholar]
- Kim, E.K.; Kim, E.Y.; Moon, P.D.; Um, J.Y.; Kim, H.M.; Lee, H.S.; Sohn, Y.; Park, S.K.; Jung, H.S.; Sohn, N.W. Lithospermi radix extract inhibits histamine release and production of inflammatory cytokine in mast cells. Biosci. Biotech. Biochem. 2007, 71, 2886–1892. [Google Scholar]
- Hosoda, M.; Yamaya, M.; Suzuki, T.; Yamada, N.; Kamanakam, M.; Sekizawa, K.; Butterfield, J.H.; Watanabe, T.; Nishimura, H.; Sasaki, H. Effects of rhinovirus infection on histamine and cytokine production by cell lines from human mast cells and basophils. J. Immunol. 2002, 169, 1482–1491. [Google Scholar]
- Rasheed, Z.; Akhtar, H.; Khan, A.; Khan, K.A.; Haqqi, T.M. Butrin, isobutrin, and butein from medicinal plant Butea monosperma selectively inhibit nuclear factor-κB in activated human mast cells: Suppression of tumor necrosis factor-α, interleukin (IL)-6, and IL-8. J. Pharmacol. Exp. Ther. 2010, 333, 354–363. [Google Scholar]
- Mekori, Y.A.; Metcalfe, D. Mast cells in innate immunity. Immunol. Rev. 2000, 173, 131–140. [Google Scholar]
- Ikoma, A.; Steinhoff, M.; Ständer, S.; Yosipovitch, G.; Schmelz, M. The neurobiology of itch. Nat. Rev. Neurosci. 2006, 7, 535–547. [Google Scholar]
- Steinhoff, M.; Bienenstock, J.; Schmelz, M.; Maurer, M.; Wei, E.; Bíró, T. Neurophysiological, neurological, and neuroendocrine basis of pruritus. J. Invest. Dermatol. 2006, 126, 1705–1718. [Google Scholar]
- Hong, J.; Buddenkotte, J.; Berger, T.G.; Steinhoff, M. Management of itch in atopic dermatitis. Semin. Cutan. Med. Surg. 2011, 30, 71–86. [Google Scholar]
- Kobayashi, Y.; Takahashi, R.; Ogino, F. Antipruritic effect of the single oral administration of German chamomile flower extract and its combined effect with antiallergic agents in ddY mice. J. Ethnopharmacol. 2005, 101, 308–312. [Google Scholar]
- Liu, J.; Wan, Y.; Zhao, Z.; Chen, H. Determination of the content of rosmarinic acid by HPLC and analytical comparison of volatile constituents by GC-MS in different parts of Perilla frutescens (L.). Britt. Chem. Cent. J. 2013, 7, 1–11. [Google Scholar]
- Kim, D.H.; Kim, Y.C.; Choi, U.K. Optimization of antibacterial activity of Perilla frutescens var. acuta leaf against Staphylococcus aureus using evolutionary operation factorial design technique. Int. J. Mol. Sci. 2011, 12, 2395–2407. [Google Scholar]
- Makino, T.; Furuta, Y.; Wakushima, H.; Fujii, H.; Saito, K.; KanoY., Y. Anti-allergic effect of Perilla frutescens and its active constituents. Phytother. Res. 2003, 17, 240–243. [Google Scholar]
- Seo, W.H.; Baek, H.H. Characteristic aroma-active compounds of Korean perilla (Perillafrutescens Britton) leaf. J. Agric. Food Chem. 2009, 57, 11537–11542. [Google Scholar]
- Ragazinskiene, O.; Gailys, V.; Jankauskiene, K.; Simoniene, G.; Jurkstiene, V. Common perilla (Perilla frutescens (L.) Britton.) as a perspective immunomodulator. Medicina (Kaunas) 2004, 40, 220–224. [Google Scholar]
- Ueda, H.; Yamazaki, C.; Yamazaki, M. Luteolin as an anti-inflammatory and anti-allergic constituent of Perilla frutescens. Biol. Pharmaceut. Bull. 2002, 25, 1197–1202. [Google Scholar]
- Haghi, G.; Hatami, A. Simultaneous quantification of flavonoids and phenolic acids in plant materials by a newly developed isocratic high-performance liquid chromatography approach. J. Agric. Food Chem. 2010, 58, 10812–10816. [Google Scholar]
- Harris, G.K.; Qian, Y.; Leonard, S.S.; Sbarra, D.C.; Shi, X. Luteolin and chrysin differentially inhibit cyclooxygenase-2 expression and scavenge reactive oxygen species but similarly inhibit prostaglandin-E2 formation in RAW 264.7 cells. J. Nutri. 2006, 136, 1517–1521. [Google Scholar]
- Choi, C.W.; Jung, H.A.; Kang, S.S.; Choi, J.S. Antioxidant constituents and a new triterpenoid glycoside from Flos Lonicera. Arch. Pharm. Res. 2007, 30, 1–7. [Google Scholar]
- Chan, B.C.; Hon, K.L.; Leung, P.C.; Sam, S.W.; Fung, K.P.; Lee, M.Y.; Lau, H.Y. Traditional Chinese medicine for atopic eczema: Penta herbs formula suppresses inflammatory mediators release from mast cells. J. Ethnopharmacol. 2008, 120, 85–91. [Google Scholar]
- Chan, C.L.; Jones, R.L.; Lau, H.Y. Characterization of prostanoid receptors mediating inhibition of histamine release from anti-IgE-activated rat peritoneal mast cells. Br. J. Pharmacol. 2000, 129, 589–597. [Google Scholar]
- Kang, O.H.; Choi, J.G.; Lee, J.H.; Kwon, D.Y. Luteolin isolated from the flowers of Lonicera japonica suppresses inflammatory mediator release by blocking NF-kappaB and APKs activation pathways in HMC-1 cells. Molecules 2010, 15, 385–398. [Google Scholar]
- Stassen, M.; Müller, C.; Arnold, M.; Hültner, L.; Klein-Hessling, S.; Neudörfl, C.; Reineke, T.; Serfling, E.; Schmitt, E. IL-9 and IL-13 production by activated mast cells is strongly enhanced in the presence of lipopolysaccharide: NF-kappa B is decisively involved in the expression of IL-9. J. Immunol. 2000, 166, 4391–4398. [Google Scholar]
- Guma, M.; Kashiwakura, J.I.; Crain, B.; Kawakami, Y.; Beutler, B.; Firestein, G.S.; Kawakami, T.; Karin, M.; Corr, M. JNK1 controls mast cell degranulation and IL-1β production in inflammatory arthritis. Proc. Natl. Acad. Sci. USA 2010, 107, 22122–22127. [Google Scholar]
- Kim, C.D.; Lee, W.K.; No, K.O.; Park, S.K.; Lee, M.H.; Lim, S.R.; Roh, S.S. Anti-allergic action of buckwheat (Fagopyrum esculentum Moench) grain extract. Int. Immunopharmacol. 2003, 3, 129–136. [Google Scholar]
- Choi, Y.H.; Chai, O.H.; Han, E.H.; Choi, S.Y.; Kim, H.T.; Song, C.H. Lipoic acid suppresses compound 48/80-induced anaphylaxis-like reaction. Anat. Cell Biol. 2010, 43, 317–324. [Google Scholar]
- Rukwied, R.; Lischetzki, G.; McGlone, F.; Heyer, G.; Schmilz, M. Mast cell mediators other than histamine induce pruritus in atopic dermatitis patterns: A dermal microdialysis study. Br. J. Pharmacol. 2000, 142, 1114–1120. [Google Scholar]
- Inagaki, N.; Igeta, K.; Kim, J.F.; Nagao, M.; Shiraishi, N.; Nakamura, N.H.; Nagai, H. Involvement of unique mechanisms in the induction of scratching behavior in BALB/c mice by compound 48/80. Eur. J. Pharmacol. 2002, 448, 175–183. [Google Scholar]
- Ohmura, T.; Hayashi, T.; Satoh, Y.; Konomi, A.; Jung, B.; Sato, H. Involvement of substance P in scratching behavior in an atopic dermatitis model. Eur. J. Pharmacol. 2004, 491, 191–194. [Google Scholar]
- Stephane, D.; Emmanuel, V.; Vincent, S.; Jacques, C.; Denis, H.; Jean-Marie, L.; Luc, M. Serotonin 5-HT2B receptors are required for 3,4-Methylenedioxymethamphetamine-induced hyperlocomotion and 5-HT release in vivo and in vitro. J. Neurosci. 2008, 28, 2933–2940. [Google Scholar]
- Takubo, M.; Ueda, Y.; Yatsuzuka, R.; Jiang, S.; Fujii, Y.; Kamei, C. Characteristics of scratching behavior induced by some chemical mediators in hairless mice. J. Pharmacol. Sci. 2006, 100, 285–288. [Google Scholar]
- Sugimoto, Y.; Umakoshi, K.; Nojiri, N.; Kamei, C. Effect of histamine H1 receptor antagonists on compound-induced scratching behavior in mice. Eur. J. Pharmacol. 1998, 351, 1–5. [Google Scholar]
- Sample Availability: Not available.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jeon, I.H.; Kim, H.S.; Kang, H.J.; Lee, H.-S.; Jeong, S.I.; Kim, S.J.; Jang, S.I. Anti-Inflammatory and Antipruritic Effects of Luteolin from Perilla (P. frutescens L.) Leaves. Molecules 2014, 19, 6941-6951. https://doi.org/10.3390/molecules19066941
Jeon IH, Kim HS, Kang HJ, Lee H-S, Jeong SI, Kim SJ, Jang SI. Anti-Inflammatory and Antipruritic Effects of Luteolin from Perilla (P. frutescens L.) Leaves. Molecules. 2014; 19(6):6941-6951. https://doi.org/10.3390/molecules19066941
Chicago/Turabian StyleJeon, In Hwa, Hyeon Soo Kim, Hyun Ju Kang, Hyun-Seo Lee, Seung Il Jeong, Sang Jun Kim, and Seon Il Jang. 2014. "Anti-Inflammatory and Antipruritic Effects of Luteolin from Perilla (P. frutescens L.) Leaves" Molecules 19, no. 6: 6941-6951. https://doi.org/10.3390/molecules19066941