
Moonlighting Proteins in the Fuzzy Logic of Cellular Metabolism


1
Center for Biomolecular Sciences, College of Pharmacy, University of Illinois at Chicago, 900 South Ashland Avenue, Chicago, IL 60607, USA
2
Department of Biological Sciences, University of Illinois at Chicago, 900 South Ashland Avenue, Chicago, IL 60607, USA
*
Author to whom correspondence should be addressed.
Molecules 2020, 25(15), 3440; https://doi.org/10.3390/molecules25153440
Submission received: 30 May 2020
/
Revised: 9 July 2020
/
Accepted: 23 July 2020
/
Published: 29 July 2020
(This article belongs to the Special Issue The Fuzziness in Molecular, Supramolecular, and Systems Chemistry)
Abstract
:The numerous interconnected biochemical pathways that make up the metabolism of a living cell comprise a fuzzy logic system because of its high level of complexity and our inability to fully understand, predict, and model the many activities, how they interact, and their regulation. Each cell contains thousands of proteins with changing levels of expression, levels of activity, and patterns of interactions. Adding more layers of complexity is the number of proteins that have multiple functions. Moonlighting proteins include a wide variety of proteins where two or more functions are performed by one polypeptide chain. In this article, we discuss examples of proteins with variable functions that contribute to the fuzziness of cellular metabolism.
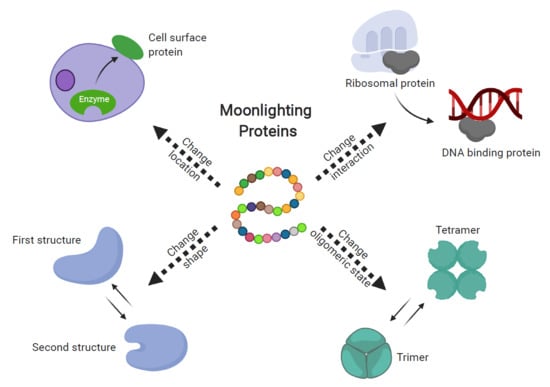
1. Introduction
Fuzzy logic systems include variables that can be any real number between 0 and 1 instead of being limited to the Boolean logic variables of only 0 and 1. This enables expression of complexity, uncertainty, and imprecision. In general, the vast interconnected biochemical pathways that make up the metabolism of a living cell can appear fuzzy because they are complex and hard to predict, and we have incomplete and not yet accurate knowledge. A single cell contains thousands of proteins performing a wide variety of activities, and the proteins have complex and constantly changing levels of expression, levels of activity, and patterns of interactions with other proteins and other molecules. Adding even more layers of complexity is the ability of many proteins, called moonlighting proteins, to perform more than one function. Moonlighting proteins are proteins in which one polypeptide chain performs more than one physiologically relevant biochemical or biophysical function [1,2,3] (Figure 1). The MoonProt Database (www.moonlightingproteins.org) contains annotations for over 300 experimentally confirmed moonlighting proteins, of which about 130 proteins are from human [4,5]. Although the mechanisms by which one protein performs two different functions are not always understood, it is clear that the function (or functions) performed at any specific time can be affected by multiple factors, and sometimes combinations of factors, including targeting to different cellular compartments, changes in the intracellular concentration of ligands, and changes in environmental conditions. In this paper, we describe examples of moonlighting proteins and some of the mechanisms by which they change function. These examples help illustrate and complement the ideas in this collection of papers on the topic of “The Fuzziness in Molecular Supramolecular, and Systems Chemistry”, where Gentili presents the “Fuzziness of the Molecular World” and describes natural information systems that involve fuzzy logic in large part due to proteins having multiple features and functions that vary in a context-dependent manner [6]. In addition, the paper by Fuxreiter describes using fuzzy set theory in a quantitative framework for describing the relationships between changing protein structures, interactions, and functions under changing, and somewhat unknown or unpredictable, cellular conditions [7].
2. Examples of Moonlighting Proteins and Factors that Affect Function
2.1. Cellular Localization
The most often observed subclass of moonlighting protein includes proteins that perform different functions in different cellular localizations. Over 100 enzymes and chaperones that catalyze reactions in the cytosol can be secreted and act as cytokines that modify the host’s immune system or become bound to the cell membrane where they serve as cell surface receptors, and in some cases these second functions contribute to virulence [8,9,10,11].
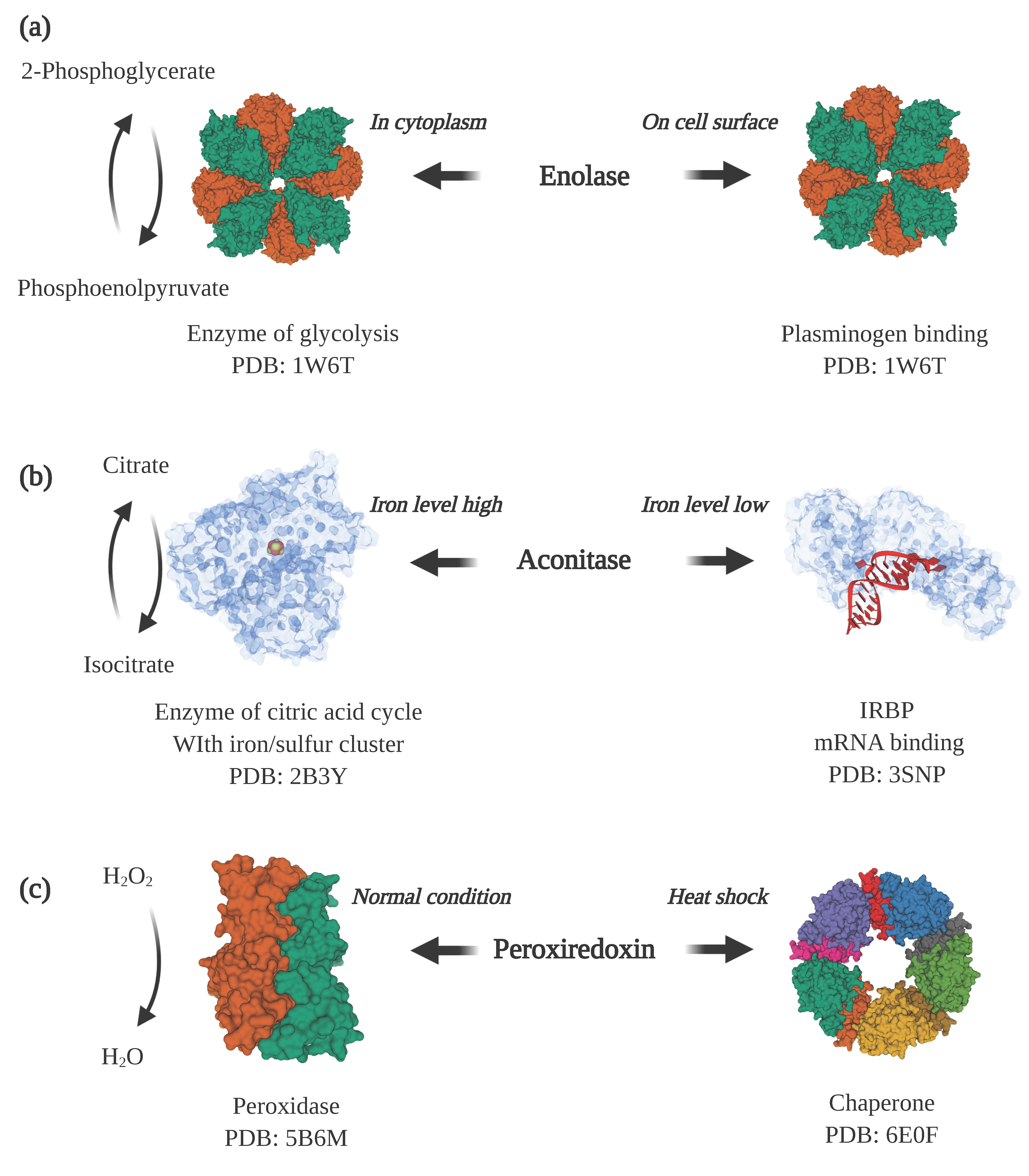
Enolase is one of these intracellular/surface moonlighting proteins in many species, including eukaryotes as well as prokaryotes. Inside the cell, it catalyzes the conversion of 2-phosphoglycerate to phosphoenolpyruvate in glycolysis. When displayed on the cell surface, it binds to host proteins (Figure 2a). The enolases from Aeromonas hydrophila, Bacillus anthracis, Neisseria meningitidis, Streptococcus pneumoniae, Trichomoniasis vaginalis and Lactobacillus crispatus can bind to host plasminogen [12,13,14,15,16,17]. The binding of plasminogen plays an important role in invasion of host tissues because, once bound to the cell surface receptor, the plasminogen becomes converted to the active protease, plasmin, which can aid in breaking down host extracellular matrix and invasion of tissues [18,19]. In some species, surface-located enolase and other intracellular/surface moonlighting proteins bind to other host proteins for colonization or for modulating the host immune system. Streptococcus suis enolase can also bind to host fibronectin, and Staphylococcus aureus enolase exhibits laminin binding activity [20,21].
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is another commonly found intracellular/surface moonlighting protein. It catalyzes the conversion of glyceraldehyde 3-phosphate to glycerate 1,3-bisphosphate in glycolysis in the cytoplasm. Some commensal bacteria that colonize the human gut use GAPDH on the cell surface to bind to host mucin and enable colonization of the gut [22,23]. When expressed on the cell surface, Streptococcus pyogenes GADPH can bind to plasminogen and fibronectin, and can also function as a ADP-ribosylating enzyme and assist with neutrophil evasion [24,25,26,27]. In addition, Streptococcus agalactiae GADPH can act as a modulator of the host’s immune system [28].
As the bacterial HSP70 (heat shock protein 70), DnaK is abundantly expressed in the cytosol as a stress-inducible chaperone. Mycobacterium tuberculosis DnaK can be displayed on the cell surface to bind to plasminogen, and it can also bind to CD40 (cluster of differentiation 40) to stimulate the synthesis of monocyte chemokines and the maturation of dendritic cells [29,30].
2.2. Interactions with Other Proteins and Molecules
Changes in cellular concentration of substrates or other ligands can serve as a trigger for changing protein functions. Aconitase is an enzyme in the citric acid cycle that contains an iron–sulfur cluster in the active site (Figure 2b). When the intracellular concentration of iron is high, the iron–sulfur cluster in the active site enables the isomerization of citrate to isocitrate [39]. In contrast, when the cellular level of iron is low, the iron–sulfur cluster is lost, and the enzyme changes conformation to expose an RNA binding surface and becomes an iron-responsive element binding protein (IRBP). As an IRBP, it binds to Iron Responsive Element (IRE) sequence motifs in mRNA, leading to increased translation of proteins involved in cellular iron uptake [40,41]. Another example is BirA, which performs functions as an enzyme and a transcription repressor in the biotin regulatory system [42]. The enzyme’s function is determined by the cellular need for biotin. When the need is high, when the cells are growing rapidly, BirA functions as a biotin–[acetyl–CoA-carboxylase] ligase that transfers a biotinyl moiety to the biotin carboxyl carrier protein subunit of acetyl–CoA carboxylase. When the demand decreases, under slower growth rates, BirA binds to DNA and functions as a biotin–operon repressor to inhibit the production of biotin [43].
As with aconitase and BirA, the different functions of many moonlighting proteins require interactions with different proteins, multiprotein complexes, DNA, RNA and other macromolecules. Many ribosomal proteins are moonlighting proteins, such as ribosomal proteins S3, S13, S14, L2, L4, L5, L7, L11, L13a, L23 and L26 [44,45,46,47,48,49,50,51,52,53,54]. These proteins have a second function when they dissociate from the ribosome and interact with other molecules. For example, the ribosomal protein S3 is a component of the 40S subunit of the ribosome, which is located in the cytoplasm. When S3 dissociates from the ribosome, it can enter the nucleus and act as a deoxyribonuclease (DNase) that cleaves apurinic/apyrimidinic sites during DNA repair [55]. Like the cytosolic aconitase mentioned above, mitochondrial aconitase acts as an enzyme in the citric acid cycle to catalyze the isomerization of citrate to isocitrate. However, instead of a function in translation, mitochondrial aconitase plays an important role in the maintenance of mitochondrial DNA that is independent of its catalytic role in the citric acid cycle [56].
2.3. Environmental Stress
Another common factor for moonlighting proteins to switch functions is environmental stresses. One example is peroxiredoxin, which in many species changes function from an enzyme to a protein-folding chaperone in response to oxidative stress or heat shock [57] (Figure 2c). In a lower molecular weight form, peroxiredoxin acts as a peroxidase that reduces hydrogen peroxide to water. However, under stress conditions, peroxiredoxin undergoes a shift to a higher molecular weight homo-oligomeric complex, comprised of five dimers connected by hydrophobic interactions [35]. The high molecular weight complex is a molecular chaperone that helps with folding and stabilizing proteins disrupted by the cell stress conditions.
Another example is the DegP protease, which also has a temperature-dependent change of function, where it transitions between a protease and a molecular chaperone. Under low temperature conditions, DegP functions as a molecular chaperone with the proteolytic site inactivated. When temperatures increase, the proteolytic site is activated and DegP can catalyze protein degradation [58].
2.4. Changes in Protein Structure
Changes in cellular localization, interaction partners, and environmental conditions can trigger a change in protein function with little or no change in protein structure. For example, in some moonlighting enzymes that are also transcription factors, ligand binding can turn on the transcription factor function by causing a relatively small change in overall structure that increases its DNA binding affinity [59]. Other moonlighting proteins undergo a large conformational change, such as cellular aconitase becoming IRBP, which involves a large relative movement of several domains within a protein subunit to uncover a previously buried mRNA binding surface. The peroxiredoxins described above undergo changes in quaternary structure to become chaperones.
In general, many moonlighting proteins undergo changes in structure, which can range from small movements of surface loops to the more drastic change in tertiary or quaternary structure observed in intrinsically disordered proteins (IDPs), metamorphic proteins, or morpheeins.
Intrinsically disordered proteins have a region or subunit that is unfolded and can in some cases enable a switch from unfolded to multiple folded structures that enable interactions with different proteins. Metamorphic proteins contain a domain or subunit that folds into more than one stable structure. Morpheein proteins have subunits that disassemble, change conformation, and reassemble into different quaternary structures. In some cases, these structural changes are involved in regulation of a single function, but in other cases, the structural changes are correlated with a switch between two different functions.
In this next section, we describe examples of IDPs, metamorphic proteins, and morpheeins and how variability in protein structure contributes to some of these proteins being moonlighting proteins with “fuzziness” in protein function.
2.4.1. Intrinsically Disordered Proteins
Intrinsically disordered proteins contain regions, sometimes the entire polypeptide chain, that are unfolded under physiological conditions. Some unfolded regions are fully functional without becoming completely folded, and others undergo reversible folding when binding with other molecules [60,61]. In some cases, this flexibility enables one IDP to bind to a variety of cellular components, including small molecules, other proteins, DNA, or RNA, which enables many proteins to perform more than one function and contributes to the complexity in cellular metabolism and regulation of transcription and translation, molecular translocation, DNA repair and replication, and cell signaling [62,63,64,65].
Thirteen proteins in the database of moonlighting proteins (MoonProt, moonlightingproteins.org) are also found in the database of intrinsically disordered proteins (DisProt.org) (Table 1) [5,66]. Two proteins are discussed further below, p53 and thymosin beta-4.
The tumor suppressor protein p53 is a moonlighting protein and also an IDP with roles in cell cycle regulation, DNA repair and apoptosis [67,68]. In normal cells, the levels of p53 are low, but if cells sense environmental dangers that cause DNA damage, such as toxins, viruses or radiation, the level of p53 rises. Then, p53 binds to regulatory elements in the genome, activating a cascade of cellular responses to stop cell division and prevent cells from uncontrolled growth. In DNA repair, p53 can interact directly with DNA polymerase and AP endonuclease to stimulate base excision repair [69]. Outside the nucleus, p53 also has several cytoplasmic functions, including in centrosome duplication, induction of apoptosis and inhibition of autophagy [68].
In the native state, p53 has both folded and unfolded domains. The folded core domain is a DNA binding domain that recognizes specific regulatory elements. The folded tetramerization domain at the center of the protein joins protein subunits together into a homo-tetramer [70]. The N-terminal transactivation domain that interacts with and activates transcription factors is intrinsically disordered [71]. The C-terminal domain and the linker regions in between domains are also intrinsically disordered and fairly flexible, which enables the protein to adjust its conformation upon binding to specific regulatory sites in the DNA [72]. The flexibility of the intrinsically disordered domains allows p53 to recognize and bind to a large number of regulatory elements, so that it can regulate transcription in many different sites of the genome.
Thymosin beta-4 (Tβ4) is another moonlighting protein that is also an IDP. Tβ4 mainly functions in sequestering G-actin (monomeric actin) to prevent it from polymerization [73,74]. Tβ4 also has multiple moonlighting functions that are involved in diverse cellular roles including enhancement of endothelial cell differentiation, stimulation of angiogenesis, tissue regeneration and inhibition of inflammatory responses [75,76,77,78].
The free form of Tβ4 is intrinsically disordered and predominantly unstructured in solution. However, upon binding with G-actin, Tβ4 becomes fully folded and structured, where an extended conformation in the central region and two helices at the N-, C-termini can be identified [79,80]. In addition, Tβ4 forms complexes with PINCH (Particularly Interesting New Cys His-containing protein), ILK (Integrin-Linked Kinase) and stabilin-2 (an endocytic receptor for hyaluronic acid) respectively, where weak, transient and structurally ambiguous protein–protein interactions take place [81,82,83].
Protein structures in which significant conformational heterogeneity, disorder or ambiguity remain after formation of the complex are referred to as “fuzzy interactions” or “fuzzy complexes” and the remaining flexibility or disorder can be important in the assembly or activity of the complexes [116,117,118]. Fuzzy interactions and complexes can enable interactions with alternative partners and sensitivity to post-translational modifications. Fuzzy interactions also play a large part in several types of supramolecular interactions including in intracellular lipid droplets, which are described in another paper in this collection by Uversky [119].
The fuzzy complexes formed between IDPs and their binding partners, as mentioned above, can be an important feature of IDPs in fulfilling their functional versatility. Moonlighting proteins GCN4, HMGB1, CFTR, and Ure2 are found to be part of fuzzy complexes in the Fuzzy Complex Database (http://protdyn-database.org) [120]. As an example, GCN4 is a transcription activator for several genes, and is also a ribonuclease [86]. As a transcription activator, GCN4 binds Gal11 (an activator) in a weak and low affinity mode with multiple conformations. This conformational ambiguity is a typical example of a fuzzy complex where no single binding conformational state has been identified [117].
2.4.2. Metamorphic Proteins
Metamorphic proteins add another layer of complexity to our understanding of protein structure and function [121,122]. In stark contrast to the dogma of one sequence, one structure, one function, metamorphic proteins have two or more folded structures as their native structures, and in some cases the different structures have different functions. Distinct from the intrinsically disordered proteins, where the native states are folded or unfolded, the native states of metamorphic proteins are both folded and structured. The interconversions between native structures are reversible, meaning that at equilibrium there is a balance between the native structures [123]. Although the fact of two native structures appears to be against the thermodynamic principles of protein folding, in which the native structure of a protein has the overall lowest free energy, it has been shown that the two native structures can have similar energies with a low activation barrier of refolding [124]. So far, a small number of metamorphic proteins have been discovered, including lymphotactin, RfaH, CLIC1, Mad2, KaiB, IscU, Selecase and HIV-1 reverse transcriptase, of which the first three are also moonlighting proteins [125,126,127,128,129,130,131,132].
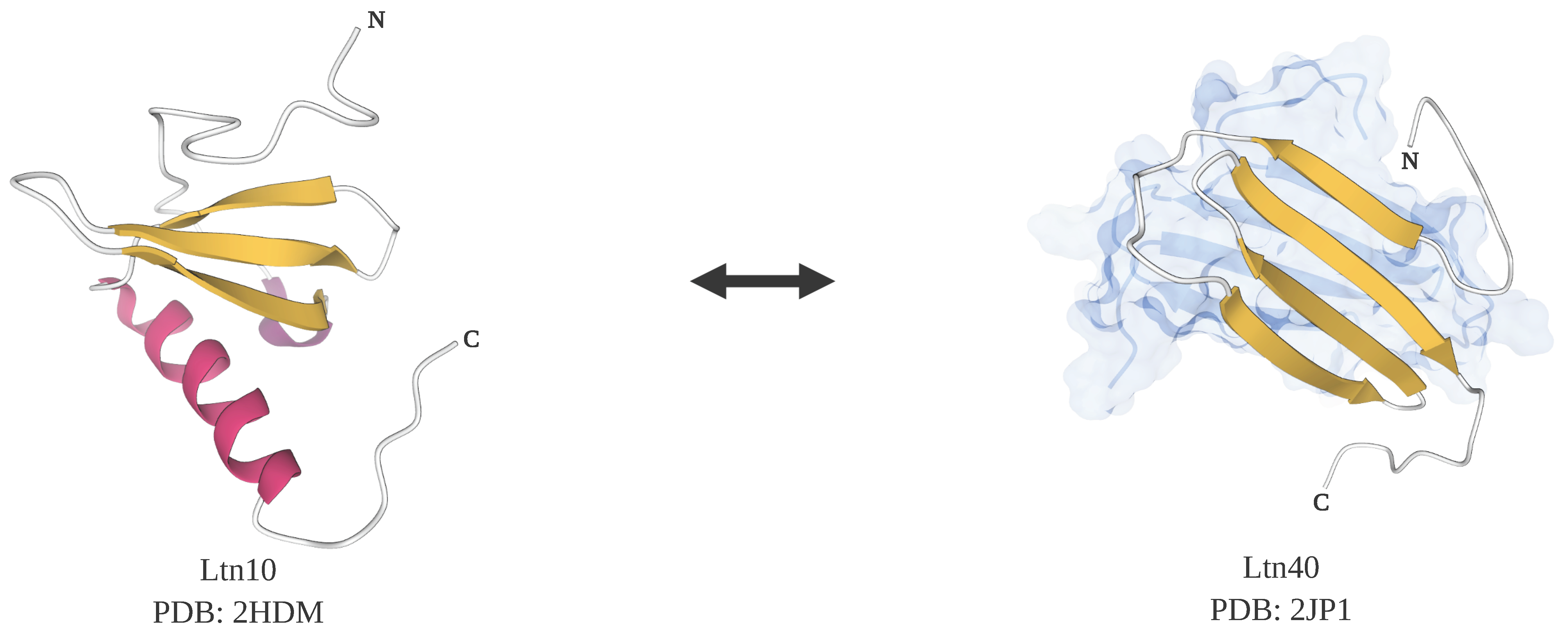
The C family chemokine lymphotactin (Ltn) is a metamorphic protein and also a moonlighting protein with heparin-binding activity [133]. Under normal physiological conditions, 37 °C and 150 mM NaCl, lymphotactin exists in an equilibrium between two native states, Ltn10 and Ltn40. Ltn10 is a monomer possessing a mix of beta sheet and alpha helix in a canonical chemokine fold that undergoes refolding and dimerization to become Ltn40, which contains a beta sandwich (Figure 3). While Ltn10 is an agonist for the X-C G-protein coupled chemokine receptor 1 (XCR1), Ltn40 can’t bind to XCR1 but instead can bind to heparin, a glycosaminoglycan component of the extracellular matrix. At equilibrium, there are nearly equal amounts of Ltn10 and Ltn40. The interconversions between Ltn10 and Ltn40 can be controlled by small changes in salt concentration and temperature. When the salt concentration is high and the temperature is low, the presence of Ltn10 is predominant, however, at lower salt concentrations and higher temperatures, but still below 40 °C, Ltn40 is the predominant species [125]. Most other chemokines don’t appear to undergo these transformations because they contain two disulfide bonds. Because lymphotactin only has one disulfide bond, it is less restricted and more flexible in changing conformations compared to other chemokines, which partially explains the reversibility between two distinct native structures. In experiments where an extra disulfide bond was introduced, lymphotactin could be locked in only one native state and the transition to the other state was prohibited, suggesting that a single amino acid modification can change the functionality of lymphotactin significantly [134].
Another metamorphic protein with moonlighting activity is RfaH, which functions as a transcription factor that inhibits termination and is also a translation factor. RfaH has two domains: a C-terminal domain (CTD) and an N-terminal domain (NTD). As a transcription factor, these two domains are tightly bound together, and the CTD is in an all-helical conformation that masks the RNAP (RNA polymerase) binding surface on the NTD, preventing the NTD from interacting with RNAP. When the NTD binds to specific operons in DNA, the two domains are separated, which enables the NTD to interact with RNAP. Meanwhile, the CTD undergoes a transformation from an all-helical conformation to an all-beta one, after which the CTD is able to recruit ribosomes and potentiates the translation of operons controlled by RfaH [126,135].
In addition, CLIC1 is a metamorphic protein with two native folded states and also a moonlighting protein with two different functions [136]. In an oligomeric form, CLIC1 functions as a transmembrane chloride ion channel with its N-terminus folded in an all-alpha native state. In a monomeric and soluble form, CLIC1 functions as an oxidoreductase with a transformed N-terminus that is a mixed structure containing an alpha helix and a beta sheet [137,138].
2.4.3. Morpheein Proteins
The subunits of morpheeins form a multimer that can disassemble, change conformation (without refolding), and reassemble into a different multimer [139].
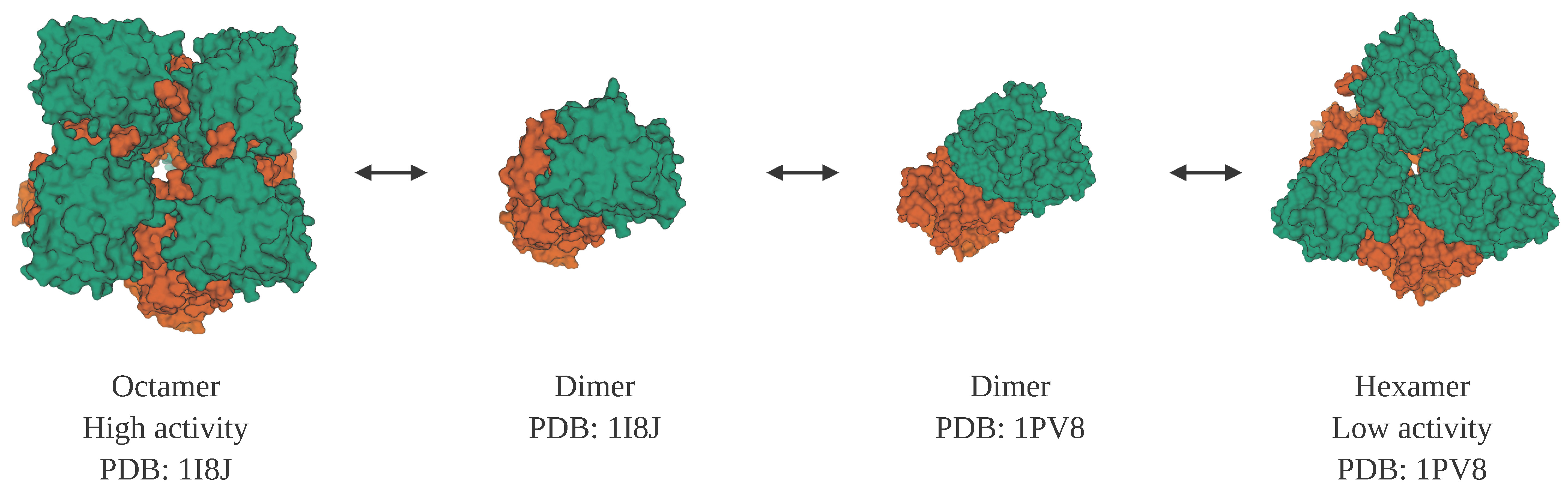
Porphobilinogen synthase (also known as delta-aminolevulinic acid dehydratase) is the prototype of a morpheein. It has two oligomeric states that correlate with different levels of enzyme activity and binding of allosteric effectors [140]. An octamer can disassemble into dimers. While part of a dimer, domains within the subunits can shift in their relative positions to result in subunits with a different conformation that can then assemble into a hexamer (Figure 4). While these different homomultimers vary in their level of activity of one function, porphobilinogen synthase enzyme activity, the protein is also a moonlighting protein because it has a second function in which it binds to and inhibits the proteasome [141,142].
The ebolavirus VP40 is a morpheein and moonlighting protein that has three different functions in the virus life cycle, and each of these functions corresponds to a different arrangement of subunits [145]. An octameric ring structure binds to viral RNA to regulate its transcription while in host cells. A butterfly-shaped dimer moves to the host cell’s plasma membrane. Then a linear hexameric form assembles into a larger structure that is needed for budding.
3. Moonlighting Proteins in Cellular Complexity
The variety of functions and combinations of functions of moonlighting proteins contribute to the complexity of cellular metabolism. Protein function, and in many cases the structure, is dependent on cellular factors that can vary due to intracellular conditions and the extracellular environment, and the output can change in a dynamic way. The examples given above are just some of the known factors affecting the functions of moonlighting proteins, and one protein often responds to multiple signals or combinations of signals. Some switches in function are reversible, others are not. Within a single cell, some copies of a moonlighting protein can be performing one function, some another, and some both simultaneously, depending on the protein, the cell type, and on the individual cell’s metabolic state and environmental conditions.
These factors that make our understanding of the cell difficult are valuable to the cell because they help enable dynamic responses to fluctuations in conditions within the cell and in its environment. Because the function of moonlighting proteins can depend on multiple factors, they can also be components of controllable cellular responses and be involved in processing information. Some moonlighting proteins also help regulate the level of activity of other proteins in the cell, for example by an enzyme with a second function in a cell signaling pathway or as translation or transcription factors. Because moonlighting proteins can have different activities in different cell types, they also contribute to different cell types having different phenotypes with specialized functions.
In addition to actually contributing to the complexity of the many interconnected pathways and processes in the cell, the ability of a protein to perform two different functions adds to the “fuzziness” of our inability to fully understand, predict, and model the activities in a cell and how they interact and are regulated. There are many things we still don’t know about moonlighting proteins. First, we don’t know how many proteins are moonlighting proteins. Many protein functions were found by serendipity and researchers are often looking for one type of function when they study a protein, not all of the functions that might be there. There are also many proteins identified through sequencing projects for which we don’t know any functions. We also don’t completely understand the triggers and mechanisms for switching between different functions—the cellular conditions, ligands, protein–protein interactions, conformational changes, PTMs, etc. involved and how all the different triggers combine.
Our understanding of moonlighting proteins and our ability to predict which proteins are moonlighting proteins and what are their functions is also complicated because a protein can have one second function, but a homologous protein can have a different second function, for example the cytoplasmic and mitochondrial aconitases mentioned above. Leucyl-tRNA synthetase is another example. It is an enzyme that attaches leucine to tRNA, but it has additional functions that vary in yeast and humans. In the yeast Saccharomyces cerevisiae, leucyl-tRNA synthetase is involved in intron splicing in RNA [146]. However, in humans it is involved in cell signaling, where it senses the cellular leucine concentration and binds to and activates Rag GTPase, leading to the activation of mTORC1 (mammalian target of rapamycin complex 1) [147].
In fact, a family of homologous enzymes can include enzymes with the canonical catalytic function of the protein family, moonlighting proteins with different combinations of catalytic and non-catalytic functions, as well as enzymes with variations on the canonical catalytic function (i.e., different substrates or chemical reaction catalyzed) and even pseudoenzymes, which can resemble active enzymes but have no catalytic function [148,149]. Though noncatalytic, pseudoenzymes play important roles in regulating the activity of their catalytic homologues, facilitating the assembly of scaffolding complexes and coordinating transcription and translation [150]. The loss of the catalytic functions can be attributed to a variety of aspects, such as the loss of essential amino acid residues needed for catalysis in the active site, a blockage of the entrance to the active site, and mutations of the amino acids involved in binding the substrate [151,152]. The first reported pseudoenzyme was alpha-lactalbumin, which is homologous to the enzyme lysozyme but does not have catalytic activity [153,154]. Instead, alpha-lactalbumin is a component of lactose synthase, serving as a regulatory subunit that increases the substrate binding affinity for the catalytic subunit of the enzyme [155]. Another example is found in the argininosuccinate lyase protein family. The canonical argininosuccinate lyase enzymes catalyze the breakdown of argininosuccinate into arginine and fumarate. Delta1-crystallin is a pseudoenzyme member of the family with a function as a structural protein in the lens of the eye in birds and reptiles. Another member of the protein family, delta2-crystallin, shares 94% amino acid sequence identity with the delta1-crystallin and is both a crystallin and a catalytically active argininosuccinate lyase that catalyzes the breakdown of argininosuccinate into arginine and fumarate [156,157].
4. Conclusions
The variability in the functions of moonlighting proteins, including some intrinsically disordered, metamorphic and morpheein proteins, contribute immensely to the fuzziness concept of cellular metabolism as described by Gentili [6]. Many types of proteins have multiple functions that interact in a complex pattern of interacting pathways and processes. As cellular conditions change due to metabolism and environmental conditions, the functions of these proteins change, resulting in different combinations of interactions and processes. The fuzziness concept also represents our limited understanding of these players and our inability to fully predict and model their actions and interactions. We don’t yet know how many proteins are moonlighting proteins or the full complement of their functions, and in many cases, we also don’t know the cellular factors that affect their functions. Moreover, while many advances have been made in our ability to predict protein functions, the variety of functions found even among homologous proteins adds to the fuzziness of our predictions.
Author Contributions
Writing—original draft preparation, H.L. and C.J.J.; writing—review and editing, H.L. and C.J.J. All authors have read and agreed to the published version of the manuscript.
Funding
Research on moonlighting proteins in the Jeffery Lab is supported by an award from the University of Illinois Cancer Center.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jeffery, C.J. Moonlighting proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 2003, 19, 415–417. [Google Scholar] [CrossRef]
- Jeffery, C.J. Protein moonlighting: What is it, and why is it important? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef] [PubMed]
- Mani, M.; Chen, C.; Amblee, V.; Liu, H.; Mathur, T.; Zwicke, G.; Zabad, S.; Patel, B.; Thakkar, J.; Jeffery, C.J. MoonProt: A database for proteins that are known to moonlight. Nucleic Acids Res. 2015, 43, D277–D282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zabad, S.; Liu, H.; Wang, W.; Jeffery, C. MoonProt 2.0: An expansion and update of the moonlighting proteins database. Nucleic Acids Res. 2018, 46, D640–D644. [Google Scholar] [CrossRef] [Green Version]
- Gentili, P.L. The Fuzziness of the Molecular World and Its Perspectives. Molecules 2018, 23, 2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxreiter, M. Towards a Stochastic Paradigm: From Fuzzy Ensembles to Cellular Functions. Molecules 2018, 23, 3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, B.; Martin, A. Bacterial virulence in the moonlight: Multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, C. Intracellular proteins moonlighting as bacterial adhesion factors. Aims Microbiol. 2018, 4, 362–376. [Google Scholar] [CrossRef]
- Wang, W.; Jeffery, C.J. An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol. Biosyst. 2016, 12, 1420–1431. [Google Scholar] [CrossRef]
- Kainulainen, V.; Korhonen, T.K. Dancing to another tune-adhesive moonlighting proteins in bacteria. Biology 2014, 3, 178–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, J.; Erova, T.E.; Alyea, R.A.; Wang, S.; Olano, J.P.; Pancholi, V.; Chopra, A.K. Surface-expressed enolase contributes to the pathogenesis of clinical isolate SSU of Aeromonas hydrophila. J. Bacteriol. 2009, 191, 3095–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, S.; Kulshreshtha, P.; Bambah Mukku, D.; Bhatnagar, R. α-Enolase binds to human plasminogen on the surface of Bacillus anthracis. Biochim. Biophys. Acta 2008, 1784, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Knaust, A.; Weber, M.V.; Hammerschmidt, S.; Bergmann, S.; Frosch, M.; Kurzai, O. Cytosolic proteins contribute to surface plasminogen recruitment of Neisseria meningitidis. J. Bacteriol. 2007, 189, 3246–3255. [Google Scholar] [CrossRef] [Green Version]
- Kolberg, J.; Aase, A.; Bergmann, S.; Herstad, T.K.; Rodal, G.; Frank, R.; Rohde, M.; Hammerschmidt, S. Streptococcus pneumoniae enolase is important for plasminogen binding despite low abundance of enolase protein on the bacterial cell surface. Microbiology 2006, 152, 1307–1317. [Google Scholar] [CrossRef] [Green Version]
- Mundodi, V.; Kucknoor, A.S.; Alderete, J.F. Immunogenic and plasminogen-binding surface-associated alpha-enolase of Trichomonas vaginalis. Infect. Immun. 2008, 76, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Antikainen, J.; Kuparinen, V.; Lahteenmaki, K.; Korhonen, T.K. Enolases from Gram-positive bacterial pathogens and commensal lactobacilli share functional similarity in virulence-associated traits. Fems Immunol. Med. Microbiol. 2007, 51, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Collen, D.; Verstraete, M. Molecular biology of human plasminogen. II. Metabolism in physiological and some pathological conditions in man. Thromb. Diath. Haemorrh. 1975, 34, 403–408. [Google Scholar]
- Dano, K.; Andreasen, P.A.; Grondahl-Hansen, J.; Kristensen, P.; Nielsen, L.S.; Skriver, L. Plasminogen activators, tissue degradation, and cancer. Adv. Cancer Res. 1985, 44, 139–266. [Google Scholar] [CrossRef]
- Esgleas, M.; Li, Y.; Hancock, M.A.; Harel, J.; Dubreuil, J.D.; Gottschalk, M. Isolation and characterization of alpha-enolase, a novel fibronectin-binding protein from Streptococcus suis. Microbiology 2008, 154, 2668–2679. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, C.R.; Postol, E.; Nomizo, R.; Reis, L.F.; Brentani, R.R. Identification of enolase as a laminin-binding protein on the surface of Staphylococcus aureus. Microbes Infect. 2004, 6, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Uchida, H.; Kawai, Y.; Kawasaki, T.; Wakahara, N.; Matsuo, H.; Watanabe, M.; Kitazawa, H.; Ohnuma, S.; Miura, K.; et al. Cell surface Lactobacillus plantarum LA 318 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) adheres to human colonic mucin. J. Appl. Microbiol. 2008, 104, 1667–1674. [Google Scholar] [CrossRef]
- Patel, D.K.; Shah, K.R.; Pappachan, A.; Gupta, S.; Singh, D.D. Cloning, expression and characterization of a mucin-binding GAPDH from Lactobacillus acidophilus. Int. J. Biol. Macromol. 2016, 91, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Lottenberg, R.; Broder, C.C.; Boyle, M.D.; Kain, S.J.; Schroeder, B.L.; Curtiss, R., 3rd. Cloning, sequence analysis, and expression in Escherichia coli of a streptococcal plasmin receptor. J. Bacteriol. 1992, 174, 5204–5210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pancholi, V.; Fischetti, V.A. A major surface protein on group a streptococci is a glyceraldehyde-3-phosphate-dehydrogenase with multiple binding activity. J. Exp. Med. 1992, 176, 415–426. [Google Scholar] [CrossRef]
- Pancholi, V.; Fischetti, V.A. Glyceraldehyde-3-phosphate dehydrogenase on the surface of group A streptococci is also an ADP-ribosylating enzyme. Proc. Natl. Acad. Sci. USA 1993, 90, 8154–8158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terao, Y.; Yamaguchi, M.; Hamada, S.; Kawabata, S. Multifunctional glyceraldehyde-3-phosphate dehydrogenase of Streptococcus pyogenes is essential for evasion from neutrophils. J. Biol. Chem. 2006, 281, 14215–14223. [Google Scholar] [CrossRef] [Green Version]
- Madureira, P.; Baptista, M.; Vieira, M.; Magalhaes, V.; Camelo, A.; Oliveira, L.; Ribeiro, A.; Tavares, D.; Trieu-Cuot, P.; Vilanova, M.; et al. Streptococcus agalactiae GAPDH is a virulence-associated immunomodulatory protein. J. Immunol. 2007, 178, 1379–1387. [Google Scholar] [CrossRef] [Green Version]
- Xolalpa, W.; Vallecillo, A.J.; Lara, M.; Mendoza-Hernandez, G.; Comini, M.; Spallek, R.; Singh, M.; Espitia, C. Identification of novel bacterial plasminogen-binding proteins in the human pathogen Mycobacterium tuberculosis. Proteomics 2007, 7, 3332–3341. [Google Scholar] [CrossRef]
- Wang, Y.; Kelly, C.G.; Karttunen, J.T.; Whittall, T.; Lehner, P.J.; Duncan, L.; MacAry, P.; Younson, J.S.; Singh, M.; Oehlmann, W.; et al. CD40 is a cellular receptor mediating mycobacterial heat shock protein 70 stimulation of CC-chemokines. Immunity 2001, 15, 971–983. [Google Scholar] [CrossRef] [Green Version]
- Ehinger, S.; Schubert, W.D.; Bergmann, S.; Hammerschmidt, S.; Heinz, D.W. Plasmin(ogen)-binding alpha-enolase from Streptococcus pneumoniae: Crystal structure and evaluation of plasmin(ogen)-binding sites. J. Mol. Biol. 2004, 343, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, J.; Volbeda, A.; Carpentier, P.; Darnault, C.; Moulis, J.M.; Fontecilla-Camps, J.C. Crystal structure of human iron regulatory protein 1 as cytosolic aconitase. Structure 2006, 14, 129–139. [Google Scholar] [CrossRef]
- Walden, W.E.; Selezneva, A.I.; Dupuy, J.; Volbeda, A.; Fontecilla-Camps, J.C.; Theil, E.C.; Volz, K. Structure of dual function iron regulatory protein 1 complexed with ferritin IRE-RNA. Science 2006, 314, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lee, W.; Kim, E.E. Crystal structures of human peroxiredoxin 6 in different oxidation states. Biochem. Biophys. Res. Commun. 2016, 477, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.; Tse, E.; Castro, H.; Makepeace, K.A.T.; Meinen, B.A.; Borchers, C.H.; Poole, L.B.; Bardwell, J.C.; Tomas, A.M.; Southworth, D.R.; et al. Chaperone activation and client binding of a 2-cysteine peroxiredoxin. Nat. Commun. 2019, 10, 659. [Google Scholar] [CrossRef] [PubMed]
- Sehnal, D.; Rose, A.S.; Kovca, J.; Burley, S.K.; Velankar, S. Mol*: Towards a common library and tools for web molecular graphics. In Workshop on Molecular Graphics and Visual Analysis of Molecular Data; Byška, J., Krone, M., Sommer, B., Eds.; The Eurographics Association: Aire-la-Ville, Switzerland, 2018; pp. 29–33. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.K.; Berman, H.M.; Bhikadiya, C.; Bi, C.; Chen, L.; Di Costanzo, L.; Christie, C.; Dalenberg, K.; Duarte, J.M.; Dutta, S.; et al. RCSB Protein Data Bank: Biological macromolecular structures enabling research and education in fundamental biology, biomedicine, biotechnology and energy. Nucleic Acids Res. 2019, 47, D464–D474. [Google Scholar] [CrossRef] [Green Version]
- Artymiuk, P.J.; Green, J. The double life of aconitase. Structure 2006, 14, 2–4. [Google Scholar] [CrossRef]
- Banerjee, S.; Nandyala, A.K.; Raviprasad, P.; Ahmed, N.; Hasnain, S.E. Iron-dependent RNA-binding activity of Mycobacterium tuberculosis aconitase. J. Bacteriol. 2007, 189, 4046–4052. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, M.C.; Mende-Mueller, L.; Blondin, G.A.; Beinert, H. Purification and characterization of cytosolic aconitase from beef liver and its relationship to the iron-responsive element binding protein. Proc. Natl. Acad. Sci. USA 1992, 89, 11730–11734. [Google Scholar] [CrossRef] [Green Version]
- Howard, P.K.; Shaw, J.; Otsuka, A.J. Nucleotide sequence of the birA gene encoding the biotin operon repressor and biotin holoenzyme synthetase functions of Escherichia coli. Gene 1985, 35, 321–331. [Google Scholar] [CrossRef]
- Barker, D.F.; Campbell, A.M. Genetic and biochemical characterization of the birA gene and its product: Evidence for a direct role of biotin holoenzyme synthetase in repression of the biotin operon in Escherichia coli. J. Mol. Biol. 1981, 146, 469–492. [Google Scholar] [CrossRef]
- Wan, F.; Anderson, D.E.; Barnitz, R.A.; Snow, A.; Bidere, N.; Zheng, L.; Hegde, V.; Lam, L.T.; Staudt, L.M.; Levens, D.; et al. Ribosomal protein S3: A KH domain subunit in NF-kappaB complexes that mediates selective gene regulation. Cell 2007, 131, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Malygin, A.A.; Parakhnevitch, N.M.; Ivanov, A.V.; Eperon, I.C.; Karpova, G.G. Human ribosomal protein S13 regulates expression of its own gene at the splicing step by a feedback mechanism. Nucleic Acids Res. 2007, 35, 6414–6423. [Google Scholar] [CrossRef] [Green Version]
- Fewell, S.W.; Woolford, J.L., Jr. Ribosomal protein S14 of Saccharomyces cerevisiae regulates its expression by binding to RPS14B pre-mRNA and to 18S rRNA. Mol. Cell Biol. 1999, 19, 826–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presutti, C.; Ciafre, S.A.; Bozzoni, I. The ribosomal protein L2 in S. cerevisiae controls the level of accumulation of its own mRNA. EMBO J. 1991, 10, 2215–2221. [Google Scholar] [CrossRef]
- Zengel, J.M.; Lindahl, L. A hairpin structure upstream of the terminator hairpin required for ribosomal protein L4-mediated attenuation control of the S10 operon of Escherichia coli. J. Bacteriol. 1996, 178, 2383–2387. [Google Scholar] [CrossRef] [Green Version]
- Horn, H.F.; Vousden, K.H. Cooperation between the ribosomal proteins L5 and L11 in the p53 pathway. Oncogene 2008, 27, 5774–5784. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, J.F.; Tran, E.J.; Maxwell, E.S. Archaeal ribosomal protein L7 is a functional homolog of the eukaryotic 15.5kD/Snu13p snoRNP core protein. Nucleic Acids Res. 2002, 30, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Lohrum, M.A.; Ludwig, R.L.; Kubbutat, M.H.; Hanlon, M.; Vousden, K.H. Regulation of HDM2 activity by the ribosomal protein L11. Cancer Cell 2003, 3, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, B.; Sampath, P.; Seshadri, V.; Maitra, R.K.; DiCorleto, P.E.; Fox, P.L. Regulated release of L13a from the 60S ribosomal subunit as a mechanism of transcript-specific translational control. Cell 2003, 115, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.S.; Zeng, S.X.; Jin, Y.; Sun, X.X.; David, L.; Lu, H. Ribosomal protein L23 activates p53 by inhibiting MDM2 function in response to ribosomal perturbation but not to translation inhibition. Mol. Cell Biol. 2004, 24, 7654–7668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, M.; Absalon, M.J.; McLure, K.G.; Kastan, M.B. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell 2005, 123, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.M., 3rd; Deutsch, W.A.; Kelley, M.R. Drosophila ribosomal protein S3 contains an activity that cleaves DNA at apurinic/apyrimidinic sites. J. Biol. Chem. 1994, 269, 25359–25364. [Google Scholar] [PubMed]
- Chen, X.J.; Wang, X.; Kaufman, B.A.; Butow, R.A. Aconitase couples metabolic regulation to mitochondrial DNA maintenance. Science 2005, 307, 714–717. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two enzymes in one; two yeast peroxiredoxins display oxidative stress-dependent switching from a peroxidase to a molecular chaperone function. Cell 2004, 117, 625–635. [Google Scholar] [CrossRef]
- Krojer, T.; Garrido-Franco, M.; Huber, R.; Ehrmann, M.; Clausen, T. Crystal structure of DegP (HtrA) reveals a new protease-chaperone machine. Nature 2002, 416, 455–459. [Google Scholar] [CrossRef]
- Jeffery, C.J. An enzyme in the test tube, and a transcription factor in the cell: Moonlighting proteins and cellular factors that affect their behavior. Protein Sci. 2019, 28, 1233–1238. [Google Scholar] [CrossRef]
- Romero, P.; Obradovic, Z.; Dunker, A.K. Natively disordered proteins: Functions and predictions. Appl. Bioinform. 2004, 3, 105–113. [Google Scholar] [CrossRef]
- Dyson, H.J. Making Sense of Intrinsically Disordered Proteins. Biophys. J. 2016, 110, 1013–1016. [Google Scholar] [CrossRef] [Green Version]
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; et al. Intrinsically disordered protein. J. Mol. Graph. Model. 2001, 19, 26–59. [Google Scholar] [CrossRef] [Green Version]
- Tompa, P.; Szasz, C.; Buday, L. Structural disorder throws new light on moonlighting. Trends Biochem. Sci. 2005, 30, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.E.; Dyson, H.J. Intrinsically unstructured proteins: Re-assessing the protein structure-function paradigm. J. Mol. Biol. 1999, 293, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunker, A.K.; Brown, C.J.; Lawson, J.D.; Iakoucheva, L.M.; Obradovic, Z. Intrinsic disorder and protein function. Biochemistry 2002, 41, 6573–6582. [Google Scholar] [CrossRef] [Green Version]
- Hatos, A.; Hajdu-Soltesz, B.; Monzon, A.M.; Palopoli, N.; Alvarez, L.; Aykac-Fas, B.; Bassot, C.; Benitez, G.I.; Bevilacqua, M.; Chasapi, A.; et al. DisProt: Intrinsic protein disorder annotation in 2020. Nucleic Acids Res. 2020, 48, D269–D276. [Google Scholar] [CrossRef] [Green Version]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the p53 network. Nature 2000, 408, 307–310. [Google Scholar] [CrossRef]
- Green, D.R.; Kroemer, G. Cytoplasmic functions of the tumour suppressor p53. Nature 2009, 458, 1127–1130. [Google Scholar] [CrossRef]
- Zhou, J.; Ahn, J.; Wilson, S.H.; Prives, C. A role for p53 in base excision repair. EMBO J. 2001, 20, 914–923. [Google Scholar] [CrossRef] [Green Version]
- Clore, G.M.; Omichinski, J.G.; Sakaguchi, K.; Zambrano, N.; Sakamoto, H.; Appella, E.; Gronenborn, A.M. High-resolution structure of the oligomerization domain of p53 by multidimensional NMR. Science 1994, 265, 386–391. [Google Scholar] [CrossRef]
- Wells, M.; Tidow, H.; Rutherford, T.J.; Markwick, P.; Jensen, M.R.; Mylonas, E.; Svergun, D.I.; Blackledge, M.; Fersht, A.R. Structure of tumor suppressor p53 and its intrinsically disordered N-terminal transactivation domain. Proc. Natl. Acad. Sci. USA 2008, 105, 5762–5767. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.; Klein, C.; Muller, L.; Hansen, S.; Buchner, J. p53 contains large unstructured regions in its native state. J. Mol. Biol. 2002, 322, 917–927. [Google Scholar] [CrossRef]
- Safer, D.; Elzinga, M.; Nachmias, V.T. Thymosin beta 4 and Fx, an actin-sequestering peptide, are indistinguishable. J. Biol. Chem. 1991, 266, 4029–4032. [Google Scholar] [PubMed]
- Cassimeris, L.; Safer, D.; Nachmias, V.T.; Zigmond, S.H. Thymosin beta 4 sequesters the majority of G-actin in resting human polymorphonuclear leukocytes. J. Cell Biol. 1992, 119, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.S.; Rose, W.; Yaen, C.; Goldstein, A.; Martinez, J.; Kleinman, H. Thymosin beta4 enhances endothelial cell differentiation and angiogenesis. Angiogenesis 1999, 3, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Smart, N.; Rossdeutsch, A.; Riley, P.R. Thymosin beta4 and angiogenesis: Modes of action and therapeutic potential. Angiogenesis 2007, 10, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Philp, D.; Kleinman, H.K. Animal studies with thymosin beta, a multifunctional tissue repair and regeneration peptide. Ann. NY Acad. Sci. 2010, 1194, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.D.; Lawrence, A.J.; MacLean, A.G.; Leung, B.P.; McInnes, I.B.; Canas, B.; Pappin, D.J.; Stevenson, R.D. Thymosin beta 4 sulfoxide is an anti-inflammatory agent generated by monocytes in the presence of glucocorticoids. Nat. Med. 1999, 5, 1424–1427. [Google Scholar] [CrossRef] [PubMed]
- Safer, D.; Sosnick, T.R.; Elzinga, M. Thymosin beta 4 binds actin in an extended conformation and contacts both the barbed and pointed ends. Biochemistry 1997, 36, 5806–5816. [Google Scholar] [CrossRef]
- Domanski, M.; Hertzog, M.; Coutant, J.; Gutsche-Perelroizen, I.; Bontems, F.; Carlier, M.F.; Guittet, E.; van Heijenoort, C. Coupling of folding and binding of thymosin beta4 upon interaction with monomeric actin monitored by nuclear magnetic resonance. J. Biol. Chem. 2004, 279, 23637–23645. [Google Scholar] [CrossRef] [Green Version]
- Bock-Marquette, I.; Saxena, A.; White, M.D.; Dimaio, J.M.; Srivastava, D. Thymosin beta4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature 2004, 432, 466–472. [Google Scholar] [CrossRef]
- Ho, J.H.; Tseng, K.C.; Ma, W.H.; Chen, K.H.; Lee, O.K.; Su, Y. Thymosin beta-4 upregulates anti-oxidative enzymes and protects human cornea epithelial cells against oxidative damage. Br. J. Ophthalmol. 2008, 92, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Tantos, A.; Szabo, B.; Lang, A.; Varga, Z.; Tsylonok, M.; Bokor, M.; Verebelyi, T.; Kamasa, P.; Tompa, K.; Perczel, A.; et al. Multiple fuzzy interactions in the moonlighting function of thymosin-beta4. Intrinsically Disord Proteins 2013, 1, e26204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czisch, M.; Schleicher, M.; Horger, S.; Voelter, W.; Holak, T.A. Conformation of thymosin beta 4 in water determined by NMR spectroscopy. Eur. J. Biochem. 1993, 218, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Hope, I.A.; Struhl, K. Functional dissection of a eukaryotic transcriptional activator protein, GCN4 of yeast. Cell 1986, 46, 885–894. [Google Scholar] [CrossRef]
- Nikolaev, Y.; Deillon, C.; Hoffmann, S.R.; Bigler, L.; Friess, S.; Zenobi, R.; Pervushin, K.; Hunziker, P.; Gutte, B. The leucine zipper domains of the transcription factors GCN4 and c-Jun have ribonuclease activity. PLoS ONE 2010, 5, e10765. [Google Scholar] [CrossRef]
- Weiss, M.A.; Ellenberger, T.; Wobbe, C.R.; Lee, J.P.; Harrison, S.C.; Struhl, K. Folding transition in the DNA-binding domain of GCN4 on specific binding to DNA. Nature 1990, 347, 575–578. [Google Scholar] [CrossRef]
- Leon-Del-Rio, A.; Valadez-Graham, V.; Gravel, R.A. Holocarboxylase Synthetase: A Moonlighting Transcriptional Coregulator of Gene Expression and a Cytosolic Regulator of Biotin Utilization. Annu. Rev. Nutr. 2017, 37, 207–223. [Google Scholar] [CrossRef]
- Wilson, K.P.; Shewchuk, L.M.; Brennan, R.G.; Otsuka, A.J.; Matthews, B.W. Escherichia coli biotin holoenzyme synthetase/bio repressor crystal structure delineates the biotin- and DNA-binding domains. Proc. Natl. Acad. Sci. USA 1992, 89, 9257–9261. [Google Scholar] [CrossRef] [Green Version]
- Merenmies, J.; Pihlaskari, R.; Laitinen, J.; Wartiovaara, J.; Rauvala, H. 30-kDa heparin-binding protein of brain (amphoterin) involved in neurite outgrowth. Amino acid sequence and localization in the filopodia of the advancing plasma membrane. J. Biol. Chem. 1991, 266, 16722–16729. [Google Scholar]
- Hardman, C.H.; Broadhurst, R.W.; Raine, A.R.; Grasser, K.D.; Thomas, J.O.; Laue, E.D. Structure of the A-domain of HMG1 and its interaction with DNA as studied by heteronuclear three- and four-dimensional NMR spectroscopy. Biochemistry 1995, 34, 16596–16607. [Google Scholar] [CrossRef]
- Stott, K.; Watson, M.; Howe, F.S.; Grossmann, J.G.; Thomas, J.O. Tail-mediated collapse of HMGB1 is dynamic and occurs via differential binding of the acidic tail to the A and B domains. J. Mol. Biol. 2010, 403, 706–722. [Google Scholar] [CrossRef] [PubMed]
- Brunagel, G.; Shah, U.; Schoen, R.E.; Getzenberg, R.H. Identification of calreticulin as a nuclear matrix protein associated with human colon cancer. J. Cell Biochem. 2003, 89, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Goicoechea, S.; Orr, A.W.; Pallero, M.A.; Eggleton, P.; Murphy-Ullrich, J.E. Thrombospondin mediates focal adhesion disassembly through interactions with cell surface calreticulin. J. Biol. Chem. 2000, 275, 36358–36368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, M.; Stafford, W.F. Probing the three-dimensional structure of human calreticulin. Biochemistry 2000, 39, 14950–14959. [Google Scholar] [CrossRef] [PubMed]
- Yates, J.L.; Nomura, M.E. coli ribosomal protein L4 is a feedback regulatory protein. Cell 1980, 21, 517–522. [Google Scholar] [CrossRef]
- Timsit, Y.; Acosta, Z.; Allemand, F.; Chiaruttini, C.; Springer, M. The role of disordered ribosomal protein extensions in the early steps of eubacterial 50 S ribosomal subunit assembly. Int. J. Mol. Sci. 2009, 10, 817–834. [Google Scholar] [CrossRef] [Green Version]
- Wohlschlegel, J.A.; Dwyer, B.T.; Dhar, S.K.; Cvetic, C.; Walter, J.C.; Dutta, A. Inhibition of eukaryotic DNA replication by geminin binding to Cdt1. Science 2000, 290, 2309–2312. [Google Scholar] [CrossRef]
- Varma, D.; Chandrasekaran, S.; Sundin, L.J.; Reidy, K.T.; Wan, X.; Chasse, D.A.; Nevis, K.R.; DeLuca, J.G.; Salmon, E.D.; Cook, J.G. Recruitment of the human Cdt1 replication licensing protein by the loop domain of Hec1 is required for stable kinetochore-microtubule attachment. Nat. Cell Biol. 2012, 14, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Khayrutdinov, B.I.; Bae, W.J.; Yun, Y.M.; Lee, J.H.; Tsuyama, T.; Kim, J.J.; Hwang, E.; Ryu, K.S.; Cheong, H.K.; Cheong, C.; et al. Structure of the Cdt1 C-terminal domain: Conservation of the winged helix fold in replication licensing factors. Protein Sci. 2009, 18, 2252–2264. [Google Scholar] [CrossRef] [Green Version]
- Coschigano, P.W.; Magasanik, B. The URE2 gene product of Saccharomyces cerevisiae plays an important role in the cellular response to the nitrogen source and has homology to glutathione s-transferases. Mol. Cell Biol. 1991, 11, 822–832. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Zhou, J.M.; Perrett, S. The yeast prion protein Ure2 shows glutathione peroxidase activity in both native and fibrillar forms. J. Biol. Chem. 2004, 279, 50025–50030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, M.M.; Baxa, U.; Steven, A.C.; Bax, A.; Wickner, R.B. Is the prion domain of soluble Ure2p unstructured? Biochemistry 2005, 44, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Costi, M.P.; Ferrari, S.; Venturelli, A.; Calo, S.; Tondi, D.; Barlocco, D. Thymidylate synthase structure, function and implication in drug discovery. Curr. Med. Chem. 2005, 12, 2241–2258. [Google Scholar] [CrossRef] [PubMed]
- Chu, E.; Koeller, D.M.; Casey, J.L.; Drake, J.C.; Chabner, B.A.; Elwood, P.C.; Zinn, S.; Allegra, C.J. Autoregulation of human thymidylate synthase messenger RNA translation by thymidylate synthase. Proc. Natl. Acad. Sci. USA 1991, 88, 8977–8981. [Google Scholar] [CrossRef] [Green Version]
- Phan, J.; Steadman, D.J.; Koli, S.; Ding, W.C.; Minor, W.; Dunlap, R.B.; Berger, S.H.; Lebioda, L. Structure of human thymidylate synthase suggests advantages of chemotherapy with noncompetitive inhibitors. J. Biol. Chem. 2001, 276, 14170–14177. [Google Scholar] [CrossRef] [Green Version]
- Margoliash, E.; Smith, E.L.; Kreil, G.; Tuppy, H. Amino-acid sequence of horse heart cytochrome c. Nature 1961, 192, 1125–1127. [Google Scholar] [CrossRef]
- Yu, T.; Wang, X.; Purring-Koch, C.; Wei, Y.; McLendon, G.L. A mutational epitope for cytochrome C binding to the apoptosis protease activation factor-1. J. Biol. Chem. 2001, 276, 13034–13038. [Google Scholar] [CrossRef] [Green Version]
- Stellwagen, E.; Rysavy, R.; Babul, G. The conformation of horse heart apocytochrome c. J. Biol. Chem. 1972, 247, 8074–8077. [Google Scholar]
- Lux, S.E.; John, K.M.; Kopito, R.R.; Lodish, H.F. Cloning and characterization of band 3, the human erythrocyte anion-exchange protein (AE1). Proc. Natl. Acad. Sci. USA 1989, 86, 9089–9093. [Google Scholar] [CrossRef] [Green Version]
- Low, P.S.; Rathinavelu, P.; Harrison, M.L. Regulation of glycolysis via reversible enzyme binding to the membrane protein, band 3. J. Biol. Chem. 1993, 268, 14627–14631. [Google Scholar]
- Zhang, D.; Kiyatkin, A.; Bolin, J.T.; Low, P.S. Crystallographic structure and functional interpretation of the cytoplasmic domain of erythrocyte membrane band 3. Blood 2000, 96, 2925–2933. [Google Scholar] [CrossRef]
- Zielenski, J.; Rozmahel, R.; Bozon, D.; Kerem, B.; Grzelczak, Z.; Riordan, J.R.; Rommens, J.; Tsui, L.C. Genomic DNA sequence of the cystic fibrosis transmembrane conductance regulator (CFTR) gene. Genomics 1991, 10, 214–228. [Google Scholar] [CrossRef]
- Stutts, M.J.; Canessa, C.M.; Olsen, J.C.; Hamrick, M.; Cohn, J.A.; Rossier, B.C.; Boucher, R.C. CFTR as a cAMP-dependent regulator of sodium channels. Science 1995, 269, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Ostedgaard, L.S.; Baldursson, O.; Vermeer, D.W.; Welsh, M.J.; Robertson, A.D. A functional R domain from cystic fibrosis transmembrane conductance regulator is predominantly unstructured in solution. Proc. Natl. Acad. Sci. USA 2000, 97, 5657–5662. [Google Scholar] [CrossRef] [Green Version]
- Fuxreiter, M. Fuzziness in Protein Interactions-A Historical Perspective. J. Mol. Biol. 2018, 430, 2278–2287. [Google Scholar] [CrossRef]
- Tompa, P.; Fuxreiter, M. Fuzzy complexes: Polymorphism and structural disorder in protein-protein interactions. Trends Biochem. Sci. 2008, 33, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Raduly, Z.; Miskei, M.; Fuxreiter, M. Fuzzy complexes: Specific binding without complete folding. FEBS Lett. 2015, 589, 2533–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uversky, V.N. Supramolecular Fuzziness of Intracellular Liquid Droplets: Liquid-Liquid Phase Transitions, Membrane-Less Organelles, and Intrinsic Disorder. Molecules 2019, 24, 3265. [Google Scholar] [CrossRef] [Green Version]
- Miskei, M.; Antal, C.; Fuxreiter, M. FuzDB: Database of fuzzy complexes, a tool to develop stochastic structure-function relationships for protein complexes and higher-order assemblies. Nucleic Acids Res. 2017, 45, D228–D235. [Google Scholar] [CrossRef] [Green Version]
- Murzin, A.G. Biochemistry. Metamorphic proteins. Science 2008, 320, 1725–1726. [Google Scholar] [CrossRef]
- Dishman, A.F.; Volkman, B.F. Unfolding the Mysteries of Protein Metamorphosis. Acs. Chem. Biol. 2018, 13, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Lella, M.; Mahalakshmi, R. Metamorphic Proteins: Emergence of Dual Protein Folds from One Primary Sequence. Biochemistry 2017, 56, 2971–2984. [Google Scholar] [CrossRef] [PubMed]
- Goodchild, S.C.; Curmi, P.M.G.; Brown, L.J. Structural gymnastics of multifunctional metamorphic proteins. Biophys. Rev. 2011, 3, 143. [Google Scholar] [CrossRef] [PubMed]
- Tuinstra, R.L.; Peterson, F.C.; Kutlesa, S.; Elgin, E.S.; Kron, M.A.; Volkman, B.F. Interconversion between two unrelated protein folds in the lymphotactin native state. Proc. Natl. Acad. Sci. USA 2008, 105, 5057–5062. [Google Scholar] [CrossRef] [Green Version]
- Burmann, B.M.; Knauer, S.H.; Sevostyanova, A.; Schweimer, K.; Mooney, R.A.; Landick, R.; Artsimovitch, I.; Rosch, P. An alpha helix to beta barrel domain switch transforms the transcription factor RfaH into a translation factor. Cell 2012, 150, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Goodchild, S.C.; Howell, M.W.; Littler, D.R.; Mandyam, R.A.; Sale, K.L.; Mazzanti, M.; Breit, S.N.; Curmi, P.M.; Brown, L.J. Metamorphic response of the CLIC1 chloride intracellular ion channel protein upon membrane interaction. Biochemistry 2010, 49, 5278–5289. [Google Scholar] [CrossRef]
- Luo, X.; Yu, H. Protein metamorphosis: The two-state behavior of Mad2. Structure 2008, 16, 1616–1625. [Google Scholar] [CrossRef] [Green Version]
- Tseng, R.; Goularte, N.F.; Chavan, A.; Luu, J.; Cohen, S.E.; Chang, Y.G.; Heisler, J.; Li, S.; Michael, A.K.; Tripathi, S.; et al. Structural basis of the day-night transition in a bacterial circadian clock. Science 2017, 355, 1174–1180. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Tonelli, M.; Markley, J.L. Metamorphic protein IscU changes conformation by cis-trans isomerizations of two peptidyl-prolyl peptide bonds. Biochemistry 2012, 51, 9595–9602. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Pelegrin, M.; Cerda-Costa, N.; Cintas-Pedrola, A.; Herranz-Trillo, F.; Bernado, P.; Peinado, J.R.; Arolas, J.L.; Gomis-Ruth, F.X. Multiple stable conformations account for reversible concentration-dependent oligomerization and autoinhibition of a metamorphic metallopeptidase. Angew. Chem. Int. Ed. Engl. 2014, 53, 10624–10630. [Google Scholar] [CrossRef] [Green Version]
- London, R.E. HIV-1 Reverse Transcriptase: A Metamorphic Protein with Three Stable States. Structure 2019, 27, 420–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, F.C.; Elgin, E.S.; Nelson, T.J.; Zhang, F.; Hoeger, T.J.; Linhardt, R.J.; Volkman, B.F. Identification and characterization of a glycosaminoglycan recognition element of the C chemokine lymphotactin. J. Biol. Chem. 2004, 279, 12598–12604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuinstra, R.L.; Peterson, F.C.; Elgin, E.S.; Pelzek, A.J.; Volkman, B.F. An engineered second disulfide bond restricts lymphotactin/XCL1 to a chemokine-like conformation with XCR1 agonist activity. Biochemistry 2007, 46, 2564–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knauer, S.H.; Artsimovitch, I.; Rosch, P. Transformer proteins. Cell Cycle 2012, 11, 4289–4290. [Google Scholar] [CrossRef] [Green Version]
- Al Khamici, H.; Brown, L.J.; Hossain, K.R.; Hudson, A.L.; Sinclair-Burton, A.A.; Ng, J.P.; Daniel, E.L.; Hare, J.E.; Cornell, B.A.; Curmi, P.M.; et al. Members of the chloride intracellular ion channel protein family demonstrate glutaredoxin-like enzymatic activity. PLoS ONE 2015, 10, e115699. [Google Scholar] [CrossRef] [Green Version]
- Littler, D.R.; Harrop, S.J.; Fairlie, W.D.; Brown, L.J.; Pankhurst, G.J.; Pankhurst, S.; DeMaere, M.Z.; Campbell, T.J.; Bauskin, A.R.; Tonini, R.; et al. The intracellular chloride ion channel protein CLIC1 undergoes a redox-controlled structural transition. J. Biol. Chem. 2004, 279, 9298–9305. [Google Scholar] [CrossRef] [Green Version]
- Warton, K.; Tonini, R.; Fairlie, W.D.; Matthews, J.M.; Valenzuela, S.M.; Qiu, M.R.; Wu, W.M.; Pankhurst, S.; Bauskin, A.R.; Harrop, S.J.; et al. Recombinant CLIC1 (NCC27) assembles in lipid bilayers via a pH-dependent two-state process to form chloride ion channels with identical characteristics to those observed in Chinese hamster ovary cells expressing CLIC1. J. Biol. Chem. 2002, 277, 26003–26011. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, E.K. Morpheeins—A new structural paradigm for allosteric regulation. Trends Biochem. Sci. 2005, 30, 490–497. [Google Scholar] [CrossRef]
- Jaffe, E.K.; Lawrence, S.H. Allostery and the dynamic oligomerization of porphobilinogen synthase. Arch. Biochem. Biophys. 2012, 519, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.G.; Gu, M.; Etlinger, J.D. 240-kDa proteasome inhibitor (CF-2) is identical to delta-aminolevulinic acid dehydratase. J. Biol. Chem. 1994, 269, 12399–12402. [Google Scholar]
- Li, X.C.; Gu, M.Z.; Etlinger, J.D. Isolation and characterization of a novel endogenous inhibitor of the proteasome. Biochemistry 1991, 30, 9709–9715. [Google Scholar] [CrossRef] [PubMed]
- Breinig, S.; Kervinen, J.; Stith, L.; Wasson, A.S.; Fairman, R.; Wlodawer, A.; Zdanov, A.; Jaffe, E.K. Control of tetrapyrrole biosynthesis by alternate quaternary forms of porphobilinogen synthase. Nat. Struct. Biol. 2003, 10, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Kervinen, J.; Jaffe, E.K.; Stauffer, F.; Neier, R.; Wlodawer, A.; Zdanov, A. Mechanistic basis for suicide inactivation of porphobilinogen synthase by 4,7-dioxosebacic acid, an inhibitor that shows dramatic species selectivity. Biochemistry 2001, 40, 8227–8236. [Google Scholar] [CrossRef] [Green Version]
- Bornholdt, Z.A.; Noda, T.; Abelson, D.M.; Halfmann, P.; Wood, M.R.; Kawaoka, Y.; Saphire, E.O. Structural rearrangement of ebola virus VP40 begets multiple functions in the virus life cycle. Cell 2013, 154, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, J.; Poruri, K.; Boniecki, M.T.; McTavish, K.K.; Martinis, S.A. Yeast mitochondrial leucyl-tRNA synthetase CP1 domain has functionally diverged to accommodate RNA splicing at expense of hydrolytic editing. J. Biol. Chem. 2012, 287, 14772–14781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.M.; Jeong, S.J.; Park, M.C.; Kim, G.; Kwon, N.H.; Kim, H.K.; Ha, S.H.; Ryu, S.H.; Kim, S. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef] [Green Version]
- Eyers, P.A.; Murphy, J.M. The evolving world of pseudoenzymes: Proteins, prejudice and zombies. Bmc Biol. 2016, 14, 98. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M.; Farhan, H.; Eyers, P.A. Bio-Zombie: The rise of pseudoenzymes in biology. Biochem. Soc. Trans. 2017, 45, 537–544. [Google Scholar] [CrossRef]
- Jeffery, C.J. The demise of catalysis, but new functions arise: Pseudoenzymes as the phoenixes of the protein world. Biochem. Soc. Trans. 2019, 47, 371–379. [Google Scholar] [CrossRef]
- Todd, A.E.; Orengo, C.A.; Thornton, J.M. Sequence and structural differences between enzyme and nonenzyme homologs. Structure 2002, 10, 1435–1451. [Google Scholar] [CrossRef] [Green Version]
- Pils, B.; Schultz, J. Inactive enzyme-homologues find new function in regulatory processes. J. Mol. Biol. 2004, 340, 399–404. [Google Scholar] [CrossRef]
- Qasba, P.K.; Safaya, S.K. Similarity of the nucleotide sequences of rat alpha-lactalbumin and chicken lysozyme genes. Nature 1984, 308, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Brew, K.; Vanaman, T.C.; Hill, R.L. Comparison of the amino acid sequence of bovine alpha-lactalbumin and hens egg white lysozyme. J. Biol. Chem. 1967, 242, 3747–3749. [Google Scholar] [PubMed]
- Powell, J.T.; Brew, K. On the interaction of alpha-lactalbumin and galactosyltransferase during lactose synthesis. J. Biol. Chem. 1975, 250, 6337–6343. [Google Scholar] [PubMed]
- Piatigorsky, J.; O’Brien, W.E.; Norman, B.L.; Kalumuck, K.; Wistow, G.J.; Borras, T.; Nickerson, J.M.; Wawrousek, E.F. Gene sharing by delta-crystallin and argininosuccinate lyase. Proc. Natl. Acad. Sci. USA 1988, 85, 3479–3483. [Google Scholar] [CrossRef] [Green Version]
- Sampaleanu, L.M.; Vallee, F.; Slingsby, C.; Howell, P.L. Structural studies of duck delta 1 and delta 2 crystallin suggest conformational changes occur during catalysis. Biochemistry 2001, 40, 2732–2742. [Google Scholar] [CrossRef]
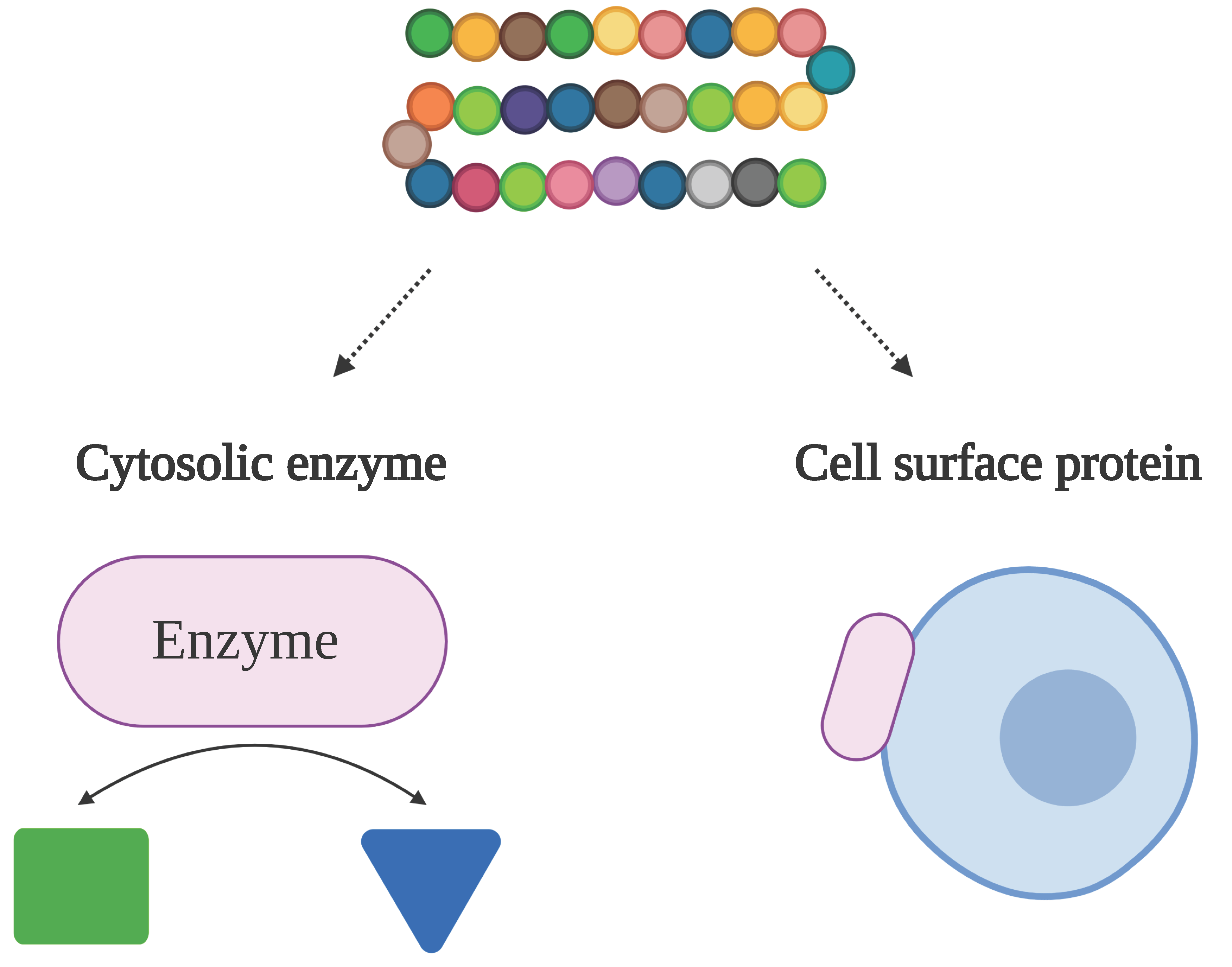
Figure 1.
In a moonlighting protein (purple oval), more than one physiologically relevant biochemical or biophysical function is performed by a single polypeptide chain. Note: This figure was “Created with BioRender.com”.
Figure 1.
In a moonlighting protein (purple oval), more than one physiologically relevant biochemical or biophysical function is performed by a single polypeptide chain. Note: This figure was “Created with BioRender.com”.

Figure 2.
Examples of moonlighting proteins. (a) Enolase is a cytosolic enzyme in glycolysis and also a plasminogen receptor when displayed on the cell surface (PDB ID: 1W6T [31]). (b) Aconitase is an enzyme in the citric acid cycle when it contains an iron/sulfur cluster bound in the active site of the protein (PDB ID: 2B3Y [32]). When the cellular iron level decreases and the iron/sulfur cluster disassociates, aconitase undergoes a large conformational change that enables it to bind to iron-responsive elements in mRNA (PDB ID: 3SNP [33]) to promote the expression of proteins involved in iron uptake. (c) Under normal cellular conditions, peroxiredoxin is predominantly a dimer (PDB ID: 5B6M [34]) that functions as a peroxidase that converts hydrogen peroxide to water. Under heat shock or oxidative stress, it converts to a higher molecular weight form, a decamer, that acts as a molecular chaperone that assists with protein folding (PDB ID: 6E0F [35]). Note: This figure was “Created with BioRender.com”, and the visualizations of the protein structures were created with Mol* [36] on the RCSB PDB website (rcsb.org) [37,38].
Figure 2.
Examples of moonlighting proteins. (a) Enolase is a cytosolic enzyme in glycolysis and also a plasminogen receptor when displayed on the cell surface (PDB ID: 1W6T [31]). (b) Aconitase is an enzyme in the citric acid cycle when it contains an iron/sulfur cluster bound in the active site of the protein (PDB ID: 2B3Y [32]). When the cellular iron level decreases and the iron/sulfur cluster disassociates, aconitase undergoes a large conformational change that enables it to bind to iron-responsive elements in mRNA (PDB ID: 3SNP [33]) to promote the expression of proteins involved in iron uptake. (c) Under normal cellular conditions, peroxiredoxin is predominantly a dimer (PDB ID: 5B6M [34]) that functions as a peroxidase that converts hydrogen peroxide to water. Under heat shock or oxidative stress, it converts to a higher molecular weight form, a decamer, that acts as a molecular chaperone that assists with protein folding (PDB ID: 6E0F [35]). Note: This figure was “Created with BioRender.com”, and the visualizations of the protein structures were created with Mol* [36] on the RCSB PDB website (rcsb.org) [37,38].

Figure 3.
Structures of lymphotactin, a metamorphic protein that is also a moonlighting protein. There are two tertiary folds for lymphotactin, Ltn10 and Ltn40. Ltn10 has a classical chemokine fold with a mix of alpha-helix and beta-sheet (PDB ID: 2HDM [134]). Ltn40 possesses a dimeric form with each subunit composed of mainly beta-sheets (PDB ID: 2JP1 [125]). Note: This figure was “Created with BioRender.com”, and the visualizations of the protein structures were created with Mol* [36] on the RCSB PDB website (rcsb.org) [37,38].
Figure 3.
Structures of lymphotactin, a metamorphic protein that is also a moonlighting protein. There are two tertiary folds for lymphotactin, Ltn10 and Ltn40. Ltn10 has a classical chemokine fold with a mix of alpha-helix and beta-sheet (PDB ID: 2HDM [134]). Ltn40 possesses a dimeric form with each subunit composed of mainly beta-sheets (PDB ID: 2JP1 [125]). Note: This figure was “Created with BioRender.com”, and the visualizations of the protein structures were created with Mol* [36] on the RCSB PDB website (rcsb.org) [37,38].

Figure 4.
Structures of porphobilinogen synthase, which is both a morpheein and a moonlighting protein. Porphobilinogen synthase can form two homo-multimers, a low activity hexamer (PDB ID: 1PV8 [143]) and a high activity octamer (PDB ID: 1I8J [144]). The two multimers can interconvert through two homo-dimers, with different subunit conformations. In addition to its catalytic function, porphobilinogen synthase has a second function as an inhibitor of the proteasome. Note: This figure was “Created with BioRender.com”, and the visualizations of the protein structures were created with Mol* [36] on the RCSB PDB website (rcsb.org) [37,38].
Figure 4.
Structures of porphobilinogen synthase, which is both a morpheein and a moonlighting protein. Porphobilinogen synthase can form two homo-multimers, a low activity hexamer (PDB ID: 1PV8 [143]) and a high activity octamer (PDB ID: 1I8J [144]). The two multimers can interconvert through two homo-dimers, with different subunit conformations. In addition to its catalytic function, porphobilinogen synthase has a second function as an inhibitor of the proteasome. Note: This figure was “Created with BioRender.com”, and the visualizations of the protein structures were created with Mol* [36] on the RCSB PDB website (rcsb.org) [37,38].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Examples of moonlighting proteins that contain intrinsically disordered regions.
| Protein | Organism | Function 1 | Function 1 Location | Function 2 | Function 2 Location | Disorder Content | UniProt ID | Ref. |
| P53 | Homo sapiens | Binds to regulatory element in the genome | Nucleus | Centrosome duplication, induction of apoptosis, autophagy inhibition | Cytoplasm | 37.00% | P04637 | [67,69,71] |
| Thymosin beta-4 | Homo sapiens | Involved in sequestering G-actin in human polymorphonuclear leukocytes | Cytoplasm | Secreted anti-inflammatory agent | Extracellular | 100.00% | P62328 | [73,78,84] |
| General control protein GCN4 | Saccharomyces cerevisiae | Transcription factor | Nucleus | Ribonuclease | Cytoplasm | 32.38% | P03069 | [85,86,87] |
| Bifunctional ligase/repressor BirA | Escherichia coli | Biotin synthetase, biotin–[acetyl–CoA-carboxylase] ligase | Cytoplasm | Biotin operon repressor, activity depends on cellular concentration of biotin | Bound to DNA | 7.17% | P06709 | [42,43,88,89] |
| High mobility group protein B1 | Rattus norvegicus | Binds heparin. | Cytoplasm | DNA binding protein, without sequence specificity | Nucleus | 35.81% | P63159 | [90,91,92] |
| Calreticulin | Homo sapiens | Protein-folding chaperone | Endoplasmic reticulum | Adhesin | Cell surface | 100.00% | P27797 | [93,94,95] |
| 50S ribosomal protein L4 | Escherichia coli | Ribosomal protein, part of the 50S subunit | Cytoplasm | Transcriptional repressor, causes premature termination of transcription | Bound to DNA | 31.34% | P60723 | [48,96,97] |
| DNA replication factor Cdt1 | Homo sapiens | Helps with initiating DNA replication. | Nucleus | Role in mitosis, localizes to kinetochores through binding to Ndc80 complex | Cytoplasm | 2.20% | Q9H211 | [98,99,100] |
| Transcriptional regulator Ure2 | Saccharomyce cerevisiae | Binds to and inhibits GATA transcriptional activators GLN3 and GAT1 | Cytoplasm | Glutathione peroxidase, thiol:disulfide oxidoreductase | Cytoplasm | 31.64% | P23202 | [101,102,103] |
| Thymidylate synthase | Homo sapiens | Thymidylate synthase | Cytoplasm | mRNA translation inhibition | Cytoplasm | 15.65% | P04818 | [104,105,106] |
| Cytochrome c | Equus caballus | Electron carrier protein | Mitochondrion | Binds to apoptosis protease activation factor-1 and promotes apoptosis | Cytoplasm | 99.05% | P00004 | [107,108,109] |
| Band 3 anion transport protein | Homo sapiens | Transports inorganic anions across the plasma membrane | Plasma membrane | Scaffold protein providing binding sites for glycolytic enzymes | Plasma membrane | 5.93% | P02730 | [110,111,112] |
| Cystic fibrosis transmembrane conductance regulator CFTR | Homo sapiens | Chloride transporter, contains nucleotide binding domains that bind and hydrolyze ATP | Plasma membrane | Regulator of other ion channels | Plasma membrane | 12.50% | P13569 | [113,114,115] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, H.; Jeffery, C.J. Moonlighting Proteins in the Fuzzy Logic of Cellular Metabolism. Molecules 2020, 25, 3440. https://doi.org/10.3390/molecules25153440
AMA Style
Liu H, Jeffery CJ. Moonlighting Proteins in the Fuzzy Logic of Cellular Metabolism. Molecules. 2020; 25(15):3440. https://doi.org/10.3390/molecules25153440
Chicago/Turabian StyleLiu, Haipeng, and Constance J. Jeffery. 2020. "Moonlighting Proteins in the Fuzzy Logic of Cellular Metabolism" Molecules 25, no. 15: 3440. https://doi.org/10.3390/molecules25153440