Analysis of Snail-1, E-Cadherin and Claudin-1 Expression in Colorectal Adenomas and Carcinomas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Snail-1
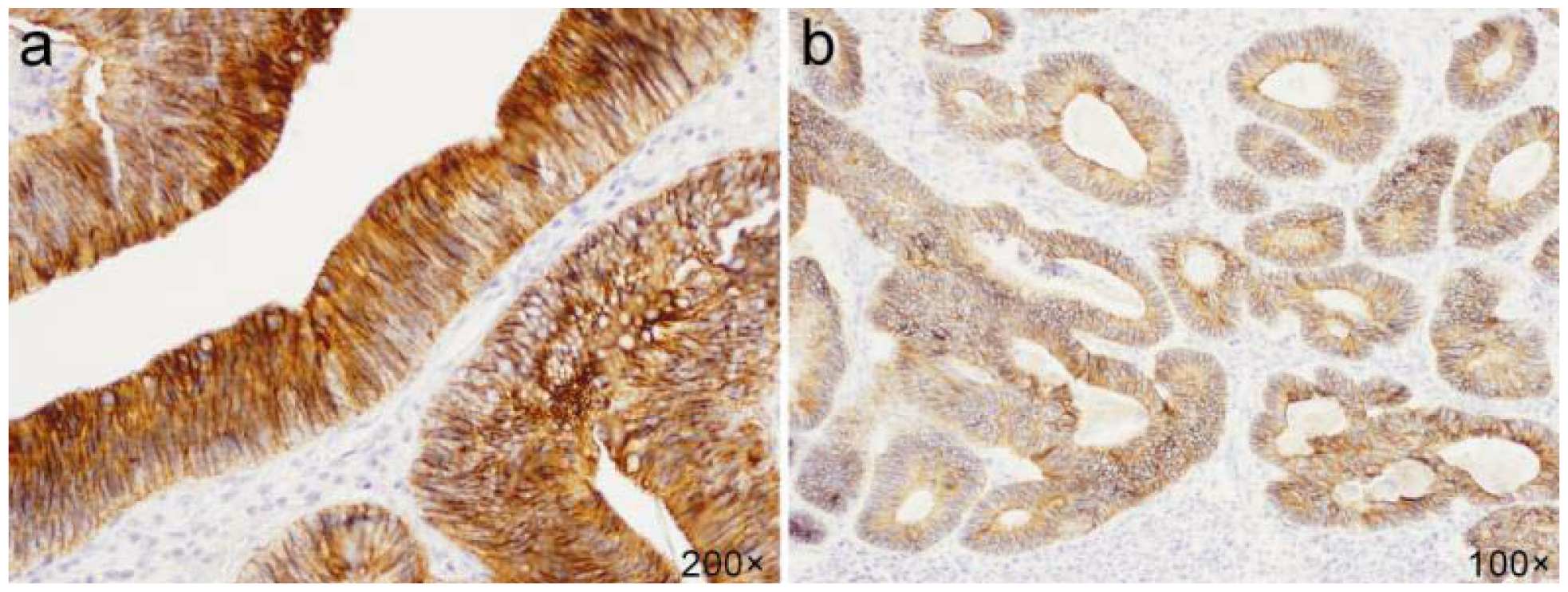
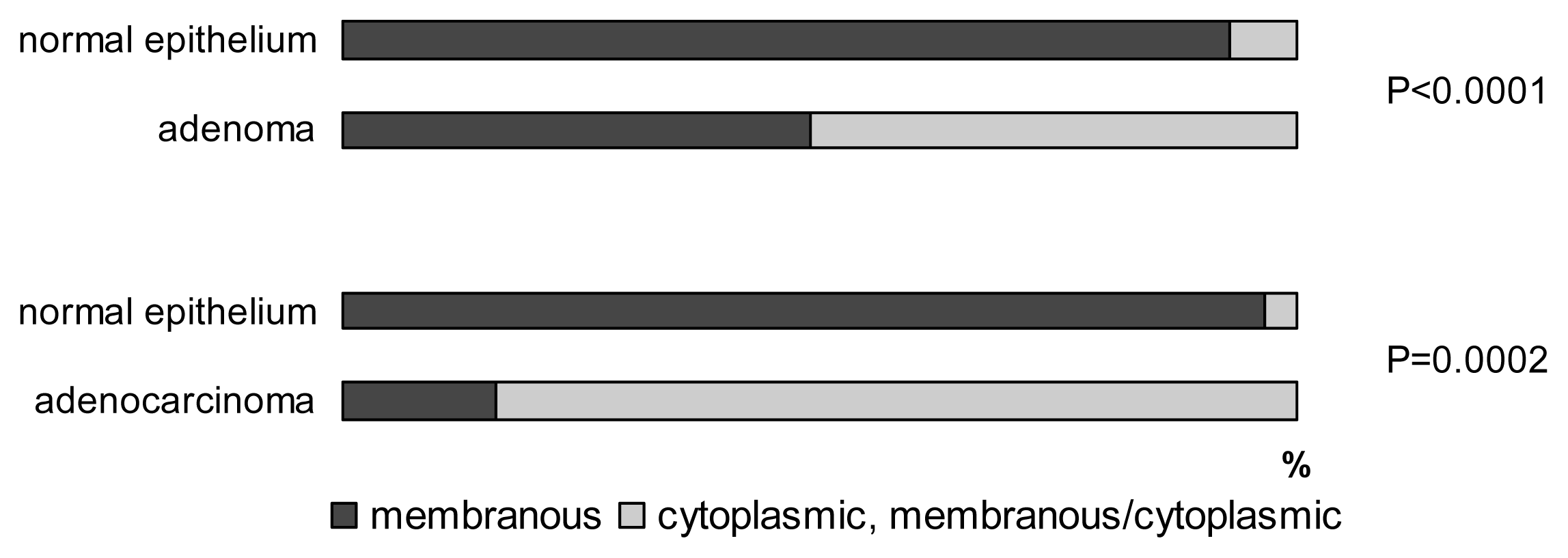
2.2. E-cadherin
2.3. Claudin-1
3. Discussion
4. Material and Methods
4.1. Immunohistochemistry
4.2. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Kreizenbeck, G.M.; Berger, A.J.; Subtil, A.; Rimm, D.L.; Gould Rothberg, B.E. Prognostic significance of cadherin-based adhesion molecules in cutaneous malignant melanoma. Cancer Epidemiol. Biomark. Prev 2008, 17, 949–958. [Google Scholar]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar]
- Yokoyama, K.; Kamata, N.; Hayashi, E.; Hoteiya, T.; Ueda, N.; Fujimoto, R.; Nagayama, M. Reverse correlation of E-cadherin and snail expression in oral squamous cell carcinoma cells in vitro. Oral Oncol 2001, 37, 65–71. [Google Scholar]
- Blanco, M.J.; Moreno-Bueno, G.; Sarrio, D.; Locascio, A.; Cano, A.; Palacios, J.; Nieto, M.A. Correlation of Snail expression with histological grade and lymph node status in breast carcinomas. Oncogene 2002, 21, 3241–3246. [Google Scholar]
- Jiao, W.; Miyazaki, K.; Kitajima, Y. Inverse correlation between E-cadherin and Snail expression in hepatocellular carcinoma cell lines in vitro and in vivo. Br. J. Cancer 2002, 86, 98–101. [Google Scholar]
- Rosivatz, E.; Becker, I.; Specht, K.; Fricke, E.; Luber, B.; Busch, R.; Höfler, H.; Becker, K.F. Differential expression of the epithelial-mesenchymal transition regulators snail, SIP1, and twist in gastric cancer. Am. J. Pathol 2002, 161, 1881–1891. [Google Scholar]
- Poser, I.; Domínguez, D.; de Herreros, A.G.; Varnai, A.; Buettner, R.; Bosserhoff, A.K. Loss of E-cadherin expression in melanoma cells involves up-regulation of the transcriptional repressor Snail. J. Biol. Chem 2001, 276, 24661–24666. [Google Scholar]
- Francí, C.; Gallén, M.; Alameda, F.; Baró, T.; Iglesias, M.; Virtanen, I.; García de Herreros, A. Snail1 protein in the stroma as a new putative prognosis marker for colon tumours. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar]
- Batlle, E.; Sancho, E.; Francí, C.; Domínguez, D.; Monfar, M.; Baulida, J.; García de Herreros, A. The transcription factor snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nat. Cell Biol 2000, 2, 84–89. [Google Scholar]
- Cano, A.; Pérez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; del Barrio, M.G.; Portillo, F.; Nieto, M.A. The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat. Cell Biol 2000, 2, 76–83. [Google Scholar]
- Hemavathy, K.; Ashraf, S.I.; Ip, Y.T. Snail/slug family of repressors: Slowly going into the fast lane of development and cancer. Gene 2000, 257, 1–12. [Google Scholar]
- Strumane, K.; van Roy, F.; Berx, G. The Role of E-Cadherin in Epithelial Differentiation and Cance Progression. In Recent Research Development in Cellular Biochemistry; Pandalai, S.G., Ed.; Transworld Research Network: Kerala, India, 2003; pp. 33–77. [Google Scholar]
- Miwa, N.; Furuse, M.; Tsukita, S.; Niikawa, N.; Nakamura, Y.; Furukawa, Y. Involvement of claudin-1 in the beta-catenin/Tcf signaling pathway and its frequent upregulation in human colorectal cancers. Oncol. Res 2001, 12, 469–476. [Google Scholar]
- Dhawan, P.; Singh, A.B.; Deane, N.G.; No, Y.; Shiou, S.R.; Schmidt, C.; Neff, J.; Washington, M.K.; Beauchamp, R.D. Claudin-1 regulates cellular transformation and metastatic behavior in colon cancer. J. Clin. Invest 2005, 115, 1765–1776. [Google Scholar]
- Resnick, M.B.; Konkin, T.; Routhier, J.; Sabo, E.; Pricolo, V.E. Claudin-1 is a strong prognostic indicator in stage II colonic cancer: A tissue microarray study. Mod. Pathol 2005, 18, 511–518. [Google Scholar]
- Wu, Y.; Zhou, B.P. TNF-α/NF-κB/Snail pathway in cancer cell migration and invasion. Br. J. Cancer 2010, 102, 639–644. [Google Scholar]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar]
- Roy, H.K.; Iversen, P.; Hart, J.; Liu, Y.; Koetsier, J.L.; Kim, Y.; Kunte, D.P.; Madugula, M.; Backman, V.; Wali, R.K. Down-regulation of SNAIL suppresses MIN mouse tumorigenesis: Modulation of apoptosis, proliferation, and fractal dimension. Mol. Cancer Ther 2004, 3, 1159–1165. [Google Scholar]
- Massoumi, R.; Kuphal, S.; Hellerbrand, C.; Haas, B.; Wild, P.; Spruss, T.; Pfeifer, A.; Fässler, R.; Bosserhoff, A.K. Down-regulation of SNAIL suppresses MIN mouse tumorigenesis: Modulation of apoptosis, proliferation, and fractal dimension. J. Exp. Med 2009, 206, 221–232. [Google Scholar]
- Wang, Y.; Zhou, B.P. Epithelial-mesenchymal transition in breast cancer progression and metastasis. Chin. J. Cancer 2011, 30, 603–611. [Google Scholar]
- Zhu, Y.; Nilsson, M.; Sundfeldt, K. Phenotypic plasticity of the ovarian surface epithelium: TGF-beta 1 induction of epithelial to mesenchymal transition (EMT) in vitro. Endocrinology 2010, 151, 5497–5505. [Google Scholar]
- Domínguez, D.; Montserrat-Sentís, B.; Virgós-Soler, A.; Guaita, S.; Grueso, J.; Porta, M.; Puig, I.; Baulida, J.; Francí, C.; García de Herreros, A. Phosphorylation regulates the subcellular location and activity of the snail transcriptional repressor. Mol. Cell Biol 2003, 23, 5078–5089. [Google Scholar]
- Kaler, P.; Galea, V.; Augenlicht, L.; Klampfer, L. Tumor associated macrophages protect colon cancer cells from TRAIL-induced apoptosis through IL-1beta-dependent stabilization of Snail in tumor cells. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Kudo-Saito, C.; Shirako, H.; Takeuchi, T.; Kawakami, Y. Cancer metastasis is accelerated through immunosuppression during Snail-induced EMT of cancer cells. Cancer Cell 2009, 15, 195–206. [Google Scholar]
- Weis, W.I.; Nelson, W.J. Re-solving the cadherin-catenin-actin conundrum. J. Biol. Chem 2006, 281, 35593–35597. [Google Scholar]
- Perl, A.K.; Wilgenbus, P.; Dahl, U.; Semb, H.; Christofori, G. A causal role for E-cadherin in the transition from adenoma to carcinoma. Nature 1998, 392, 190–193. [Google Scholar]
- Stemmer, V.; de Craene, B.; Berx, G.; Behrens, J. Snail promotes Wnt target gene expression and interacts with beta-catenin. Oncogene 2008, 27, 5075–5080. [Google Scholar]
- Ikenouchi, J.; Matsuda, M.; Furuse, M.; Tsukita, S. Regulation of tight junctions during the epithelium-mesenchyme transition: Direct repression of the gene expression of claudins/occludin by Snail. J. Cell Sci 2003, 116, 1959–1967. [Google Scholar]
- de Craene, B.; Gilbert, B.; Stove, C.; Bruyneel, E.; van Roy, F.; Berx, G. The transcription factor snail induces tumor cell invasion through modulation of the epithelial cell differentiation program. Cancer Res 2005, 65, 6237–6244. [Google Scholar]
- Gröne, J.; Weber, B.; Staub, E.; Heinze, M.; Klaman, I.; Pilarsky, C.; Hermann, K.; Castanos-Velez, E.; Röpcke, S.; Mann, B.; et al. Differential expression of genes encoding tight junction proteins in colorectal cancer: Frequent dysregulation of claudin-1, -8 and -12. Int. J. Colorectal. Dis 2007, 22, 651–659. [Google Scholar]
- Shiou, S.R.; Singh, A.B.; Moorthy, K.; Datta, P.K.; Washington, M.K.; Beauchamp, R.D.; Dhawan, P. Smad4 regulates claudin-1 expression in a transforming growth factor-beta-independent manner in colon cancer cells. Cancer Res 2007, 67, 1571–1579. [Google Scholar]
- French, A.D.; Fiori, J.L.; Camilli, T.C.; Leotlela, P.D.; O’Connell, M.P.; Frank, B.P.; Subaran, S.; Indig, F.E.; Taub, D.D.; Weeraratna, A.T. PKC and PKA phosphorylation affect the subcellular localization of claudin-1 in melanoma cells. Int. J. Med. Sci 2009, 6, 93–101. [Google Scholar]
- Simet, S.M.; Wyatt, T.A.; Devasure, J.; Yanov, D.; Allen-Gipson, D.; Sisson, J.H. Alcohol increases the permeability of airway epithelial tight junctions in beas-2B and NHBE cells. Alcohol. Clin. Exp. Res 2011. [Google Scholar] [CrossRef]
- Mandel, I.; Paperna, T.; Glass-Marmor, L.; Volkowich, A.; Badarny, S.; Schwartz, I.; Vardi, P.; Koren, I.; Miller, A. Tight junction proteins expression and modulation in immune cells and multiple sclerosis. J. Cell Mol. Med 2011. [Google Scholar] [CrossRef]
- Cohn, M.L.; Goncharuk, V.N.; Diwan, A.H.; Zhang, P.S.; Shen, S.S.; Prieto, V.G. Loss of claudin-1 expression in tumor-associated vessels correlates with acquisition of metastatic phenotype in melanocytic neoplasms. J. Cutan. Pathol 2005, 32, 533–536. [Google Scholar]
- Kaarteenaho-Wiik, R.; Soini, Y. Claudin-1, -2, -3, -4, -5, and -7 in usual interstitial pneumonia and sarcoidosis. J. Histochem. Cytochem 2009, 57, 187–195. [Google Scholar]





© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bezdekova, M.; Brychtova, S.; Sedlakova, E.; Langova, K.; Brychta, T.; Belej, K. Analysis of Snail-1, E-Cadherin and Claudin-1 Expression in Colorectal Adenomas and Carcinomas. Int. J. Mol. Sci. 2012, 13, 1632-1643. https://doi.org/10.3390/ijms13021632
Bezdekova M, Brychtova S, Sedlakova E, Langova K, Brychta T, Belej K. Analysis of Snail-1, E-Cadherin and Claudin-1 Expression in Colorectal Adenomas and Carcinomas. International Journal of Molecular Sciences. 2012; 13(2):1632-1643. https://doi.org/10.3390/ijms13021632
Chicago/Turabian StyleBezdekova, Michala, Svetlana Brychtova, Eva Sedlakova, Katerina Langova, Tomas Brychta, and Kamil Belej. 2012. "Analysis of Snail-1, E-Cadherin and Claudin-1 Expression in Colorectal Adenomas and Carcinomas" International Journal of Molecular Sciences 13, no. 2: 1632-1643. https://doi.org/10.3390/ijms13021632