Energy-Protein Supplementation and Lactation Affect Fatty Acid Profile of Liver and Adipose Tissue of Dairy Cows


Abstract
:1. Introduction
2. Results
2.1. The Effect of Stage of Lactation and Parity on Fatty Acid Profiles
2.1.1. Blood
2.1.2. Liver
2.1.3. Adipose Tissue
2.2. The Effect of Energy-Protein Supplementation on Fatty Acid Profiles
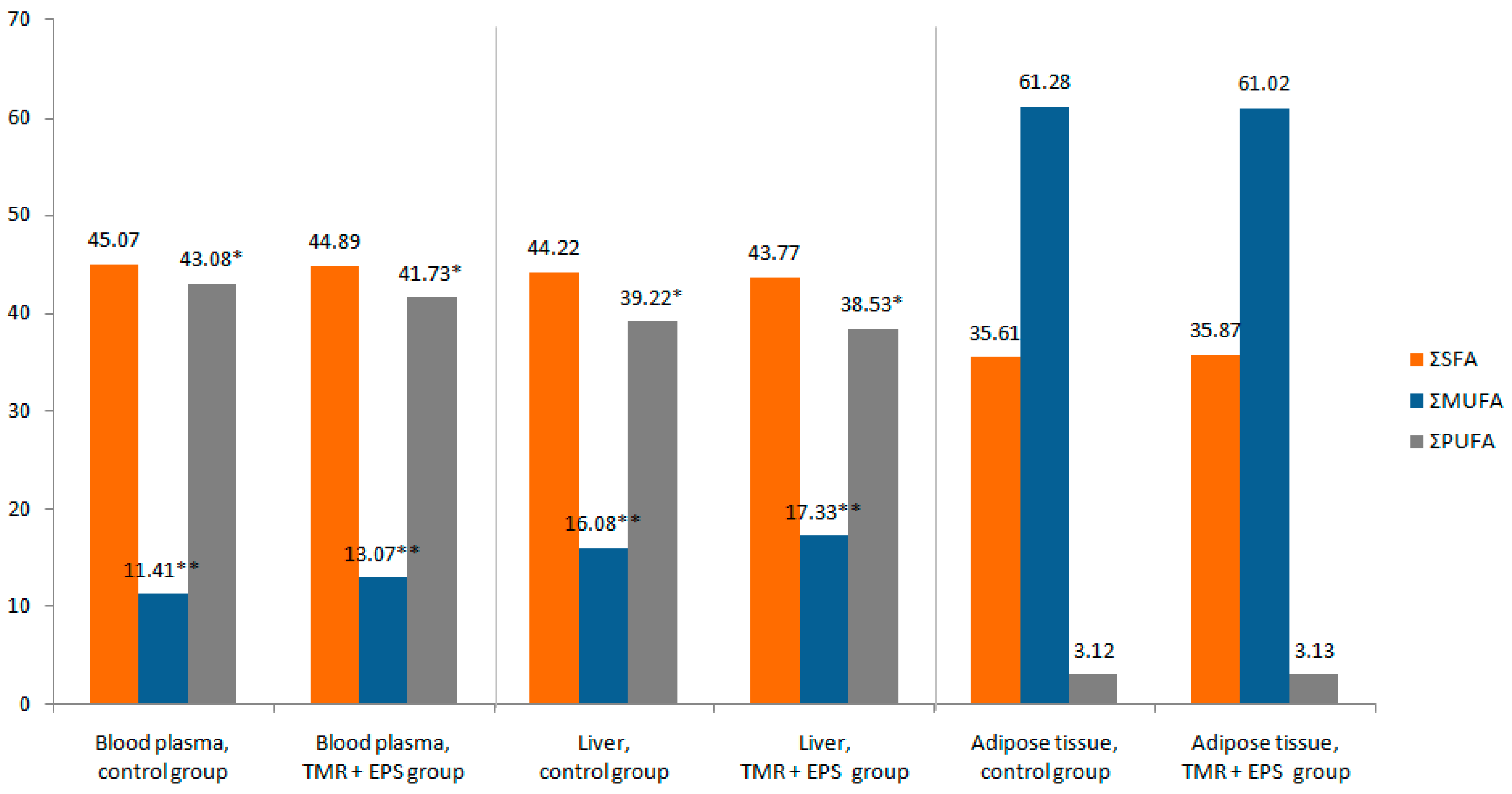
2.2.1. Blood
2.2.2. Liver
2.2.3. Adipose Tissue
3. Discussion
3.1. The Effect of Parity and Stage of Lactation on Fatty Acid Profiles
3.2. The Effect of Energy-Protein Supplementation on Fatty Acid Profiles
3.2.1. Blood
3.2.2. Liver
3.2.3. Adipose Tissue
4. Materials and Methods
4.1. Animals, Treatments and Experimental Design
4.2. Diets and Feeding Additive
4.3. Tissue Sampling
4.4. Lipid Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lock, A.L.; Bauman, D.E. Modifying milk fat composition of dairy cows to enhance fatty acids beneficial to human health. Lipids 2004, 39, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Bilby, T.R.; Block, J.; do Amaral, B.C.; Sa Filho, O.; Silvestre, F.T.; Hansen, P.J.; Staples, C.R.; Thatcher, W.W. Effects of dietary unsaturated fatty acids on oocyte quality and follicular development in lactating dairy cows in summer. J. Dairy Sci. 2006, 89, 3891–3903. [Google Scholar] [CrossRef]
- Santos, J.E.P.; Bilby, T.R.; Thatcher, W.W.; Staples, C.R.; Silvestre, F.T. Long chain fatty acids of diet as factors influencing reproduction in cattle. Reprod. Domest. Anim. 2008, 43 (Suppl. S2), 23–30. [Google Scholar] [CrossRef] [PubMed]
- Garnsworthy, P.C.; Lock, A.; Mann, G.E.; Sinclair, K.D.; Webb, R. Nutrition, metabolism, and fertility in dairy cows: 2. Dietary fatty acids and ovarian function. J. Dairy Sci. 2008, 91, 3824–3833. [Google Scholar] [CrossRef] [PubMed]
- Abayasekara, D.R.E.; Wathes, D.C. Effects of altering dietary fatty acid composition on prostaglandin synthesis and fertility. Prostaglandins Leukot. Essent. Fatty Acids 1999, 61, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Camacho, J.; Soberano-Martinez, A.; Duran, K.E.O.; Aguilar-Perez, C.; Ku-Vera, J.C. Effect of fatty acids on reproductive performance of ruminants. In Artificial Insemination in Farm Animals; Manafi, M., Ed.; InTech: Rijeka, Croatia, 2011; ISBN 978-953-307-312-5. [Google Scholar]
- Samková, E.; Špička, J.; Pešek, M.; Pelikánová, T.; Hanuš, O. Animal factors affecting fatty acid composition of cow milk fat: A review. S. Afr. J. Anim. Sci. 2012, 42, 83–100. [Google Scholar]
- Brzozowska, A.M.; Oprzadek, J.M. Metabolism of fatty acids in tissues and organs of the ruminants—A review. Anim. Sci. Pap. Rep. 2016, 34, 211–220. [Google Scholar]
- Mansbridge, R.J.; Blake, J.S. Nutritional factors affecting the fatty acid composition of bovine milk. Br. J. Nutr. 1997, 78, S37–S47. [Google Scholar] [CrossRef] [PubMed]
- Stoop, W.M.; Bovenhuis, H.; Heck, J.M.L.; van Arendonk, J.A.M. Effect of lactation stage and energy status on milk fat composition of Holstein-Friesian cows. J. Dairy Sci. 2009, 92, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Harfoot, C.G.; Hazlewood, G.P. Lipid metabolism in the rumen. In Rumen Microbial Ecosystem; Hobson, P.N., Ed.; Elsevier Applied Science Publication: London, UK, 1988; pp. 285–322. ISBN 1851661883. [Google Scholar]
- Kemp, P.; Lander, D.J. Hydrogenation in vitro of α-linolenic acid to stearic acid by mixed cultures of pure strains of rumen bacteria. J. Gen. Microbiol. 1984, 130, 527–533. [Google Scholar] [CrossRef]
- Mashek, D.G.; Bertics, S.J.; Grummer, R.R. Effects of intravenous triacylglycerol emulsions on hepatic metabolism and blood metabolites in fasted dairy cows. J. Dairy Sci. 2005, 88, 100–109. [Google Scholar] [CrossRef]
- Brzozowska, A.M.; Micek, P.; Kuczynska, B.; Lukaszewicz, M.; Oprzadek, J. The use of energy-protein supplement increases performance of high-yielding dairy cows and improves health-promoting properties of milk. Anim. Prod. Sci. 2017. [Google Scholar] [CrossRef]
- Kowalski, Z.M. Rumen fermentation, nutrient flow to the duodenum and digestibility in bulls fed calcium soaps of rapeseed fatty acids and soya bean meal coated with calcium soaps. Anim. Feed Sci. Technol. 1997, 69, 289–303. [Google Scholar] [CrossRef]
- Cook, L.J.; Scott, T.W.; Faichney, G.J.; Davies, H.L. Fatty acid interrelationships in plasma, liver, muscle, and adipose tissues of cattle fed safflower oil protected from ruminal hydrogenation. Lipids 1972, 7, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.N.; Rehage, J.; Beaulieu, A.D.; Bahaa, A.O.; Drackley, J.K. Prepartum nutrition alters fatty acid composition in plasma, adipose tissue, and liver lipids of periparturient dairy cows. J. Dairy Sci. 2007, 90, 2941–2959. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Gutiérrez, E.; de Veth, M.J.; Lock, A.L.; Dwyer, D.A.; Murphy, K.D.; Bauman, D.E. Effect of supplementation with calcium salts of fish oil on n-3 fatty acids in milk fat. J. Dairy Sci. 2007, 90, 4149–4156. [Google Scholar] [CrossRef] [PubMed]
- Emery, R.S.; Liesman, J.S.; Herdt, T.H. Metabolism of long chain fatty acids by ruminant liver. J. Nutr. 1992, 122, 832–837. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Griinari, J.M. Role of biohydrogenation intermediates in milk fat depression. Eur. J. Lipid Sci. Technol. 2007, 109, 799–816. [Google Scholar] [CrossRef]
- Petit, H.V.; Palin, M.F.; Doepel, L. Hepatic lipid metabolism in transition dairy cows fed flaxseed. J. Dairy Sci. 2007, 90, 4780–4792. [Google Scholar] [CrossRef] [PubMed]
- Ballou, M.A.; Gomes, R.C.; Juchem, S.O.; DePeters, E.J. Effects of dietary supplemental fish oil during the peripartum period on blood metabolites and hepatic fatty acid compositions and total triacylglycerol concentrations of multiparous Holstein cows. J. Dairy Sci. 2009, 92, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, K.J.; Kramer, J.K.G.; Sauer, F.D.; Emmons, D.B. Influence of triglycerides and free fatty acids in milk replacers on calf performance, blood plasma, and adipose lipids. J. Dairy Sci. 1985, 68, 669–680. [Google Scholar] [CrossRef]
- Lake, S.L.; Weston, T.R.; Scholljegerdes, E.J.; Murrieta, C.M.; Alexander, B.M.; Rule, D.C.; Moss, G.E.; Hess, B.W. Effects of postpartum dietary fat and body condition score at parturition on plasma, adipose tissue, and milk fatty acid composition of lactating beef cows. J. Anim. Sci. 2007, 85, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, A.D.; Drackley, J.K.; Merchen, N.R. Concentrations of conjugated linoleic acid (cis-9, trans-11-octadecadienoic acid) are not increased in tissue lipids of cattle fed a high-concentrate diet supplemented with soybean oil. J. Anim. Sci. 2002, 80, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, H.S. A simple method for the isolation and purification of total lipids from animal tissue. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). Animal and Vegetable Fats and Oils—Preparation of Methyl Esters of Fatty Acids; ISO 5509:2000; ISO: Geneva, Switzerland, 2000. [Google Scholar]
Sample Availability: Samples of the energy-protein supplement compounds are not available from the authors. |


{kind=link}
| Component | Dry Matter |
|---|---|
| Forage:Concentrates (%) | 67:33 |
| Corn silage (kg) | 9.4 |
| Grass silage (kg) | 4.0 |
| Wheat straw (kg) | 1.8 |
| Silaged grains of corn (kg) | 0.7 |
| Dried molassed beet pulp (kg) | 1.8 |
| Concentrates a (kg) | 5.4 |
| Chemical Composition | TMR | TMR + EPS | Energy-Protein Supplement (EPS) |
|---|---|---|---|
| Crude ash | 92 | 89 | 79 |
| Protein | 140 | 144 | 188 |
| Crude fat | 27 | 48 | 319 |
| Crude fibre | 211 | 202 | 105 |
| Neutral detergent fibre | 430 | 412 | 271 |
| Starch | 161 | 150 | 12 |
| Fatty Acid (g/100 g) | TMR | TMR + EPS | Energy-Protein Supplement (EPS) |
|---|---|---|---|
| C12:0 | 0.28 | 0.16 | 0.12 |
| C14:0 | 0.62 | 0.87 | 1.32 |
| C14:1 c9 | 0.02 | 0.02 | 0.03 |
| C15:0 | 0.08 | 0.07 | 0.11 |
| C16:0 | 16.75 | 14.28 | 10.39 |
| C16:1 c9 | 0.73 | 0.96 | 1.59 |
| C18:0 | 2.54 | 2.97 | 3.99 |
| C18:1 c9 | 21.95 | 28.63 | 42.61 |
| C18:2 n-6 | 38.32 | 28.44 | 15.07 |
| C18:3 n-3 | 15.53 | 21.12 | 21.00 |
| C20:1 | 0.14 | 0.34 | 1.33 |
| C20:2 n-6 | 0.05 | 0.13 | 0.60 |
| C20:3 n-6 | 0.05 | 0.21 | 0.28 |
| C20:3 n-3 | 0.05 | 0.09 | 0.20 |
| C20:4 n-6 | 0.00 | 0.04 | 0.04 |
| C20:5 n-3 | 0.03 | 0.12 | 0.12 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brzozowska, A.M.; Lukaszewicz, M.; Oprzadek, J.M. Energy-Protein Supplementation and Lactation Affect Fatty Acid Profile of Liver and Adipose Tissue of Dairy Cows. Molecules 2018, 23, 618. https://doi.org/10.3390/molecules23030618
Brzozowska AM, Lukaszewicz M, Oprzadek JM. Energy-Protein Supplementation and Lactation Affect Fatty Acid Profile of Liver and Adipose Tissue of Dairy Cows. Molecules. 2018; 23(3):618. https://doi.org/10.3390/molecules23030618
Chicago/Turabian StyleBrzozowska, Anna M., Marek Lukaszewicz, and Jolanta M. Oprzadek. 2018. "Energy-Protein Supplementation and Lactation Affect Fatty Acid Profile of Liver and Adipose Tissue of Dairy Cows" Molecules 23, no. 3: 618. https://doi.org/10.3390/molecules23030618
APA StyleBrzozowska, A. M., Lukaszewicz, M., & Oprzadek, J. M. (2018). Energy-Protein Supplementation and Lactation Affect Fatty Acid Profile of Liver and Adipose Tissue of Dairy Cows. Molecules, 23(3), 618. https://doi.org/10.3390/molecules23030618