Identifying Differentially Expressed Genes in Pollen from Self-Incompatible “Wuzishatangju” and Self-Compatible “Shatangju” Mandarins

Abstract
:1. Introduction
2. Results
2.1. Identification of Gene Fragments from Two SSH Libraries
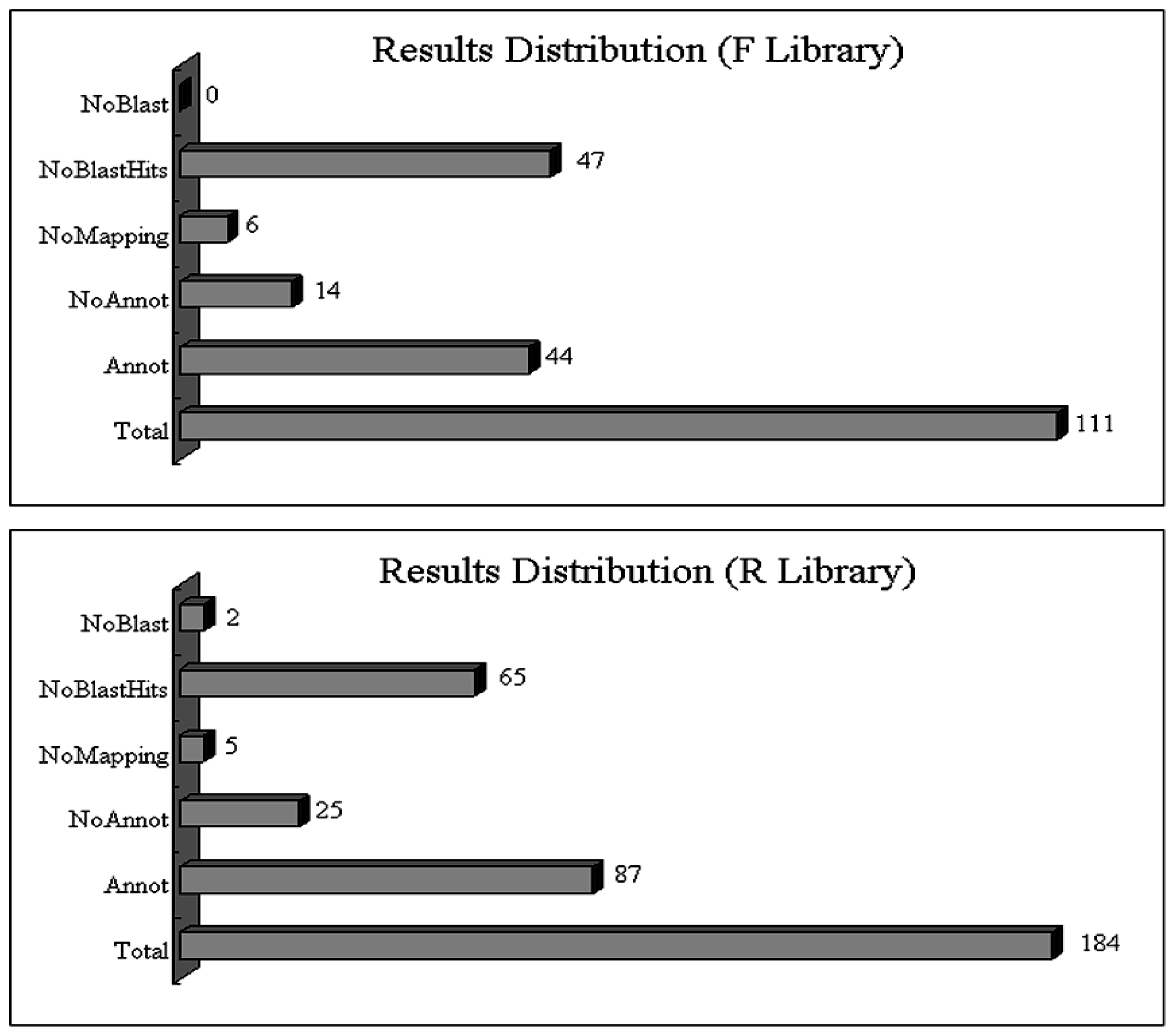
2.2. General Statistics of Two SSH Libraries
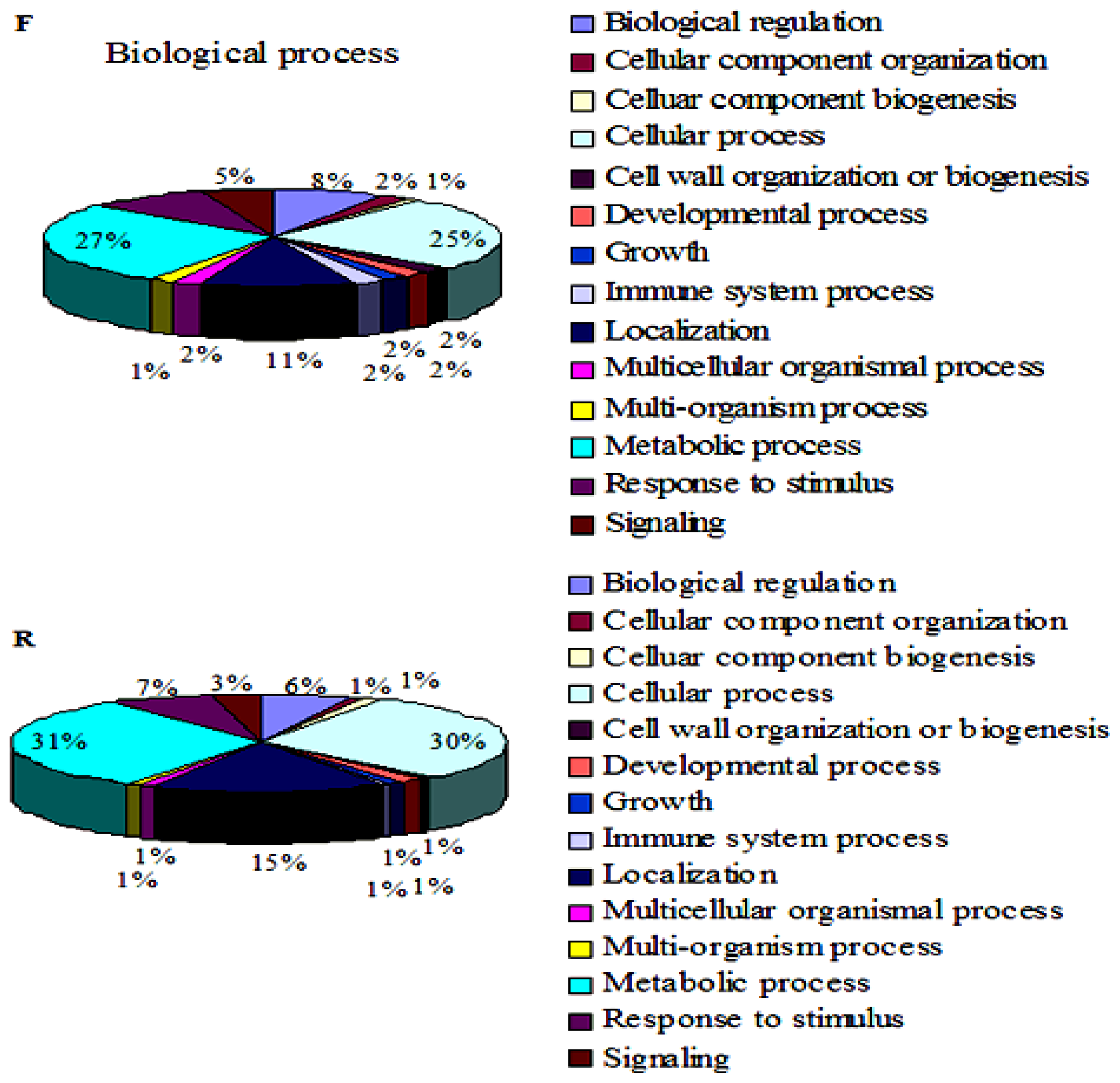
2.3. Gene Ontology Analysis
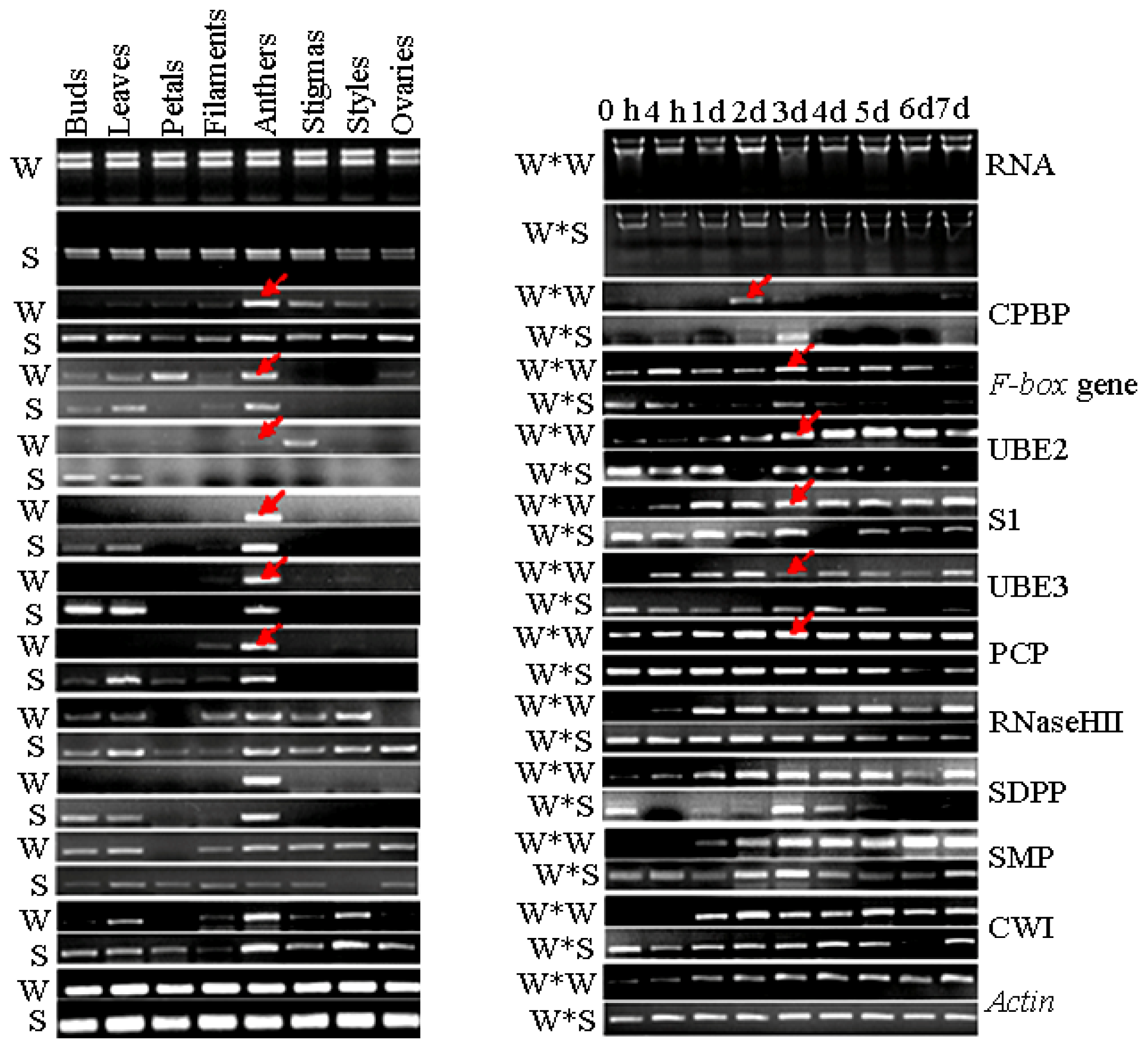
2.4. Expression Analyses of SI Candidate Genes by SqRT-PCR
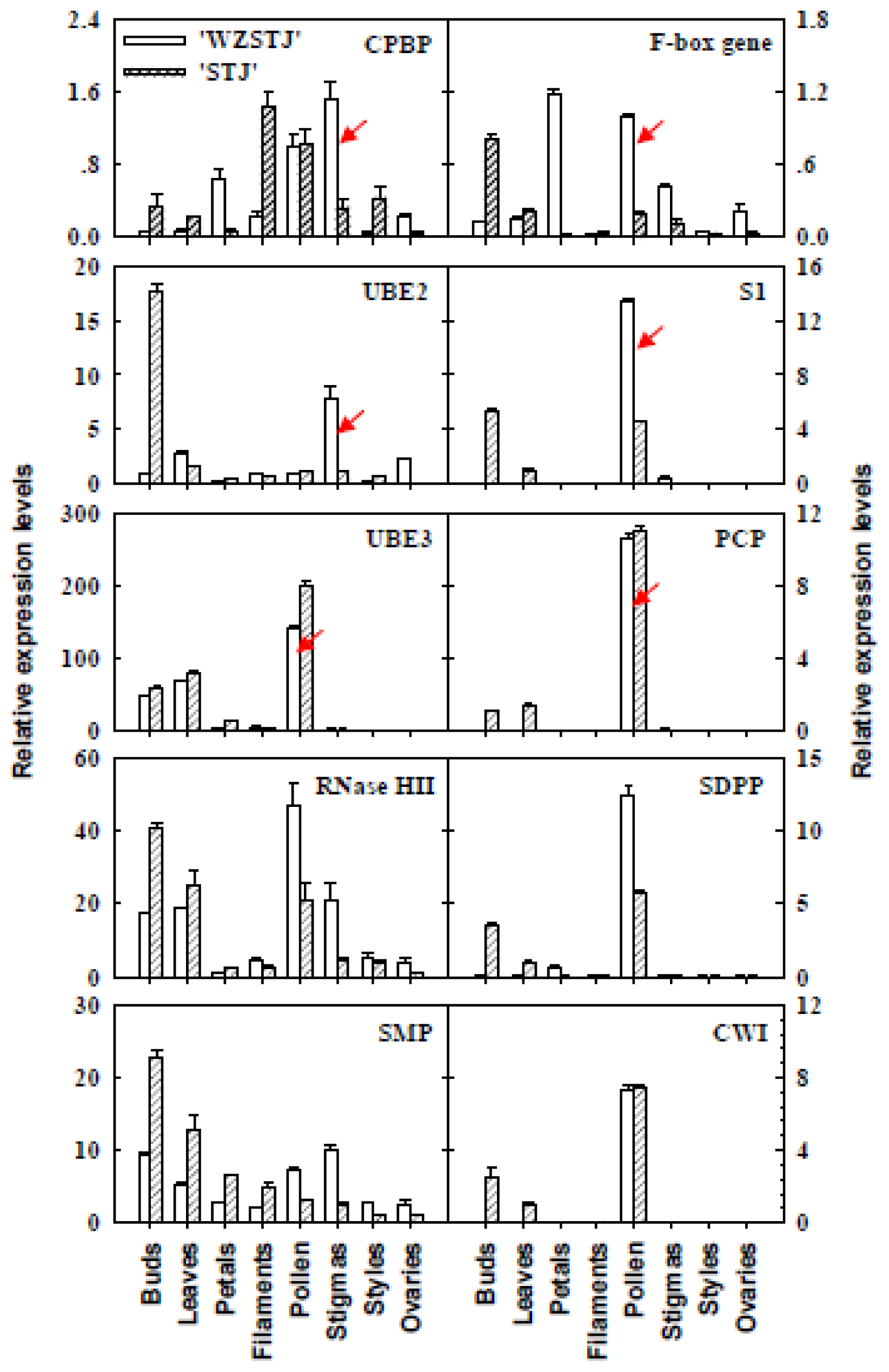
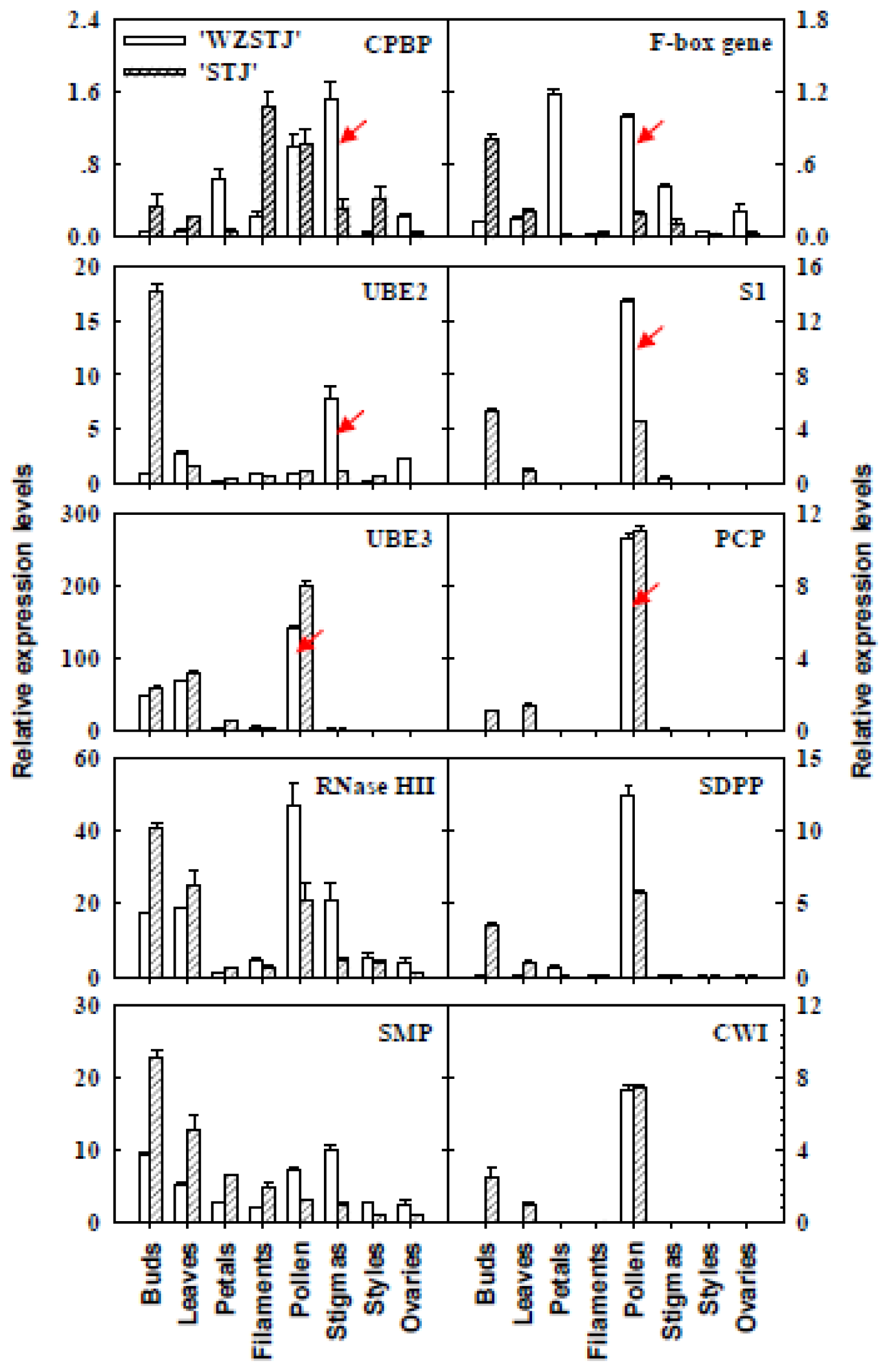
2.5. Expression Analyses of SI-Related Genes in Different Organs from “Wuzishatangju” and “Shatangju” Mandarins Using qPCR
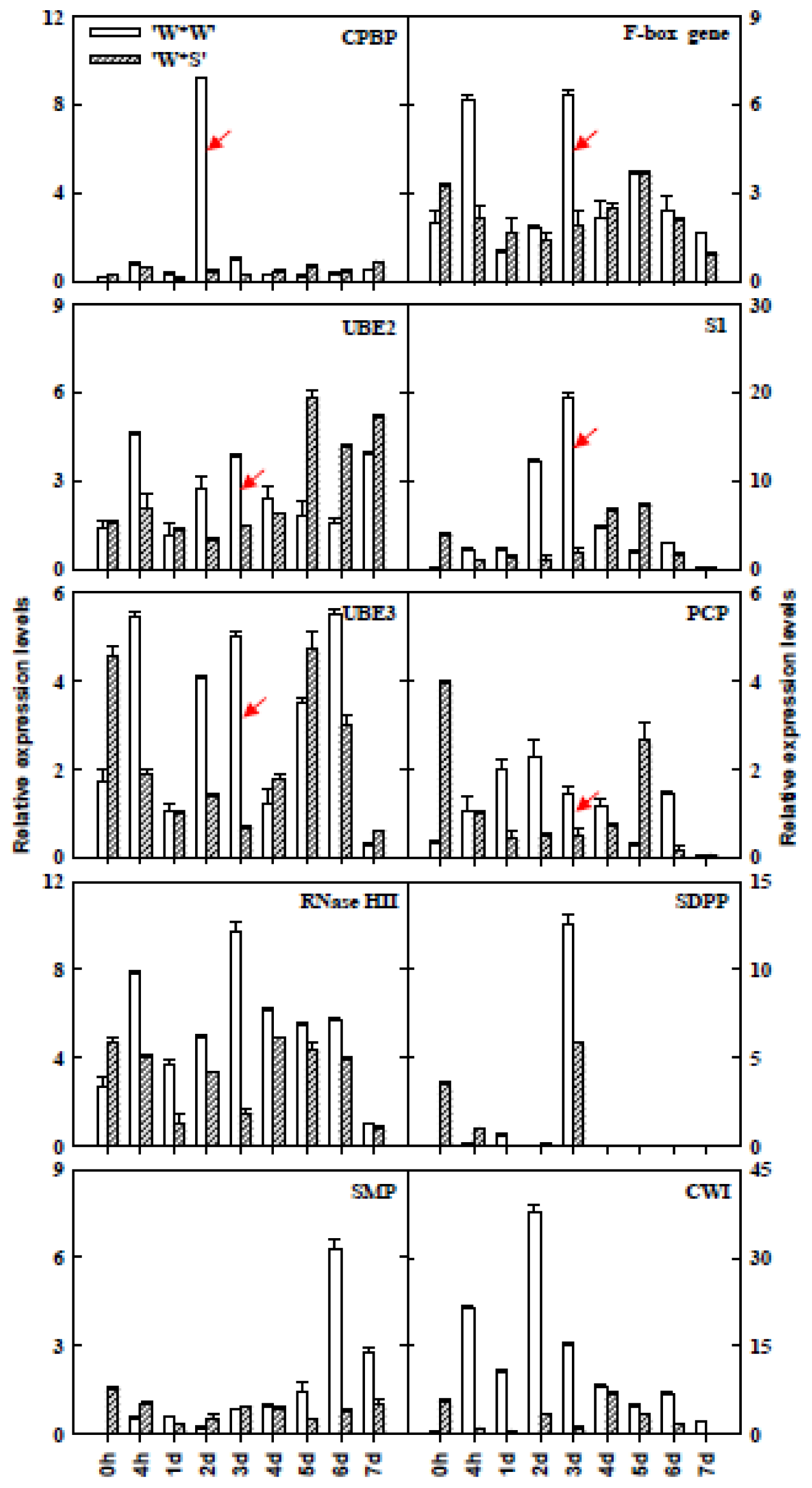
2.6. Expression Analyses of SI-Related Genes in Different Stages of the Pistil after Self-Pollination of “Wuzishatangju” and Cross-Pollination of “Wuzishatangju” × “Shatangju” Using qPCR
3. Discussion
4. Experimental Section
4.1. Plant Materials
4.2. Total RNA Extraction and mRNA Purification
4.3. Construction of SSH Libraries
4.4. Screening of SSH Libraries Using Colony-PCR and Reverse Northern Analysis
4.5. Bioinformatics Analysis of Expressed Sequence Tags (ESTs)
4.6. Expression Analysis of SI-Related Genes by SqRT-PCR and qPCR
5. Conclusion
Acknowledgements
Conflict of Interest
References
- De Nettancourt, D. Incompatibility in angiosperms. Sex Plant Reprod 1997, 10, 185–199. [Google Scholar]
- Takayama, S.; Shimosato, H.; Shiba, H.; Funato, M.; Che, F.S.; Watanabe, M.; Iwano, M.; Isogai, A. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature 2001, 413, 534–538. [Google Scholar]
- McClure, B.; Cruz-García, F.; Romero, C. Compatibility and incompatibility in S-RNase-based systems. Ann. Bot 2011, 108, 647–658. [Google Scholar]
- Lai, Z.; Ma, W.; Han, B.; Liang, L.; Zhang, Y.; Hong, G.; Xue, Y. An F-box gene linked to the self-incompatibility (S) locus of Antirrhinum is expressed specifically in pollen and tapetum. Plant Mol. Biol 2002, 50, 29–41. [Google Scholar]
- Ushijima, K.; Sassa, H.; Dandekar, A.M.; Gradziel, T.M.; Tao, R.; Hirano, H. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. Plant Cell 2003, 15, 771–781. [Google Scholar]
- Kubo, K.; Entani, T.; Takara, A.; Wang, N.; Fields, A.M.; Hua, Z.; Toyoda, M.; Kawashima, S.; Ando, T.; Isogai, A.; et al. Collaborative non-self recognition system in S-RNase-based self-incompatibility. Science 2010, 330, 796–799. [Google Scholar]
- Meng, X.; Sun, P.; Kao, T.H. S-RNase-based self-incompatibility in Petunia inflata. Ann. Bot 2010, 253, 1–10. [Google Scholar]
- Tao, R.; Iezzoni, A.F. The S-RNase-based gametophytic self-incompatibility system in Prunus exhibits distinct genetic and molecular features. Sci. Hortic 2010, 124, 423–433. [Google Scholar]
- Liang, L.; Huang, J.; Xue, Y. Identification and evolutionary analysis of a relic S-RNase in Antirrhinum. Sex Plant Reprod 2003, 16, 17–22. [Google Scholar]
- Li, J.; Nass, N.; Kusaba, M.; Dodds, P.N.; Treloar, N.; Clarke, A.E.; Newbigin, E. A genetic map of the Nicotiana alata S locus that includes three pollen-expressed genes. Theor. Appl. Genet 2000, 100, 956–964. [Google Scholar]
- Sijacic, P.; Wang, X.; Skirpan, A.L.; Wang, Y.; Dowd, P.E.; McCubbin, A.G.; Huang, S.; Kao, T.H. Identification of the pollen determinant of S-RNase-mediated self-incompatibility. Nature 2004, 429, 302–305. [Google Scholar]
- Entani, T.; Iwano, M.; Shiba, H.; Che, F.S.; Isogai, A.; Takayama, S. Comparative analysis of the self-incompatibility (S-) locus region of Prunus mume: Identification of a pollen-expressed F-box gene with allelic diversity. Genes Cells 2003, 8, 203–213. [Google Scholar]
- Ushijima, K.; Yamane, H.; Watari, A.; Kakehi, E.; Ikeda, K.; Hauck, N.R.; Iezzoni, A.F.; Tao, R. The S haplotype-specific F-box protein gene, SFB, is defective in self-compatible haplotypes of Prunus avium and P. mume. Plant J 2004, 39, 573–586. [Google Scholar]
- Vilanova, S.; Badenes, M.L.; Burgos, L.; Martínez-Calvo, J.; Llácer, G.; Romero, C. Self-compatibility of two apricot selections is associated with two pollen-part mutations of different nature. Plant Physiol 2006, 142, 629–641. [Google Scholar]
- Sonneveld, T.; Tobutt, K.R.; Vaughan, S.P.; Robbins, T.P. Loss of pollen-S function in two self-compatible selections of Prunus avium is associated with deletion/mutation of an S haplotype-specific F-box gene. Plant Cell 2005, 17, 37–51. [Google Scholar]
- Hauck, N.R.; Yamane, H.; Tao, R.; Iezzoni, A.F. Accumulation of nonfunctional S-haplotypes results in the breakdown of gametophytic self-incompatibility in tetraploid prunus. Genetics 2006, 172, 1191–1198. [Google Scholar]
- Cheng, J.; Han, Z.; Xu, X.; Li, T. Isolation and identification of the pollen-expressed polymorphic F-box genes linked to the S-locus in apple (Malus × domestica). Sex Plant Reprod 2006, 19, 175–183. [Google Scholar]
- Fields, A.M.; Wang, N.; Hua, Z.; Meng, X.; Kao, T.H. Functional characterization of two chimeric proteins between a Petunia inflata S-locus F-box protein, PiSLF2, and a PiSLF-like protein, PiSLFb-S2. Plant Mol. Biol 2010, 74, 279–292. [Google Scholar]
- Okada, K.; Tonaka, N.; Taguchi, T.; Ichikawa, T.; Sawamura, Y.; Nakanishi, T.; Takasaki-Yasuda, T. Related polymorphic F-box protein genes between haplotypes clustering in the BAC contig sequences around the S-RNase of Japanese pear. J. Exp. Bot 2011, 62, 1887–1902. [Google Scholar]
- Soost, R.K. Self-incompatibility in Citrus grandis Osbeck. J. Am. Soc. Hortic. Sci 1964, 84, 137–140. [Google Scholar]
- Distefano, G.; Caruso, M.; Malfa, S.L.; Gentile, A.; Tribulato, E. Histological and molecular analysis of pollen-pistil interaction in clementine. Plant Cell Rep 2009, 28, 1439–1451. [Google Scholar]
- Ngo, B.X.; Wakana, A.; Kim, J.H.; Mori, T.; Sakai, K. Estimation of self-incompatibility S genotypes of Citrus cultivars and plants based on controlled pollination with restricted number of pollen grains. J. Fac. Agr. Kyushu Univ 2010, 55, 67–72. [Google Scholar]
- Wang, P.; Lü, L. Self-incompatible reaction parts in Citrus grandis “Guanximiyou” and “Duweimiyou”—Observation of pollination on different pistil parts in vitro. Chin. J. Trop. Crops 2009, 30, 1105–1108. [Google Scholar]
- Ye, W.; Qin, Y.; Ye, Z.; Teixeira da Silva, J.A.; Zhang, L.; Wu, X.; Lin, S.; Hu, G. Seedless mechanism of a new mandarin cultivar “Wuzishatangju” (Citrus reticulata Blanco). Plant Sci 2009, 177, 19–27. [Google Scholar]
- Roiz, L.; Goren, R.; Shoseyov, O. Stigmatic RNase in calamondin (Citrus reticulata var. austera × Fortunella sp.). Physiol. Plantarum 1995, 94, 585–590. [Google Scholar]
- Kim, J.H.; Mori, T.; Wakana, A.; Ngo, B.X.; Masuda, J.; Sakai, K.; Kajiwara, K. Production of homozygous S1 seedlings for S gene in “Hirado Buntan” pummelo (Citrus grandis Osbeck) and determination of the S alleles (S9 and S10) by pollination with the S1 seedlings to Citrus cultivars. J. Fac. Agr. Kyushu U 2010, 55, 239–245. [Google Scholar]
- Chai, L.; Ge, X.; Xu, Q.; Deng, X. CgSL2, an S-like RNase gene in “Zigui shatian” pummelo (Citrus grandis Osbeck), is involved in ovary senescence. Mol. Biol. Rep 2011, 38, 1–8. [Google Scholar]
- Miao, H.; Qin, Y.; Teixeira da Silva, J.A.; Ye, Z.; Hu, G. Cloning and expression analysis of S-RNase homologous gene in Citrus reticulata Blanco cv. Wuzishatangju. Plant Sci 2011, 180, 358–367. [Google Scholar]
- Miao, H.; Qin, Y.; Ye, Z.; Hu, G. Molecular characterization and expression analysis of ubiquitin-activating enzyme E1 gene in Citrus reticulate. Gene 2013, 513, 249–259. [Google Scholar]
- Miao, H.; Qin, Y.; Ye, Z.; Hu, G. Molecular characterization and expression analysis of S1 self-incompatibility locus-linked pollen 3.15gene inCitrus reticulata. J. Integr. Plant Biol. 2013. [Google Scholar] [CrossRef]
- Miao, H.; Qin, Y.; Teixeira da Silva, J.A.; Ye, Z.; Hu, G. Identification of differentially expressed genes in pistils from self-incompatible Citrus reticulata by suppression subtractive hybridization. Mol. Biol. Rep 2013, 40, 159–169. [Google Scholar]
- Miao, H.; Qin, Y.; Teixeira da Silva, J.A.; Ye, Z.; Hu, G. Isolation and differential expression analysis of self-compatibility-related genes from mature pistils of “Shatangju” mandarin (Citrus reticulata Blanco). J. Hortic. Sci. Biotech 2011, 86, 575–582. [Google Scholar]
- Yang, B.; Thorogood, D.; Armstead, I.P.; Franklin, F.C.H.; Barth, S. Identification of genes expressed during the self-incompatibility response in perennial ryegrass (Lolium perenne L.). Plant Mol. Biol 2009, 70, 709–723. [Google Scholar]
- Allen, A.M.; Thorogood, C.J.; Hegarty, M.J.; Lexer, C.; Hiscock, S.J. Pollen-pistil interactions and self-incompatibility in the Asteraceae: New insights from studies of Senecio squalidus (Oxford ragwort). Ann. Bot 2011, 108, 687–698. [Google Scholar]
- Wheeler, D.; Newbigin, E. Expression of 10 S-class SLF-like genes in Nicotiana alata pollen and its implications for understanding the pollen factor of the S locus. Genetics 2007, 177, 2171–2180. [Google Scholar]
- Chai, L.; Ge, X.; Biswas, M.K.; Deng, X. Molecular analysis and expression of a floral organ-relative F-box gene isolated from “Zigui shatian” pummelo (Citrus grandis Osbeck). Mol. Biol. Rep 2011, 38, 4429–4436. [Google Scholar]
- McClure, B. Darwin’s foundation for investigating self-incompatibility and the progress toward a physiological model for S-RNase-based SI. J. Exp. Bot 2009, 60, 1069–1081. [Google Scholar]
- Caruso, M.; Merelo, P.; Distefano, G.; La Malfa, S.; Lo Piero, A.R.; Tadeo, F.R.; Talon, M.; Gentile, A. Comparative transcriptome analysis of stylar canal cells identifies novel candidate genes implicated in the self-incompatibility response ofCitrus clementina. BMC Plant Biol. 2012, 12. [Google Scholar] [CrossRef]
- Foote, H.C.; Ride, J.P.; Franklin-Tong, V.E.; Walker, E.A.; Lawrence, M.J.; Franklin, F.C. Cloning and expression of a distinctive class of self-incompatibility (S) gene from Papaver rhoeas L. Proc. Natl. Acad. Sci. USA 1994, 91, 2265–2269. [Google Scholar]
- Moon, J.; Parry, G.; Estelle, M. The ubiquitin-proteasome pathway and plant development. Plant Cell 2004, 16, 3181–3195. [Google Scholar]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci 2006, 31, 137–155. [Google Scholar]
- Stone, S.L.; Anderson, E.M.; Mullen, R.T.; Goring, D.R. ARC1 is an E3 ubiquitin ligase and promotes the ubiquitination of proteins during the rejection of self-incompatible Brassica pollen. Plant Cell 2003, 15, 885–898. [Google Scholar]
- Qiao, H.; Wang, F.; Zhao, L.; Zhou, J.; Lai, Z.; Zhang, Y.; Robbins, T.P.; Xue, Y. The F-box protein AhSLF-S2 controls the pollen function of S-RNase-based self-incompatibility. Plant Cell 2004, 16, 2307–2322. [Google Scholar]
- Franklin-Tong, V.E.; Hackett, G.; Hepler, P.K. Ratio-imaging of Ca2+ i in the self-incompatibility response in pollen tubes of Papaver rhoeas. Plant J 1997, 12, 1375–1386. [Google Scholar]
- Kunz, C.; Chang, A.; Faure, J.; Clarke, A.E.; Polya, G.M.; Anderson, M.A. Phosphorylation of style S-RNase by Ca2+-dependent protein kinases from pollen tubes. Sex Plant Reprod 1996, 9, 25–34. [Google Scholar]
- Myers, C.; Romanowsky, S.M.; Barron, Y.D.; Garg, S.; Azuse, C.L.; Curran, A.; Davis, R.M.; Hatton, J.; Harmon, A.C.; Harper, J.F. Calcium-dependent protein kinases regulate polarized tip growth in pollen tubes. Plant J 2009, 59, 528–539. [Google Scholar]
- Roberts, D.M.; Harmon, A.C. Calcium-modulated proteins: Targets of intracellular calcium signals in higher plants. Annu. Rev. Plant Biol 1992, 43, 375–414. [Google Scholar]
- Yang, W.; Lai, Y.; Li, M.; Xu, W.; Xue, Y. A novel C2-domain phospholipid-bind protein, OsPBP1, is required for pollen fertility in rice. Mol. Plant 2008, 1, 770–785. [Google Scholar]
- Doughty, J.; Dixon, S.; Hiscock, S.J.; Willis, A.C.; Parkin, I.A.; Dickinson, H.G. PCP-A1, a defensin-like Brassica pollen coat protein that binds the S locus glycoprotein, is the product of gametophytic gene expression. Plant Cell 1998, 10, 1333–1347. [Google Scholar]
- Shiba, H.; Takayama, S.; Iwano, M.; Shimosato, H.; Funato, M.; Nakagawa, T.; Che, F.S.; Suzuki, G.; Watanabe, M.; Hinata, K.; et al. A pollen coat protein, SP11/SCR, determines the pollen S-specificity in the self-incompatibility of Brassica species. Plant Physiol 2001, 125, 2095–2103. [Google Scholar]
- Itaya, M. Isolation and characterization of a second RNase H (RNase HII) of Escherichia coli K-12 encoded by the rnhB gene. Proc. Natl. Acad. Sci. USA 1990, 87, 8587–8591. [Google Scholar]
- Bracale, M.; Galli, M.G.; Falavigna, A.; Soave, C. Sexual differentiation in Asparagus officinalisL. Sex Plant Reprod. 1990, 3, 23–30. [Google Scholar]
- Engelke, T.; Hirsche, J.; Roitsch, T. Anther-specific carbohydrate supply and restoration of metabolically engineered male sterility. J. Exp. Bot 2010, 61, 2693–2706. [Google Scholar]
- Lee, P.; Chow, T.; Chen, Z.; Hsing, Y. Genomic nucleotide sequence of a soybean seed maturation protein GmPM9 gene. Plant Physiol 1992, 100, 2121–2122. [Google Scholar]
- Luo, M.; Xiao, Y.; Hou, L.; Luo, X.; Li, D.; Pei, Y. Cloning and expression analysis of a LIM-domain protein gene from cotton (Gossypium hirsutum L.). Acta Genetica Sin 2003, 30, 175–182. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seq. Name | Seq. Description | GenBank accession No. | Length (bp) | E-value | Similarity | Annotation |
|---|---|---|---|---|---|---|
| F library | ||||||
| H102 | Pollen coat-like protein (PCP) | JK724775 | 488 | 2.23 × 10−31 | 86.55% | Pollen development |
| H115 | Calcium-dependent phospholipid binding protein (CPBP) | JK724776 | 805 | 3.67 × 10−112 | 86.40% | Calcium ion binding |
| H128 | Pollen allergen Pla o1 | JK724777 | 526 | 1.80 × 10−35 | 57.85% | Extracellular space |
| H264 | Senescence-associated protein | JK724778 | 252 | 1.07 × 10−36 | 89.95% | Biological process |
| H304 | F-box protein (F-box) | JK724779 | 446 | 3.04 × 10−23 | 87.80% | Kinase activity |
| H428 | Actin depolymerizing factor (ADF) | JK724780 | 612 | 6.19 × 10−85 | 93.75% | Cytoskeleton |
| H543 | Zinc finger protein | JK724781 | 372 | 4.58 × 10−33 | 69.80% | Transcription factor activity |
| H556 | Ubiquitin-protein ligase E3 (UBE3) | JK724782 | 471 | 1.44 × 10−62 | 95.00% | CUL4 RING ubiquitin ligase |
| H858 | Ubiquitin-conjugating enzyme E2 (UBE2) | JK724783 | 314 | 1.58 × 10−35 | 97.63% | ATP binding |
| H1092 | Vacuolar H+-translocating inorganic pyrophosphatase | JK724784 | 808 | 1.33 × 10−5 | 64.56% | Inorganic diphosphatase activity |
| R library | ||||||
| F15 | Phospholipase | JK724785 | 248 | 7.28 × 10−39 | 77.30% | Serine/threonine kinase activity |
| F65 | F-box and wd40 domain protein | JK724786 | 339 | 6.79 × 10−52 | 82.15% | G-protein complex |
| F78 | Self-incompatibility S1 family protein (S1) | JK724787 | 252 | 1.38 × 10−9 | 62.83% | Pollen-pistil interaction |
| F960 | Sexual differentiation process protein isp4-like (SDPP) | JK724788 | 840 | 7.23 × 10−67 | 83.80% | Transporter activity |
| F966 | Seed maturation protein PM37 (SMP) | JK724789 | 246 | 5.47 × 10−26 | 87.90% | Biological process |
| F1166 | Ca2+-dependent membrane-binding protein annexin | JK724790 | 212 | 1.25 × 10−16 | 7.50% | Calcium-transporting ATPase activity |
| F1166 | Ca2+-dependent membrane-binding protein annexin | JK724790 | 212 | 1.25 × 10−16 | 87.50% | Calcium-transporting ATPase activity |
| F250 | Alpha glucosidase | JK724791 | 478 | 9.75 × 10−50 | 83.65% | Glucosidase activity |
| F465 | Ribonuclease HII family protein (RNaseHII) | JK724792 | 344 | 6.15 × 10−18 | 77.24% | Ribonuclease H activity |
| F499 | Anther-specific protein LAT52 precursor | JK724793 | 181 | 1.67 × 10−6 | 70.80% | Extracellular space |
| F519 | Actin 3 | JK724794 | 692 | 4.47 × 10−134 | 99.50% | Cytoskeleton |
| F546 | Pollen allergen Che a 1 precursor | JK724795 | 260 | 1.67 × 10−14 | 64.50% | Extracellular space |
| F1019 | Cell wall invertase (CWI) | JK724796 | 420 | 9.75 × 10−50 | 83.65% | Hydrolase activity |
| F580 | Zinc finger (C2H2 type) family protein | JK724797 | 373 | 3.65 × 10−22 | 75.20% | Transcription factor activity |
| F685 | Auxin-repressed protein | JK724798 | 595 | 1.18 × 10−9 | 61.67% | Signal transduction |
| F822 | WD-40 repeat protein-like (WD-40) | JK724799 | 465 | 6.61 × 10−41 | 94.25% | G-protein complex |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miao, H.; Ye, Z.; Silva, J.A.T.d.; Qin, Y.; Hu, G. Identifying Differentially Expressed Genes in Pollen from Self-Incompatible “Wuzishatangju” and Self-Compatible “Shatangju” Mandarins. Int. J. Mol. Sci. 2013, 14, 8538-8555. https://doi.org/10.3390/ijms14048538
Miao H, Ye Z, Silva JATd, Qin Y, Hu G. Identifying Differentially Expressed Genes in Pollen from Self-Incompatible “Wuzishatangju” and Self-Compatible “Shatangju” Mandarins. International Journal of Molecular Sciences. 2013; 14(4):8538-8555. https://doi.org/10.3390/ijms14048538
Chicago/Turabian StyleMiao, Hongxia, Zixing Ye, Jaime A. Teixeira da Silva, Yonghua Qin, and Guibing Hu. 2013. "Identifying Differentially Expressed Genes in Pollen from Self-Incompatible “Wuzishatangju” and Self-Compatible “Shatangju” Mandarins" International Journal of Molecular Sciences 14, no. 4: 8538-8555. https://doi.org/10.3390/ijms14048538