
Insulin Mimetic Properties of Extracts Prepared from Bellis perennis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Transfection
2.3. Cytotoxicity Assay
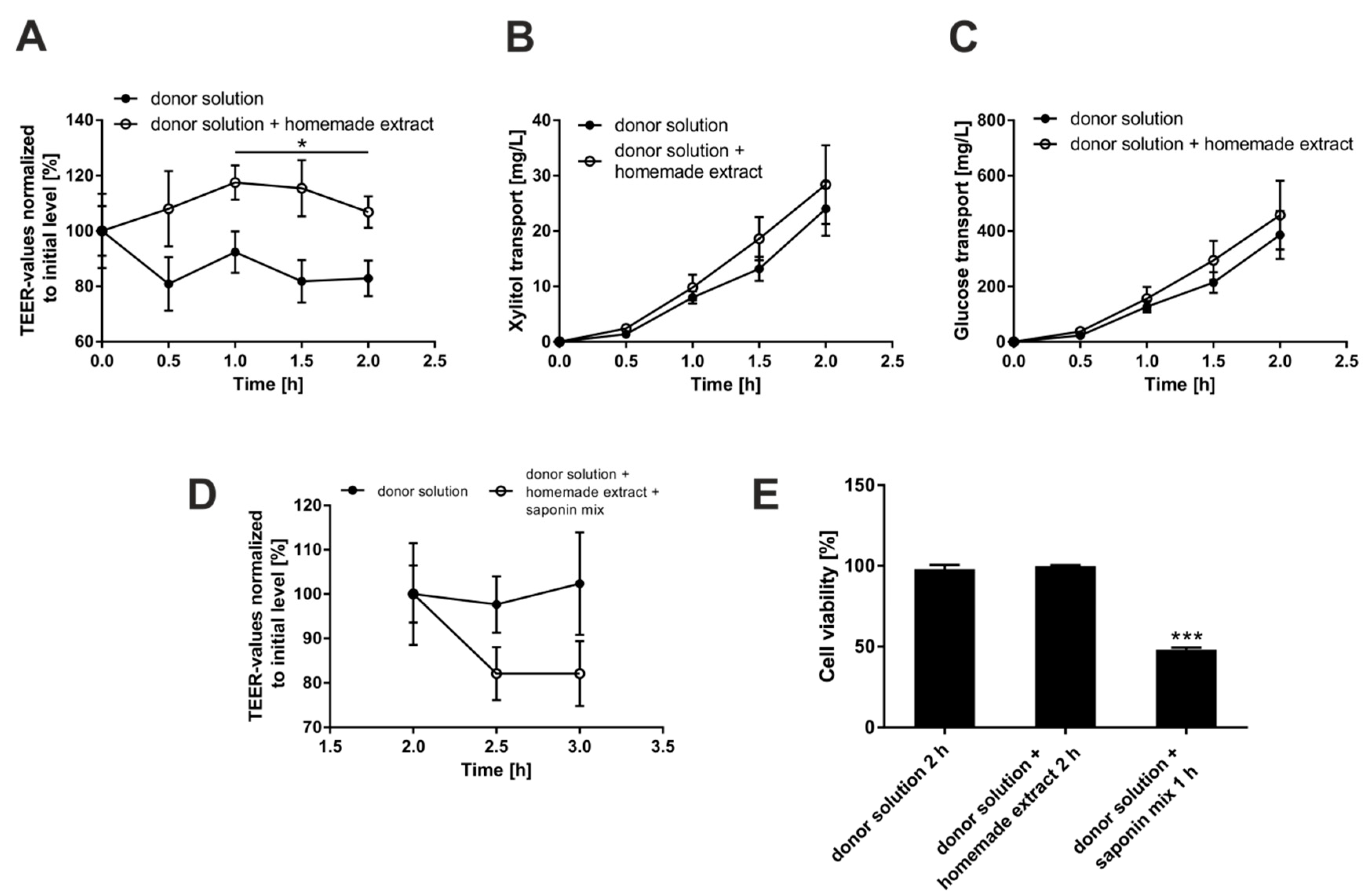
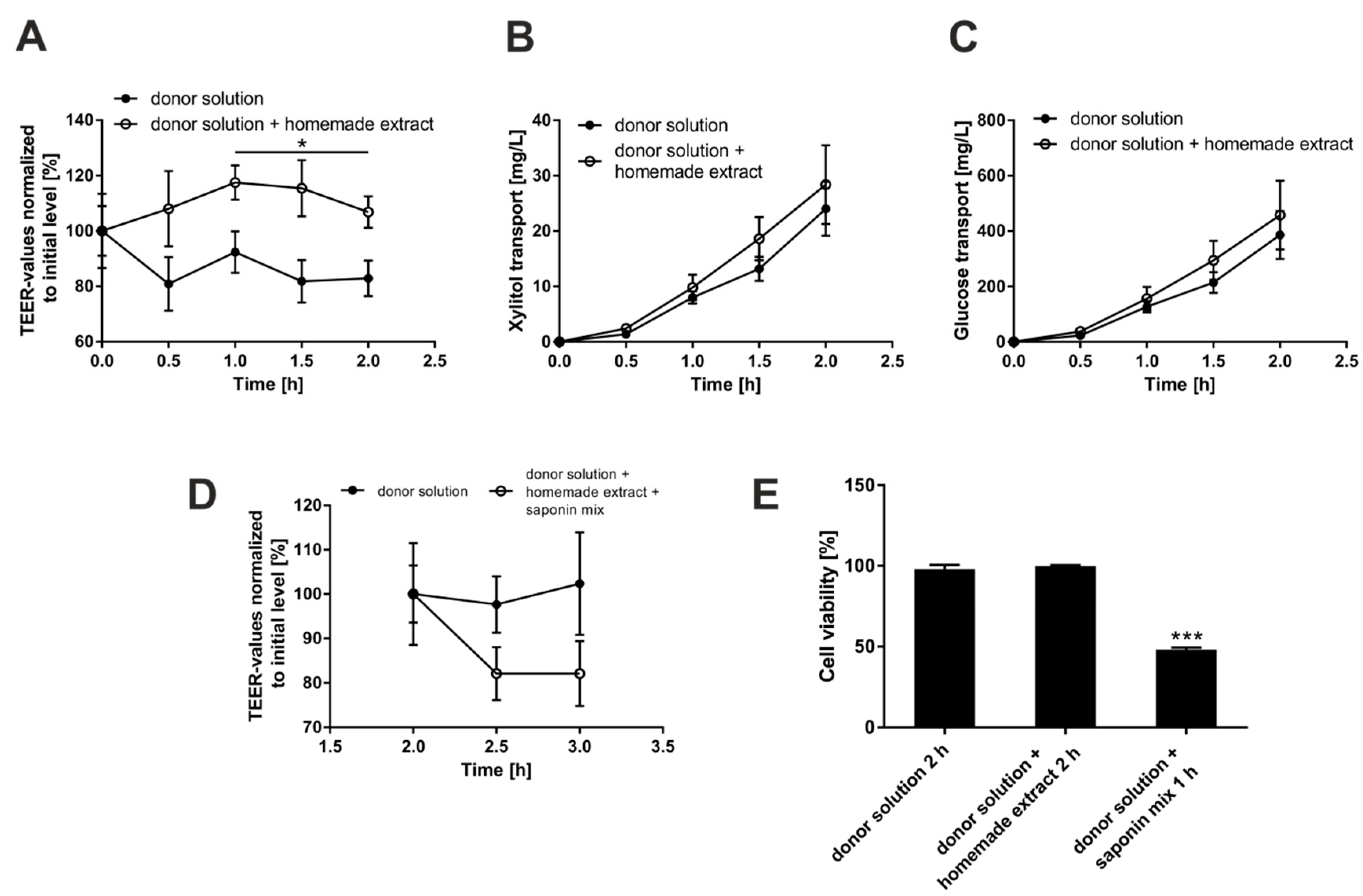
2.4. Determination of Cell Layer Integrity by Transepithelial Electrical Resistance (TEER) Measurements and Sugar Transport Quantitation
2.5. Glucose Transport Assay
2.6. Extract Preparation
2.7. Total Internal Reflection Fluorescence (TIRF) Microscopy
2.8. Hens Egg Test-Chorioallantoic Membrane (HET-CAM)
2.9. High-Performance Liquid Chromatography (HPLC) Analysis
2.10. Data Analysis
3. Results
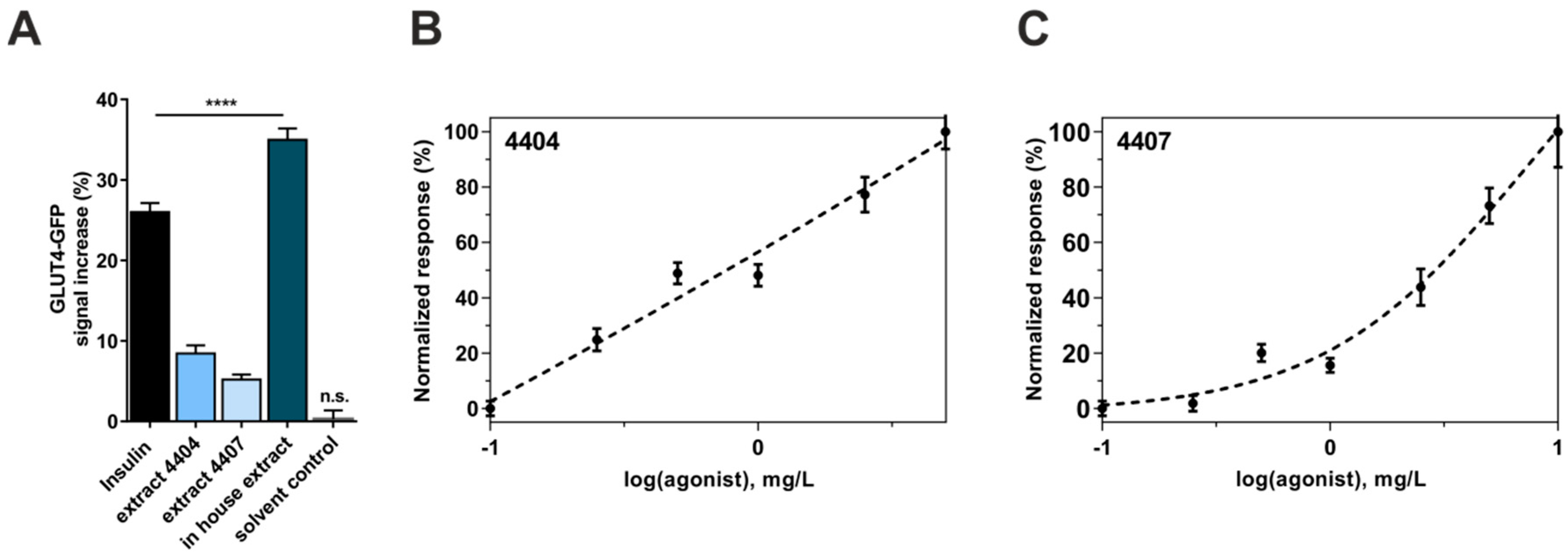
3.1. Induction of GLUT4-Translocation by Bellis Perennis Extracts
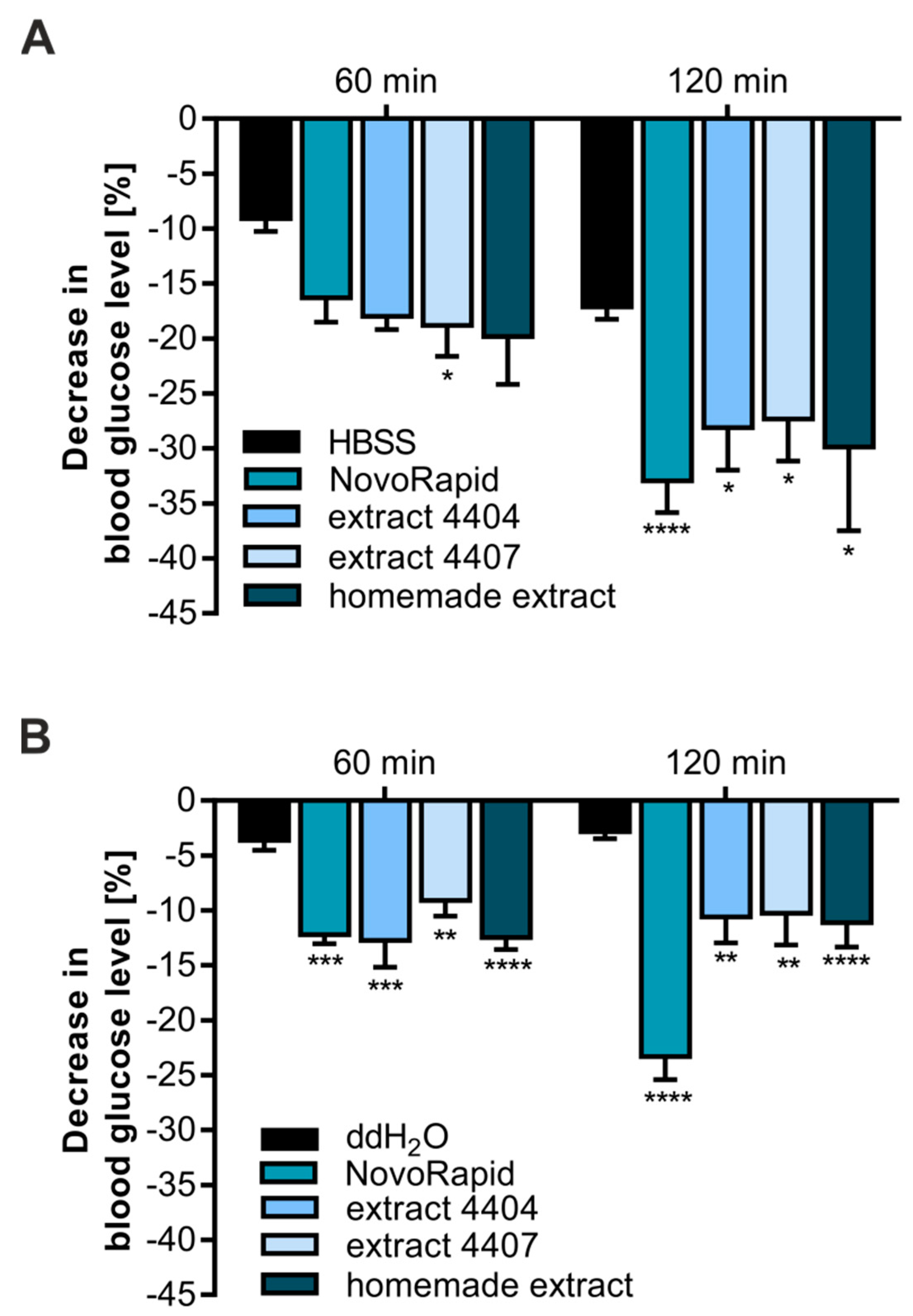
3.2. Bellis Perennis Reduces Blood Glucose Levels In Ovo
3.3. Investigation of Putative Negative Effects of Bellis Perennis Extracts on Epithelial Integrity
3.4. Identification and Quantitation of Polyphenols in Bellis Perennis Extracts
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Smyth, S.; Heron, A. Diabetes and obesity: The twin epidemics. Nat. Med. 2006, 12, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Word Health Organization. Global Report on Diabetes. Available online: http://www.who.int (accessed on 7 July 2018).
- Bommer, C.; Sagalova, V.; Heesemann, E.; Manne-Goehler, J.; Atun, R.; Barnighausen, T.; Davies, J.; Vollmer, S. Global economic burden of diabetes in adults: Projections from 2015 to 2030. Diabetes Care 2018, 41, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Chaudhuri, A.; Mohanty, P.; Garg, R. Metabolic syndrome: A comprehensive perspective based on interactions between obesity, diabetes, and inflammation. Circulation 2005, 111, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Alberti, K.G.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2010, 375, 181–183. [Google Scholar] [CrossRef]
- Scheen, A.J. Outcomes and lessons from the PROactive study. Diabetes Res. Clin. Pract. 2012, 98, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Kirpichnikov, D.; McFarlane, S.I.; Sowers, J.R. Metformin: An update. Ann. Intern. Med. 2002, 137, 25–33. [Google Scholar] [PubMed]
- List, J.F.; Woo, V.; Morales, E.; Tang, W.; Fiedorek, F.T. Sodium-glucose cotransport inhibition with dapagliflozin in type 2 diabetes. Diabetes Care 2009, 32, 650–657. [Google Scholar] [PubMed]
- Aleman-Gonzalez-Duhart, D.; Tamay-Cach, F.; Alvarez-Almazan, S.; Mendieta-Wejebe, J.E. Current advances in the biochemical and physiological aspects of the treatment of type 2 diabetes mellitus with thiazolidinediones. PPAR Res. 2016, 2016, 7614270. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, R.F. Sodium-glucose cotransporter-2 inhibitors and genital and urinary tract infections in type 2 diabetes. Postgrad. Med. 2016, 128, 409–417. [Google Scholar] [PubMed]
- Cohen, F.J.; Neslusan, C.A.; Conklin, J.E.; Song, X. Recent antihyperglycemic prescribing trends for US privately insured patients with type 2 diabetes. Diabetes Care 2003, 26, 1847–1851. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.R.; Shrank, W.H.; Fischer, M.A.; Avorn, J.; Liberman, J.N.; Schneeweiss, S.; Pakes, J.; Brennan, T.A.; Choudhry, N.K. Patterns of medication initiation in newly diagnosed diabetes mellitus: Quality and cost implications. Am. J. Med. 2012, 125, 302.e1–302.e7. [Google Scholar] [CrossRef] [PubMed]
- Filipova, E.; Uzunova, K.; Kalinov, K.; Vekov, T. Pioglitazone and the risk of bladder cancer: A meta-analysis. Diabetes Ther. 2017, 8, 705–726. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.; Ojcius, D.M.; Chang, C.J.; Lin, C.S.; Lu, C.C.; Ko, Y.F.; Tseng, S.F.; Lai, H.C.; Young, J.D. Anti-obesogenic and antidiabetic effects of plants and mushrooms. Nat. Rev. Endocrinol. 2017, 13, 149–160. [Google Scholar] [PubMed]
- Muller, U.; Stubl, F.; Schwarzinger, B.; Sandner, G.; Iken, M.; Himmelsbach, M.; Schwarzinger, C.; Ollinger, N.; Stadlbauer, V.; Hoglinger, O.; et al. In vitro and in vivo inhibition of intestinal glucose transport by guava (Psidium guajava) extracts. Mol. Nutr. Food Res. 2018, 62, 1701012. [Google Scholar] [CrossRef] [PubMed]
- Lanzerstorfer, P.; Stadlbauer, V.; Chtcheglova, L.A.; Haselgrubler, R.; Borgmann, D.; Wruss, J.; Hinterdorfer, P.; Schroder, K.; Winkler, S.M.; Hoglinger, O.; et al. Identification of novel insulin mimetic drugs by quantitative total internal reflection fluorescence (TIRF) microscopy. Br. J. Pharmacol. 2014, 171, 5237–5251. [Google Scholar] [PubMed] [Green Version]
- Stadlbauer, V.; Haselgrubler, R.; Lanzerstorfer, P.; Plochberger, B.; Borgmann, D.; Jacak, J.; Winkler, S.M.; Schroder, K.; Hoglinger, O.; Weghuber, J. Biomolecular characterization of putative antidiabetic herbal extracts. PLoS ONE 2016, 11, e0148109. [Google Scholar] [CrossRef] [PubMed]
- Pessin, J.E.; Saltiel, A.R. Signaling pathways in insulin action: Molecular targets of insulin resistance. J. Clin. Investig. 2000, 106, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.J.; Govers, R.; James, D.E. Regulated transport of the glucose transporter GLUT4. Nat. Rev. Mol. Cell Biol. 2002, 3, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Minakawa, M.; Miura, Y.; Yagasaki, K. Piceatannol, a resveratrol derivative, promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and suppresses blood glucose levels in type 2 diabetic model db/db mice. Biochem. Biophys. Res. Commun. 2012, 422, 469–475. [Google Scholar] [PubMed]
- Prasad, C.N.; Anjana, T.; Banerji, A.; Gopalakrishnapillai, A. Gallic acid induces GLUT4 translocation and glucose uptake activity in 3T3-L1 cells. FEBS Lett. 2010, 584, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Haselgrubler, R.; Stubl, F.; Essl, K.; Iken, M.; Schroder, K.; Weghuber, J. Gluc-HET, a complementary chick embryo model for the characterization of antidiabetic compounds. PLoS ONE 2017, 12, e0182788. [Google Scholar] [CrossRef] [PubMed]
- Haselgrubler, R.; Stubl, F.; Stadlbauer, V.; Lanzerstorfer, P.; Weghuber, J. An in ovo model for testing insulin-mimetic compounds. J. Vis. Exp. JoVE 2018, 134, 57237. [Google Scholar] [CrossRef] [PubMed]
- Spielmann, H. HET-CAM test. Methods Mol. Biol. 1995, 43, 199–204. [Google Scholar] [PubMed]
- Yoshiyama, Y.; Sugiyama, T.; Kanke, M. Experimental diabetes model in chick embryos treated with streptozotocin. Biol. Pharm. Bull. 2005, 28, 1986–1988. [Google Scholar] [CrossRef] [PubMed]
- Onur, S.; Stöckmann, H.; Zenthoefer, M.; Piker, L.; Döring, F. The plant extract collection kiel in schleswig-holstein (peckish) is an open access screening library. J. Food Res. 2013, 2, 101–106. [Google Scholar] [CrossRef]
- Lanzerstorfer, P.; Borgmann, D.; Schutz, G.; Winkler, S.M.; Hoglinger, O.; Weghuber, J. Quantification and kinetic analysis of Grb2-EGFR interaction on micro-patterned surfaces for the characterization of EGFR-modulating substances. PLoS ONE 2014, 9, e92151. [Google Scholar] [CrossRef] [PubMed]
- Lanzerstorfer, P.; Yoneyama, Y.; Hakuno, F.; Muller, U.; Hoglinger, O.; Takahashi, S.; Weghuber, J. Analysis of insulin receptor substrate signaling dynamics on microstructured surfaces. FEBS J. 2015, 282, 987–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Kim, J.K.; Li, Y.; Li, J.; Liu, F.; Chen, X. Tannic acid stimulates glucose transport and inhibits adipocyte differentiation in 3T3-L1 cells. J. Nutr. 2005, 135, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Bruzzone, S.; Ameri, P.; Briatore, L.; Mannino, E.; Basile, G.; Andraghetti, G.; Grozio, A.; Magnone, M.; Guida, L.; Scarfi, S.; et al. The plant hormone abscisic acid increases in human plasma after hyperglycemia and stimulates glucose consumption by adipocytes and myoblasts. FASEB J. 2012, 26, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Eid, H.M.; Thong, F.; Nachar, A.; Haddad, P.S. Caffeic acid methyl and ethyl esters exert potential antidiabetic effects on glucose and lipid metabolism in cultured murine insulin-sensitive cells through mechanisms implicating activation of AMPK. Pharm. Biol. 2017, 55, 2026–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Hu, J.; Zhao, W.; Gao, X.; Jiang, C.; Liu, K.; Liu, B.; Huang, F. Quercetin differently regulates insulin-mediated glucose transporter 4 translocation under basal and inflammatory conditions in adipocytes. Mol. Nutr. Food Res. 2014, 58, 931–941. [Google Scholar] [CrossRef] [PubMed]
- NCD Risk Factor Collaboration. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4.4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef]
- Iqbal, Z.; Azmi, S.; Yadav, R.; Ferdousi, M.; Kumar, M.; Cuthbertson, D.J.; Lim, J.; Malik, R.A.; Alam, U. Diabetic Peripheral Neuropathy: Epidemiology, Diagnosis, and Pharmacotherapy. Clin. Ther. 2018, 40, 828–849. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Fonseca, V. From guideline to patient: A review of recent recommendations for pharmacotherapy of painful diabetic neuropathy. J. Diabetes Complic. 2015, 29, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Refaat, M.; Mohammedi, K.; Jayyousi, A.; Al Suwaidi, J.; Abi Khalil, C. Macrovascular Complications in Patients with Diabetes and Prediabetes. BioMed Res. Int. 2017, 2017, 7839101. [Google Scholar] [CrossRef] [PubMed]
- Meunier, V.; Bourrie, M.; Berger, Y.; Fabre, G. The human intestinal epithelial cell line Caco-2; pharmacological and pharmacokinetic applications. Cell Biol. Toxicol. 1995, 11, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Volpe, D.A. Drug-permeability and transporter assays in Caco-2 and MDCK cell lines. Future Med. Chem. 2011, 3, 2063–2077. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M. Facilitative glucose transporters. Eur. J. Biochem. 1994, 219, 713–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueckler, M. Insulin resistance and the disruption of Glut4 trafficking in skeletal muscle. J. Clin. Investig. 2001, 107, 1211–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayem, A.S.M.; Arya, A.; Karimian, H.; Krishnasamy, N.; Ashok Hasamnis, A.; Hossain, C.F. Action of phytochemicals on insulin signaling pathways accelerating glucose transporter (glut4) protein translocation. Molecules 2018, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Deng, S.; Han, Q.; Zhao, P.; Zhou, Q.; Zheng, S.; Ma, X.; Xu, C.; Yang, J.; Yang, X. Hypoglycemic Activity and the Potential Mechanism of the Flavonoid Rich Extract from Sophora tonkinensis Gagnep. in KK-Ay Mice. Front. Pharmacol. 2016, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Kadan, S.; Sasson, Y.; Saad, B.; Zaid, H. Gundelia tournefortii Antidiabetic Efficacy: Chemical Composition and GLUT4 Translocation. Evid. Complement. Altern. Med. 2018, 2018, 8294320. [Google Scholar] [CrossRef] [PubMed]
- Malematja, R.O.; Bagla, V.P.; Njanje, I.; Mbazima, V.; Poopedi, K.W.; Mampuru, L.; Mokgotho, M.P. Potential Hypoglycaemic and Antiobesity Effects of Senna italica Leaf Acetone Extract. Evid. Complement. Altern. Med. 2018, 2018, 5101656. [Google Scholar] [CrossRef] [PubMed]
- Naowaboot, J.; Pannangpetch, P.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U.; Itharat, A. Mulberry leaf extract stimulates glucose uptake and GLUT4 translocation in rat adipocytes. Am. J. Chin. Med. 2012, 40, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H.; Tsuru, R.; Li, L.; Zhou, J.; Kudo, M.; Liu, T.; Gao, M. Cyclocarya paliurus extract activates insulin signaling via Sirtuin1 in C2C12 myotubes and decreases blood glucose level in mice with impaired insulin secretion. PLoS ONE 2017, 12, e0183988. [Google Scholar] [CrossRef] [PubMed]
- Mutlur Krishnamoorthy, R.; Carani Venkatraman, A. Polyphenols activate energy sensing network in insulin resistant models. Chem. Biol. Int. 2017, 275, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Rozanska, D.; Regulska-Ilow, B. The significance of anthocyanins in the prevention and treatment of type 2 diabetes. Adv. Clin. Exp. Med. 2018, 27, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torabi, S.; DiMarco, N.M. Original Research: Polyphenols extracted from grape powder induce lipogenesis and glucose uptake during differentiation of murine preadipocytes. Exp. Biol. Med. 2016, 241, 1776–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa Marques, T.H.; Santos De Melo, C.H.; Fonseca De Carvalho, R.B.; Costa, L.M.; De Souza, A.A.; David, J.M.; De Lima David, J.P.; De Freitas, R.M. Phytochemical profile and qualification of biological activity of an isolated fraction of Bellis perennis. Biol. Res. 2013, 46, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Karakas, F.P.; Cingoz, G.S.; Turker, A.U. The effects of oxidative stress on phenolic composition and antioxidant metabolism in callus culture of common daisy. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Nazaruk, J.; Gudej, J. Apigenin glycosides from the flowers of Bellis perennis L. Acta Pol. Pharm. 2000, 57, 129–130. [Google Scholar] [PubMed]
- Nazaruk, J.; Gudej, J. Qualitative and quantitative chromatographic investigation of flavonoids in Bellis perennis L. Acta Pol. Pharm. 2001, 58, 401–404. [Google Scholar] [PubMed]
- Siatka, T.; Kasparova, M. Seasonal variation in total phenolic and flavonoid contents and DPPH scavenging activity of Bellis perennis L. flowers. Molecules 2010, 15, 9450–9461. [Google Scholar] [CrossRef] [PubMed]
- Mitich, L. English Daisy (Bellis perennis L.). Weed Technol. 1997, 11, 626–628. [Google Scholar] [CrossRef]
- Cakilcioglu, U.; Turkoglu, I. An ethnobotanical survey of medicinal plants in Sivrice (Elazig-Turkey). J. Ethnopharmacol. 2010, 132, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Uysal, I.; Onar, S.; Karabacak, E.; Celik, S. Ethnobotanical aspects of Kapidag Peninsula (Turkey). Biodivers. Conserv. 2010, 3, 15–22. [Google Scholar]
- Uzun, E.; Sariyar, G.; Adsersen, A.; Karakoc, B.; Otuk, G.; Oktayoglu, E.; Pirildar, S. Traditional medicine in Sakarya province (Turkey) and antimicrobial activities of selected species. J. Ethnopharmacol. 2004, 95, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Li, X.; Nishida, E.; Ito, Y.; Matsuda, H.; Nakamura, S.; Muraoka, O.; Yoshikawa, M. Perennisosides I–VII, acylated triterpene saponins with antihyperlipidemic activities from the flowers of Bellis perennis. J. Nat. Prod. 2008, 71, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Li, X.; Nishida, E.; Nakamura, S.; Ninomiya, K.; Matsuda, H.; Hamao, M.; Muraoka, O.; Hayakawa, T.; Yoshikawa, M. Medicinal Flowers. XXXII. Structures of oleanane-type triterpene saponins, perennisosides VIII, IX, X, XI, and XII, from the flowers of Bellis perennis. Chem. Pharm. Bull. 2011, 59, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Ninomiya, K.; Takamori, Y.; Nishida, E.; Yasue, M.; Hayakawa, T.; Muraoka, O.; Li, X.; Nakamura, S.; Yoshikawa, M.; et al. Oleanane-type triterpene saponins with collagen synthesis-promoting activity from the flowers of Bellis perennis. Phytochemistry 2015, 116, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Li, X.; Nishida, E.; Nakamura, S.; Matsuda, H.; Muraoka, O.; Morikawa, T. Medicinal flowers. XXI. Structures of perennisaponins A, B, C, D, E, and F, acylated oleanane-type triterpene oligoglycosides, from the flowers of Bellis perennis. Chem. Pharm. Bull. 2008, 56, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Schopke, T.; Wray, V.; Kunath, A.; Hiller, K. Bayogenin and asterogenic acid glycosides from Bellis perennis. Phytochemistry 1992, 31, 2555–2557. [Google Scholar] [PubMed]
Sample Availability: Samples of the bellis perennis extracts are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
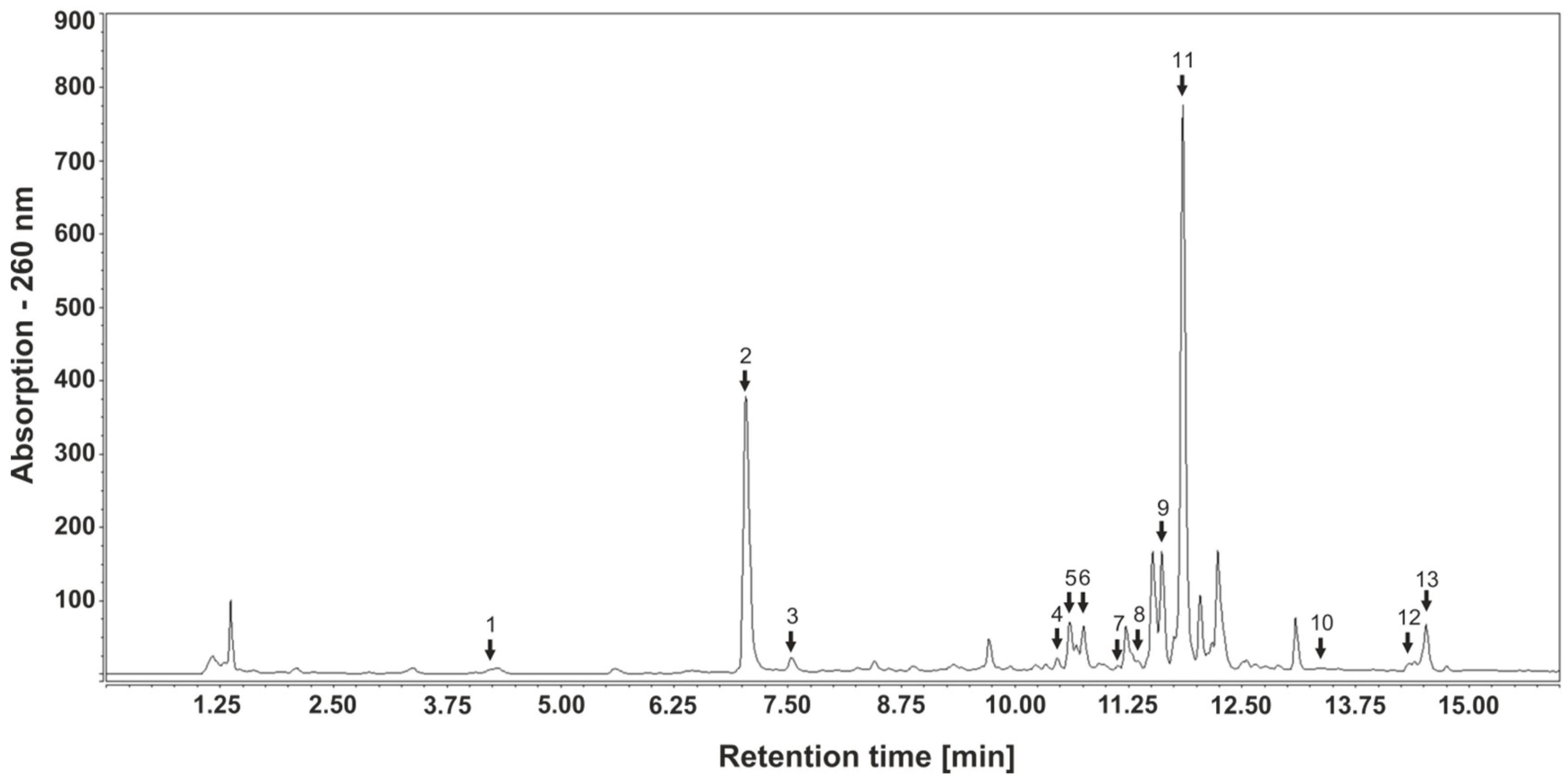{kind=link}
| Peak | Retention Time, tR [min] | Compound | Mass Spectrometry | Concentration | Concentration | Concentration |
|---|---|---|---|---|---|---|
| Number | (M-H)- | [mg/mL] | [mg/mL] | [mg/mL] | ||
| [m/z] | Home-Made Extract | Extract 4404 | Extract 4407 | |||
| Hydroxycinnamic acids | ||||||
| 1 | 4.22 | Neochlorogenic acid | 353.087 | 0.0261 | 0.0053 | 0.0094 |
| 2 | 7.04 | Chlorogenic acid | 353.087 | 1.6904 | 0.1605 | 0.2280 |
| 3 | 7.50 | Caffeic acid | 179.0352 | 0.0302 | 0.0059 | 0.0090 |
| Flavonols | ||||||
| 4 | 10.46 | Rutin | 609.1454 | 0.021 | 0.0040 | 0.0026 |
| 5 | 10.6 | Hyperoside | 463.088 | 0.0431 | 0.0020 | 0.0035 |
| 6 | 10.68 | Isoquercitrin | 464.0961 | 0.0567 | 0.0016 | 0.0111 |
| 7 | 11.22 | Guaijaverin | 433.0776 | 0.0456 | 0.0023 | 0.0048 |
| 8 | 11.4 | Avicularin | 433.0774 | 0.0115 | 0.0011 | 0.0038 |
| 9 | 11.67 | Quercitrin | 447.0933 | 0.1036 | 0.0297 | 0.0170 |
| 10 | 13.5 | Quercetin | 302.0433 | 0.0022 | 0.0002 | l.o.d. |
| 13 | 14.59 and 14.68 | Kaempferol and Luteolin | 285.0403 | n.q. | n.q. | n.q. |
| Flavones | ||||||
| 11 | 11.96 and 12.00 | Apigenin-7-glucoside and Apigenin-7-glucuronide | 431.0982 | 0.423 | n.q. | n.q. |
| 12 | 14.34 | Apigenin | 269.0444 | 0.0055 | 0.0001 | 0.0040 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haselgrübler, R.; Stadlbauer, V.; Stübl, F.; Schwarzinger, B.; Rudzionyte, I.; Himmelsbach, M.; Iken, M.; Weghuber, J. Insulin Mimetic Properties of Extracts Prepared from Bellis perennis. Molecules 2018, 23, 2605. https://doi.org/10.3390/molecules23102605
Haselgrübler R, Stadlbauer V, Stübl F, Schwarzinger B, Rudzionyte I, Himmelsbach M, Iken M, Weghuber J. Insulin Mimetic Properties of Extracts Prepared from Bellis perennis. Molecules. 2018; 23(10):2605. https://doi.org/10.3390/molecules23102605
Chicago/Turabian StyleHaselgrübler, Renate, Verena Stadlbauer, Flora Stübl, Bettina Schwarzinger, Ieva Rudzionyte, Markus Himmelsbach, Marcus Iken, and Julian Weghuber. 2018. "Insulin Mimetic Properties of Extracts Prepared from Bellis perennis" Molecules 23, no. 10: 2605. https://doi.org/10.3390/molecules23102605