Antinociceptive and Anti-Inflammatory Effects of Octacosanol from the Leaves of Sabicea grisea var. grisea in Mice

Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction and Isolation of Octacosanol
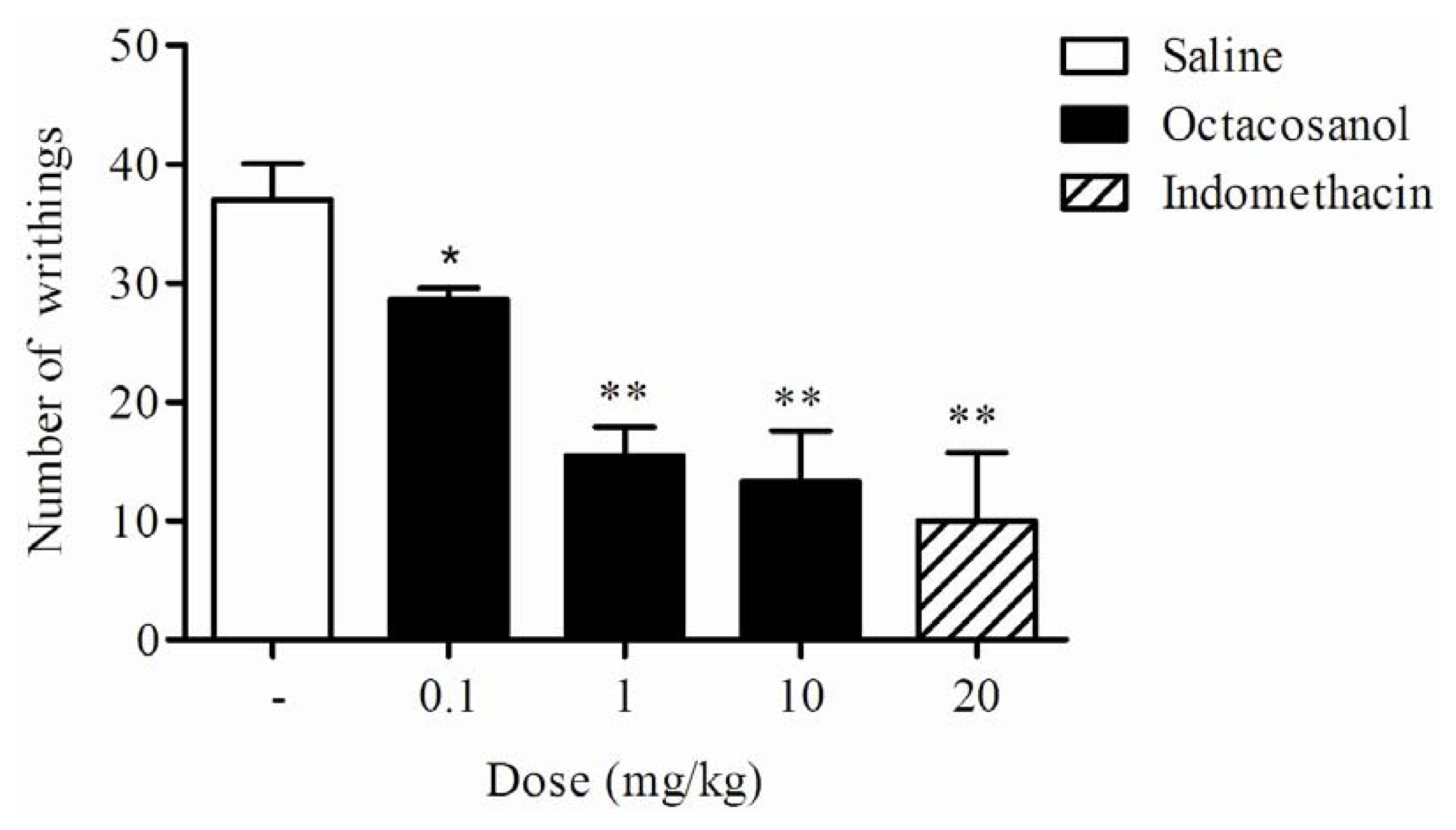
2.2. Writhing Response Induced by Acetic Acid in Mice
2.3. Effect on Rota-Rod Assay in Mice
2.4. Effect on Hot-Plate Latency Assay in Mice
2.5. Effects on Formalin-Induced Nociception in Mice
2.6. Effects on Carrageenan-Induced Pleurisy in Mice
3. Experimental Section
3.1. Plant Material
3.2. Preparation of Extracts, Purification and Structure Elucidation of Octacosanol
3.3. Animals
3.4. Acetic Acid-Induced Writhing Response in Mice
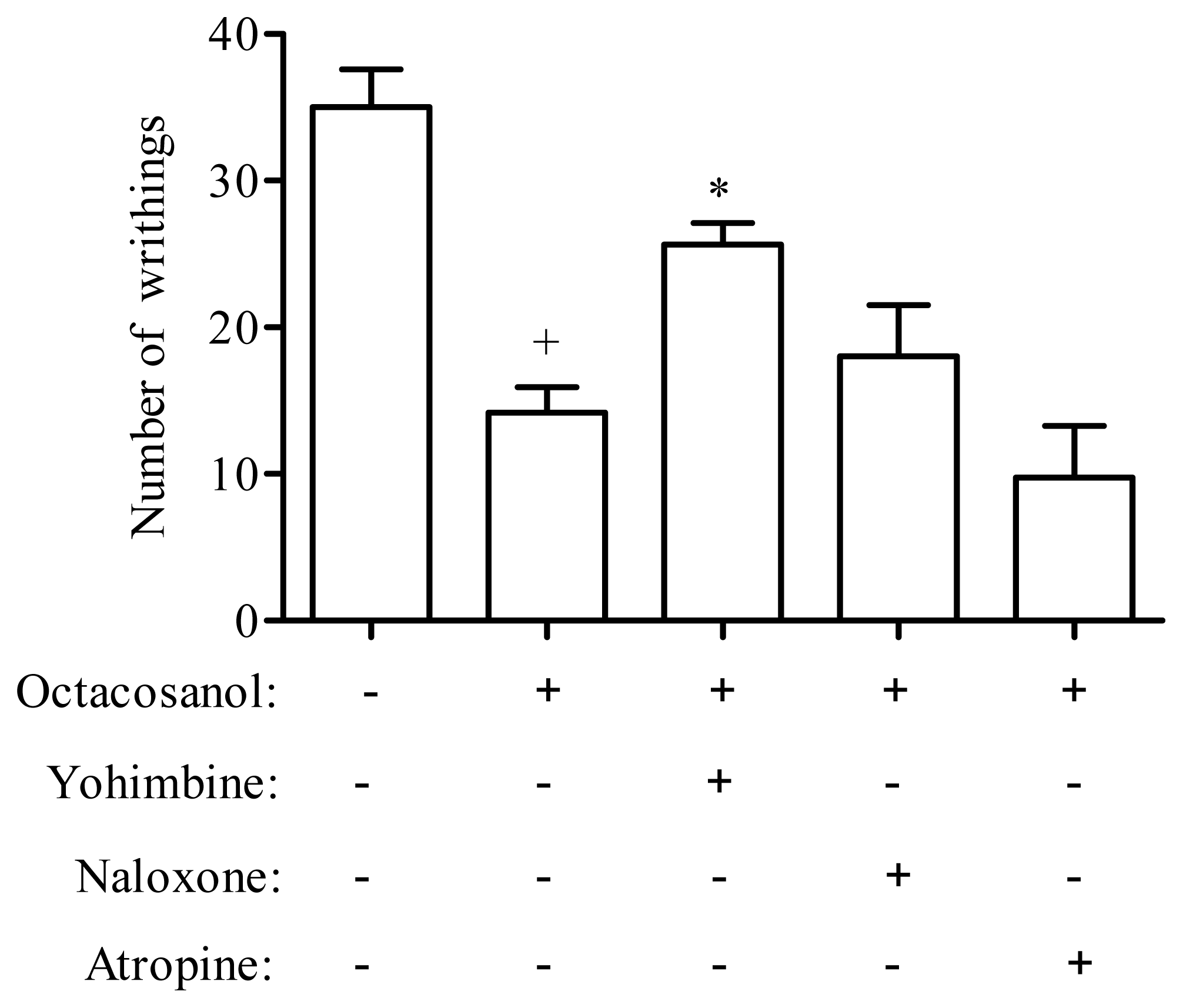
3.5. Analysis of Possible Antinociceptive Mechanisms of Octacosanol
3.6. Hot-Plate Latency Assay in Mice
3.7. Formalin-Induced Nociception in Mice
3.8. Rota-Rod Assay
3.9. Carrageenan-Induced Pleurisy in Mice
3.10. Quantification of TNF-α in Carrageenan-Induced Pleurisy
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Irmak, S.; Dunford, N.T. Policosanol contents and compositions of wheat varieties. J. Agric. Food Chem 2005, 53, 5583–5586. [Google Scholar]
- Saint-John, M.M.L. Octacosanol ingestion and its effects on metabolic responses to submaximal cycle ergometry, reaction time and chest and grip strength. Int. Clin. Nutr. Rev 1986, 1, 81–87. [Google Scholar]
- Wang, T.; Liu, Y.Y.; Wang, X.; Yang, N.; Zhu, H.B.; Zuo, P.P. Protective effects of octacosanol on 6-hydroxydopamine-induced Parkinsonism in rats via regulation of ProNGF and NGF signaling. Acta Pharmacol. Sin 2010, 31, 765–774. [Google Scholar]
- Thippeswamy, G.; Sheela, M.L.; Salimath, B.P. Octacosanol isolated from Tinospora cordifolia downregulates VEGF gene expression by inhibiting nuclear translocation of NF-<kappa>B and its DNA binding activity. Eur. J. Pharmacol 2008, 588, 141–150. [Google Scholar]
- Oliaro-Bosso, S.; Calcio Gaudino, E.; Mantegna, S.; Giraudo, E.; Meda, C.; Viola, F.; Cravotto, G. Regulation of HMGCoA reductase activity by policosanol and octacosadienol, a new synthetic analogue of octacosanol. Lipids 2009, 44, 907–916. [Google Scholar]
- Arruzazabala, M.L.; Carbajal, D.; Mas, R.; Molina, V.; Valdes, S.; Laguna, A. Cholesterol-lowering effects of policosanol in rabbits. Biol. Res 1994, 27, 205–208. [Google Scholar]
- Singh, D.K.; Li, L.; Porter, T.D. Policosanol inhibits cholesterol synthesis in hepatoma cells by activation of AMP-kinase. J. Pharmacol. Exp. Ther 2006, 318, 1020–1026. [Google Scholar]
- Arruzazabala, M.L.; Molina, V.; Mas, R.; Fernandez, L.; Carbajal, D.; Valdes, S.; Castano, G. Antiplatelet effects of policosanol (20 and 40 mg/day) in healthy volunteers and dyslipidaemic patients. Clin. Exp. Pharmacol. Physiol 2002, 29, 891–897. [Google Scholar]
- Arruzazabala, M.L.; Carbajal, D.; Mas, R.; Garcia, M.; Fraga, V. Effects of Policosanol on platelet aggregation in rats. Thromb. Res 1993, 69, 321–327. [Google Scholar]
- Carbajal, D.; Molina, V.; Valdes, S.; Arruzazabala, L.; Mas, R. Anti-ulcer activity of higher primary alcohols of beeswax. J. Pharm. Pharmacol 1995, 47, 731–733. [Google Scholar]
- Fernandez-Arche, A.; Marquez-Martin, A.; de la Puerta Vazquez, R.; Perona, J.S.; Terencio, C.; Perez-Camino, C.; Ruiz-Gutierrez, V. Long-chain fatty alcohols from pomace olive oil modulate the release of proinflammatory mediators. J. Nutr. Biochem 2009, 20, 155–162. [Google Scholar] [Green Version]
- Ravelo, Y.; Molina, V.; Carbajal, D.; Fernandez, L.; Fernandez, J.C.; Arruzazabala, M.L.; Mas, R. Evaluation of anti-inflammatory and antinociceptive effects of D-002 (beeswax alcohols). J. Nat. Med 2011, 65, 330–335. [Google Scholar]
- Awad, R.; Ahmed, F.; Bourbonnais-Spear, N.; Mullally, M.; Ta, C.A.; Tang, A.; Merali, Z.; Maquin, P.; Caal, F.; Cal, V.; et al. Ethnopharmacology of Q’eqchi’ Maya antiepileptic and anxiolytic plants: Effects on the GABAergic system. J. Ethnopharmacol 2009, 125, 257–264. [Google Scholar]
- Fagundes, L.L.; Vieira, G.D.; de Pinho, J.; Yamamoto, C.H.; Alves, M.S.; Stringheta, P.C.; de Sousa, O.V. Pharmacological proprieties of the ethanol extract of Muehlenbeckia platyclada (F. Muell.) Meisn. leaves. Int. J. Mol. Sci 2010, 11, 3942–3953. [Google Scholar]
- de Sousa, O.V.; Vieira, G.D.; De Pinho, J.J.R.G.; Yamamoto, C.H.; Alves, M.S. Antinociceptive and anti-inflammatory activities of the ethanol extract of Annona muricata L. leaves in animal models. Int. J. Mol. Sci 2010, 11, 2067–2078. [Google Scholar]
- Karou, S.D.; Tchacondo, T.; Ilboudo, D.P.; Simpore, J. Sub-Saharan Rubiaceae: A review of their traditional uses, phytochemistry and biological activities. Pak. J. Biol.Sci 2011, 14, 149–169. [Google Scholar]
- Mongrand, S.; Badoc, A.; Patouille, B.; Lacomblez, C.; Chavent, M.; Bessoule, J.J. Chemotaxonomy of the Rubiaceae family based on leaf fatty acid composition. Phytochemistry 2005, 66, 549–559. [Google Scholar]
- Cardoso, C.L.; Siqueira Silva, D.H.; Tomazela, D.M.; Verli, H.; Young, M.C.; Furlan, M.; Eberlin, M.N.; da Silva Bolzani, V. Turbinatine, a potential key intermediate in the biosynthesis of corynanthean-type indole alkaloids. J. Nat. Prod 2003, 66, 1017–1021. [Google Scholar]
- Tzakou, O.; Lempesis, K.; Loukis, A. Secondary metabolites from Asperula lutea subsp. rigidula. Nat. Prod. Commun 2011, 6, 237–238. [Google Scholar]
- Khan, S.A.; Razafimandimbison, S.G.; Bremer, B.; Liede-Schumann, S. Sabiceeae and Virectarieae (Rubiaceae, Ixoroideae): One or two tribes? New tribal and generic circumscriptions of Sabiceeae and biogeography of Sabicea s.l. Taxon 2008, 57, 7–23. [Google Scholar]
- Valadeau, C.; Castillo, J.A.; Sauvain, M.; Lores, A.F.; Bourdy, G. The rainbow hurts my skin: medicinal concepts and plants uses among the Yanesha (Amuesha), an Amazonian Peruvian ethnic group. J. Ethnopharmacol 2010, 127, 175–192. [Google Scholar]
- Roumy, V.; Garcia-Pizango, G.; Gutierrez-Choquevilca, A.L.; Ruiz, L.; Jullian, V.; Winterton, P.; Fabre, N.; Moulis, C.; Valentin, A. Amazonian plants from Peru used by Quechua and Mestizo to treat malaria with evaluation of their activity. J. Ethnopharmacol 2007, 112, 482–489. [Google Scholar]
- Collier, H.O.J.; Dinneen, L.C.; Johnson, C.A.; Schneide, C. Abdominal constriction response and its suppression by analgesic drugs in mouse. Br. J. Pharmacol 1968, 32, 295–310. [Google Scholar]
- Ribeiro, R.A.; Vale, M.L.; Thomazzi, S.M.; Paschoalato, A.B.; Poole, S.; Ferreira, S.H.; Cunha, F.Q. Involvement of resident macrophages and mast cells in the writhing nociceptive response induced by zymosan and acetic acid in mice. Eur. J. Pharmacol 2000, 387, 111–118. [Google Scholar]
- Amico-Roxas, M.; Caruso, A.; Trombadore, S.; Scifo, R.; Scapagnini, U. Gangliosides antinociceptive effects in rodents. Arch. Int. Pharmacodyn. Ther 1984, 272, 103–117. [Google Scholar]
- Yaksh, T.L. Pharmacology and mechanisms of opioid analgesic activity. Acta Anaesthesiol. Scand 1997, 41, 94–111. [Google Scholar]
- Schmauss, C.; Yaksh, T.L. In vivo studies on spinal opiate receptor systems mediating antinociception. II. Pharmacological profiles suggesting a differential association of mu, delta and kappa receptors with visceral chemical and cutaneous thermal stimuli in the rat. J. Pharmacol. Exp. Ther 1984, 228, 1–12. [Google Scholar]
- Luttinger, D.; Ferrari, R.; Perrone, M.H.; Haubrich, D.R. Pharmacological analysis of alpha-2 adrenergic mechanisms in nociception and ataxia. J. Pharmacol. Exp. Ther 1985, 232, 883–889. [Google Scholar]
- Iwamoto, E.T.; Marion, L. Characterization of the antinociception produced by intrathecally administered muscarinic agonists in rats. J. Pharmacol. Exp. Ther 1993, 266, 329–338. [Google Scholar]
- Le Bars, D.; Gozariu, M.; Cadden, S.W. Animal models of nociception. Pharmacol. Rev 2001, 53, 597–652. [Google Scholar]
- Dunham, N.W.; Miya, T.S. A note on a simple apparatus for detecting neurological deficit in rats and mice. J. Am. Pharm. Assoc 1957, 46, 208–209. [Google Scholar]
- McNamara, C.R.; Mandel-Brehm, J.; Bautista, D.M.; Siemens, J.; Deranian, K.L.; Zhao, M.; Hayward, N.J.; Chong, J.A.; Julius, D.; Moran, M.M.; et al. TRPA1 mediates formalin-induced pain. Proc. Natl. Acad. Sci. USA 2007, 104, 13525–13530. [Google Scholar]
- Tjolsen, A.; Berge, O.G.; Hunskaar, S.; Rosland, J.H.; Hole, K. The formalin test: An evaluation of the method. Pain 1992, 51, 5–17. [Google Scholar]
- Murray, C.W.; Porreca, F.; Cowan, A. Methodological refinements to the mouse paw formalin test. An animal model of tonic pain. J. Pharmacol. Methods 1988, 20, 175–186. [Google Scholar]
- Farias, J.A.; Ferro, J.N.; Silva, J.P.; Agra, I.K.; Oliveira, F.M.; Candea, A.L.; Conte, F.P.; Ferraris, F.K.; Henriques, M.D.; Conserva, L.M.; et al. Modulation of inflammatory processes by leaves extract from Clusia nemorosa both in vitro and in vivo animal models. Inflammation 2011. [Google Scholar] [CrossRef]
- Moore, A.R. Pleural models of inflammation: immune and nonimmune. Methods Mol. Biol 2003, 225, 123–128. [Google Scholar]
- Carbajal, D.; Molina, V.; Valdes, S.; Arruzazabala, M.L.; Mas, R.; Magraner, J. Anti-inflammatory activity of D-002: An active product isolated from beeswax. Prostaglandins Leukot. Essent. Fatty Acids 1998, 59, 235–238. [Google Scholar]
- Tsuji, R.F.; Hoshino, K.; Noro, Y.; Tsuji, N.M.; Kurokawa, T.; Masuda, T.; Akira, S.; Nowak, B. Suppression of allergic reaction by lambda-carrageenan: toll-like receptor 4/MyD88-dependent and -independent modulation of immunity. Clin. Exp. Allergy 2003, 33, 249–258. [Google Scholar]
- Utsunomiya, I.; Ito, M.; Oh-ishi, S. Generation of inflammatory cytokines in zymosan-induced pleurisy in rats: TNF induces IL-6 and cytokine-induced neutrophil chemoattractant (CINC) in vivo. Cytokine 1998, 10, 956–963. [Google Scholar]
- Saito, M.; Kinoshita, Y.; Satoh, I.; Shinbori, C.; Kono, T.; Hanada, T.; Uemasu, J.; Suzuki, H.; Yamada, M.; Satoh, K. N-hexacosanol ameliorates streptozotocin-induced diabetic rat nephropathy. Eur. J. Pharmacol 2006, 544, 132–137. [Google Scholar]
- Dixit, S.N.; Khosa, R.L. Chemical investigation of Tinospora cordifolia. Ind. J. Appl. Chem 1971, 34, 46–47. [Google Scholar]
- Morris, B.D.; Foster, S.P.; Harris, M.O. Identification of 1-octacosanal and 6-methoxy-2- benzoxazolinone from wheat as ovipositional stimulants for Hessian fly, Mayetiola destructor. J. Chem. Ecol 2000, 26, 859–873. [Google Scholar]
- Menendez, R.; Marrero, D.; Mas, R.; Fernandez, I.; Gonzalez, L.; Gonzalez, R.M. In vitro and in vivo study of octacosanol metabolism. Arch. Med. Res 2005, 36, 113–119. [Google Scholar]
- Eddy, N.B.; Leimbach, D. Synthetic analgesics. II. Dithienylbutenyl- and dithienylbutylamines. J. Pharmacol. Exp. Ther 1953, 107, 385–393. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups of animals | Total leukocytes (×106 cells/cavity) | Neutrophils (×106 cells/cavity) | Mononuclear cells (×106 cells/cavity) |
|---|---|---|---|
| Basal | 1.86 ± 0.14 | 0.12 ± 0.13 | 1.74 ± 0.16 |
| Cg | 11.85 ± 2.19 +++ | 8.05 ± 1.34 +++ | 3.22 ± 0.39 + |
| Cg + Octacosanol (1 mg/kg) | 5.70 ± 1.34 ** | 2.74 ± 0.53 *** | 2.89 ± 0.34 |
| Cg + Octacosanol (10 mg/kg) | 4.68 ± 0.51 *** | 2.22 ± 0.22 *** | 2.45 ± 0.36 |
| Cg + Indomethacin (20 mg/kg) | 3.26 ± 0.48 *** | 1.24 ± 0.18 *** | 1.83 ± 0.36 |
| Groups of animals | TNF-α (pg/mL) | Inhibition (%) |
|---|---|---|
| Basal | 19.53 ± 11.49 | - |
| Cg | 164.50 ± 50.99 ++ | - |
| Cg + Octacosanol (1 mg/kg) | 22.56 ± 14.80 ** | 86.28 |
| Cg + Octacosanol (10 mg/kg) | 24.17 ± 12.50 ** | 85.30 |
| Cg + Indomethacin (20 mg/kg) | 19.15 ± 4.43 ** | 88.35 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Oliveira, A.M.d.; Conserva, L.M.; De Souza Ferro, J.N.; Brito, F.d.A.; Lemos, R.P.L.; Barreto, E. Antinociceptive and Anti-Inflammatory Effects of Octacosanol from the Leaves of Sabicea grisea var. grisea in Mice. Int. J. Mol. Sci. 2012, 13, 1598-1611. https://doi.org/10.3390/ijms13021598
Oliveira AMd, Conserva LM, De Souza Ferro JN, Brito FdA, Lemos RPL, Barreto E. Antinociceptive and Anti-Inflammatory Effects of Octacosanol from the Leaves of Sabicea grisea var. grisea in Mice. International Journal of Molecular Sciences. 2012; 13(2):1598-1611. https://doi.org/10.3390/ijms13021598
Chicago/Turabian StyleOliveira, Anderson Marques de, Lucia M. Conserva, Jamylle N. De Souza Ferro, Fabíola de Almeida Brito, Rosângela P. Lyra Lemos, and Emiliano Barreto. 2012. "Antinociceptive and Anti-Inflammatory Effects of Octacosanol from the Leaves of Sabicea grisea var. grisea in Mice" International Journal of Molecular Sciences 13, no. 2: 1598-1611. https://doi.org/10.3390/ijms13021598