Impact of an Altered Wnt1/β-Catenin Expression on Clinicopathology and Prognosis in Clear Cell Renal Cell Carcinoma



Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Patient Characteristics
2.1.2. Expression of Wnt1 and β-Catenin in Normal Kidney Tissue and ccRCC
2.1.3. Correlation of Wnt1 and β-Catenin to Clinico-Pathologic Data in ccRCC
2.1.4. Correlation to OS and CSS of Wnt1 and β-Catenin
2.1.5. Prognostic Value of Wnt1 and β-Catenin on Survival
2.2. Discussion
3. Experimental Section
3.1. Patients and Tissue Microarrays
3.2. Immunohistochemical Staining and Data Analysis
3.3. Statistics
4. Conclusions
Conflict of Interest
References
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2012. CA A Cancer J. Clin 2012, 62, 10–29. [Google Scholar]
- Kruck, S.; Bedke, J.; Kuczyk, M.A.; Merseburger, A.S. Second-line systemic therapy for the treatment of metastatic renal cell cancer. Expert Rev. Anticancer Ther 2012, 12, 777–785. [Google Scholar]
- Bilim, V.; Kawasaki, T.; Katagiri, A.; Wakatsuki, S.; Takahashi, K.; Tomita, Y. Altered expression of beta-catenin in renal cell cancer and transitional cell cancer with the absence of beta-catenin gene mutations. Clin. Cancer Res 2000, 6, 460–466. [Google Scholar]
- Kim, Y.S.; Kang, Y.K.; Kim, J.B.; Han, S.A.; Kim, K.I.; Paik, S.R. Beta-catenin expression and mutational analysis in renal cell carcinomas. Pathol. Int 2000, 50, 725–730. [Google Scholar]
- Aaltomaa, S.; Lipponen, P.; Kärjä, V.; Lundstedt, S.; Lappi, J.; Kosma, V.-M. The expression and prognostic value of α-, β- and γ-catenins in renal cell carcinoma. Anticancer Res 2004, 24, 2407–2414. [Google Scholar]
- Bruder, E.; Moch, H.; Ehrlich, D.; Leuschner, I.; Harms, D.; Argani, P.; Briner, J.; Graf, N.; Selle, B.; Rufle, A.; et al. Wnt signaling pathway analysis in renal cell carcinoma in young patients. Modern Pathol. An Off. J. USA Can. Acad. Pathol. Inc 2007, 20, 1217–1229. [Google Scholar]
- Lian, X.; Duan, X.; Wu, X.; Li, C.; Chen, S.; Wang, S.; Cai, Y.; Weng, Z. Expression and clinical significance of von Hippel-Lindau downstream genes: Jade-1 and beta-catenin related to renal cell carcinoma. Urology 2012, 80, e487–e413. [Google Scholar]
- Hsu, R.J.; Ho, J.Y.; Cha, T.L.; Yu, D.S.; Wu, C.L.; Huang, W.P.; Chu, P.; Chen, Y.H.; Chen, J.T.; Yu, C.P. WNT10A plays an oncogenic role in renal cell carcinoma by activating WNT/beta-catenin pathway. PLoS One 2012, 7, e47649. [Google Scholar]
- Zang, T.; Zhuang, L.; Zhang, Z.; Xin, D.; Guo, Y. Expression of beta-catenin in renal cell carcinoma. Chin. Med. J 2001, 114, 152–154. [Google Scholar]
- Thevenod, F.; Chakraborty, P.K. The role of Wnt/beta-catenin signaling in renal carcinogenesis: lessons from cadmium toxicity studies. Curr. Mol. Med 2010, 10, 387–404. [Google Scholar]
- Valenta, T.; Hausmann, G.; Basler, K. The many faces and functions of beta-catenin. EMBO J 2012, 31, 2714–2736. [Google Scholar]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar]
- Satoh, S.; Daigo, Y.; Furukawa, Y.; Kato, T.; Miwa, N.; Nishiwaki, T.; Kawasoe, T.; Ishiguro, H.; Fujita, M.; Tokino, T.; et al. AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus-mediated transfer of AXIN1. Nat. Genet 2000, 24, 245–250. [Google Scholar]
- Cox, S.N.; Sallustio, F.; Serino, G.; Pontrelli, P.; Verrienti, R.; Pesce, F.; Torres, D.D.; Ancona, N.; Stifanelli, P.; Zaza, G.; et al. Altered modulation of WNT-[beta]-catenin and PI3K/Akt pathways in IgA nephropathy. Kidney Int 2010, 78, 396–407. [Google Scholar]
- Choi, H.; Chun, Y.S.; Kim, T.Y.; Park, J.W. HIF-2alpha enhances beta-catenin/TCF-driven transcription by interacting with beta-catenin. Cancer Res 2010, 70, 10101–10111. [Google Scholar]
- Herzlinger, D.; Qiao, J.; Cohen, D.; Ramakrishna, N.; Brown, A.M. Induction of kidney epithelial morphogenesis by cells expressing Wnt-1. Dev. Biol 1994, 166, 815–818. [Google Scholar]
- Katoh, M. Expression and regulation of WNT1 in human cancer: Up-regulation of WNT1 by beta-estradiol in MCF-7 cells. Int. J. Oncol 2003, 22, 209–212. [Google Scholar]
- Chen, S.; Guttridge, D.C.; You, Z.; Zhang, Z.; Fribley, A.; Mayo, M.W.; Kitajewski, J.; Wang, C.Y. Wnt-1 signaling inhibits apoptosis by activating beta-catenin/T cell factor-mediated transcription. J. Cell Biol 2001, 152, 87–96. [Google Scholar]
- Chen, G.; Shukeir, N.; Potti, A.; Sircar, K.; Aprikian, A.; Goltzman, D.; Rabbani, S.A. Up-regulation of Wnt-1 and β-catenin production in patients with advanced metastatic prostate carcinoma. Cancer 2004, 101, 1345–1356. [Google Scholar]
- Nusse, R.; Varmus, H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982, 31, 99–109. [Google Scholar]
- Röhrs, S.; Kutzner, N.; Vlad, A.; Grunwald, T.; Ziegler, S.; Müller, O. Chronological expression of Wnt target genes Ccnd1, Myc, Cdkn1a, Tfrc, Plf1 and Ramp3. Cell Biol. Int 2009, 33, 501–508. [Google Scholar]
- Janssens, N.; Andries, L.; Janicot, M.; Perera, T.; Bakker, A. Alteration of frizzled expression in renal cell carcinoma. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med 2004, 25, 161–171. [Google Scholar]
- Janssens, N.; Janicot, M.; Perera, T. The Wnt-dependent signaling pathways as target in oncology drug discovery. Investig. New Drugs 2006, 24, 263–280. [Google Scholar]
- Inoki, K.; Ouyang, H.; Zhu, T.; Lindvall, C.; Wang, Y.; Zhang, X.; Yang, Q.; Bennett, C.; Harada, Y.; Stankunas, K.; et al. TSC2 integrates Wnt and energy signals via a coordinated phosphorylation by AMPK and GSK3 to regulate cell growth. Cell 2006, 126, 955–968. [Google Scholar]
- Mizushima, T.; Nakagawa, H.; Kamberov, Y.G.; Wilder, E.L.; Klein, P.S.; Rustgi, A.K. Wnt-1 but not epidermal growth factor induces beta-catenin/T-cell factor-dependent transcription in esophageal cancer cells. Cancer Res 2002, 62, 277–282. [Google Scholar]
- Heuberger, J.; Birchmeier, W. Interplay of cadherin-mediated cell adhesion and canonical Wnt signaling. Cold Spring Harb. Perspect. Biol 2010, 2, a002915. [Google Scholar]
- Kraus, C.; Liehr, T.; Hulsken, J.; Behrens, J.; Birchmeier, W.; Grzeschik, K.H.; Ballhausen, W.G. Localization of the human beta-catenin gene (CTNNB1) to 3p21: A region implicated in tumor development. Genomics 1994, 23, 272–274. [Google Scholar]
- Maestro, M.L.; del Barco, V.; Sanz-Casla, M.T.; Moreno, J.; Adrover, E.; Izquierdo, L.; Zanna, I.; Fernandez, C.; Redondo, E.; Blanco, J.; et al. Loss of heterozygosity on the short arm of chromosome 3 in renal cancer. Oncology 2000, 59, 126–130. [Google Scholar]
- Zeitlin, B.D.; Ellis, L.M.; Nör, J.E. Inhibition of vascular endothelial growth factor receptor-1/Wnt/β-catenin crosstalk leads to tumor cell death. Clin. Cancer Res 2009, 15, 7453–7455. [Google Scholar]
- Kawano, Y.; Kypta, R. Secreted antagonists of the Wnt signalling pathway. J. Cell Sci 2003, 116, 2627–2634. [Google Scholar]
- Dahl, E.; Wiesmann, F.; Woenckhaus, M.; Stoehr, R.; Wild, P.J.; Veeck, J.; Knuchel, R.; Klopocki, E.; Sauter, G.; Simon, R.; et al. Frequent loss of SFRP1 expression in multiple human solid tumours: Association with aberrant promoter methylation in renal cell carcinoma. Oncogene 2007, 26, 5680–5691. [Google Scholar]
- Awakura, Y.; Nakamura, E.; Ito, N.; Kamoto, T.; Ogawa, O. Methylation-associated silencing of SFRP1 in renal cell carcinoma. Oncol. Rep 2008, 20, 1257–1263. [Google Scholar]
- Gumz, M.L.; Zou, H.; Kreinest, P.A.; Childs, A.C.; Belmonte, L.S.; LeGrand, S.N.; Wu, K.J.; Luxon, B.A.; Sinha, M.; Parker, A.S.; et al. Secreted frizzled-related protein 1 loss contributes to tumor phenotype of clear cell renal cell carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res 2007, 13, 4740–4749. [Google Scholar]
- Yamamura, S.; Kawakami, K.; Hirata, H.; Ueno, K.; Saini, S.; Majid, S.; Dahiya, R. Oncogenic functions of secreted Frizzled-related protein 2 in human renal cancer. Mol. Cancer Ther 2010, 9, 1680–1687. [Google Scholar]
- Kawakami, K.; Hirata, H.; Yamamura, S.; Kikuno, N.; Saini, S.; Majid, S.; Tanaka, Y.; Kawamoto, K.; Enokida, H.; Nakagawa, M.; et al. Functional significance of Wnt inhibitory factor-1 gene in kidney cancer. Cancer Res 2009, 69, 8603–8610. [Google Scholar]
- Hirata, H.; Hinoda, Y.; Nakajima, K.; Kawamoto, K.; Kikuno, N.; Ueno, K.; Yamamura, S.; Zaman, M.S.; Khatri, G.; Chen, Y.; et al. Wnt antagonist DKK1 acts as a tumor suppressor gene that induces apoptosis and inhibits proliferation in human renal cell carcinoma. Int. J. Cancer 2011, 128, 1793–1803. [Google Scholar]
- Hirata, H.; Hinoda, Y.; Nakajima, K.; Kawamoto, K.; Kikuno, N.; Kawakami, K.; Yamamura, S.; Ueno, K.; Majid, S.; Saini, S.; et al. Wnt antagonist gene DKK2 is epigenetically silenced and inhibits renal cancer progression through apoptotic and cell cycle pathways. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res 2009, 15, 5678–5687. [Google Scholar]
- Ueno, K.; Hirata, H.; Majid, S.; Chen, Y.; Zaman, M.S.; Tabatabai, Z.L.; Hinoda, Y.; Dahiya, R. Wnt antagonist DICKKOPF-3 (Dkk-3) induces apoptosis in human renal cell carcinoma. Mol. Carcinogen 2011, 50, 449–457. [Google Scholar]
- Ronkainen, H.; Kauppila, S.; Hirvikoski, P.; Vaarala, M.H. Evaluation of myosin VI, E-cadherin and beta-catenin immunostaining in renal cell carcinoma. J. Exp. Clin. Cancer Res. CR 2010, 29, 2. [Google Scholar]
- Kruck, S.; Merseburger, A.S.; Hennenlotter, J.; Scharpf, M.; Eyrich, C.; Amend, B.; Sievert, K.D.; Stenzl, A.; Bedke, J. High cytoplasmic expression of p27(Kip1) is associated with a worse cancer-specific survival in clear cell renal cell carcinoma. BJU Int 2012, 109, 1565–1570. [Google Scholar]
- Cheng, S.L.; Shao, J.S.; Cai, J.; Sierra, O.L.; Towler, D.A. Msx2 exerts bone anabolism via canonical Wnt signaling. J. Biol. Chem 2008, 283, 20505–20522. [Google Scholar]
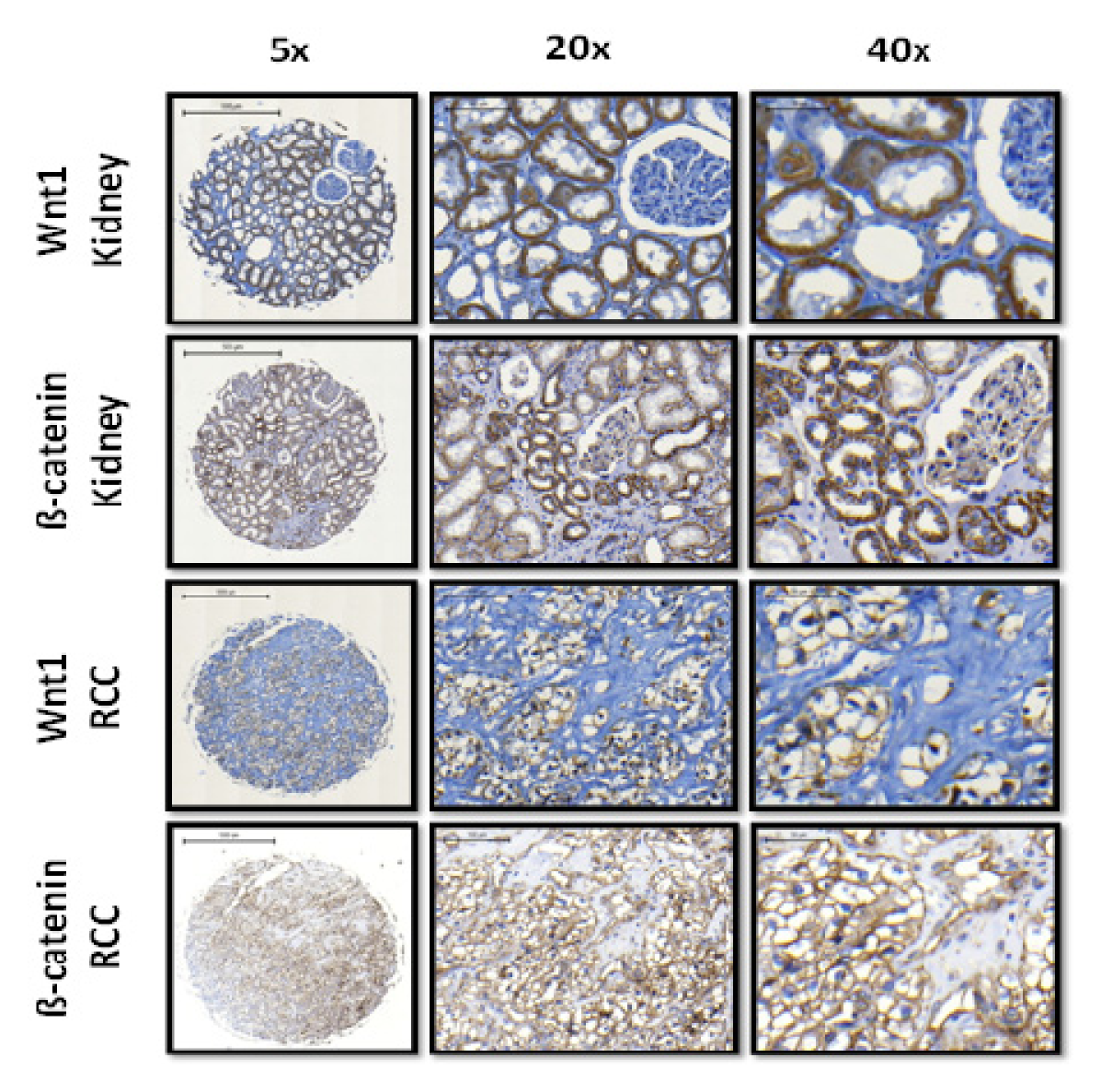


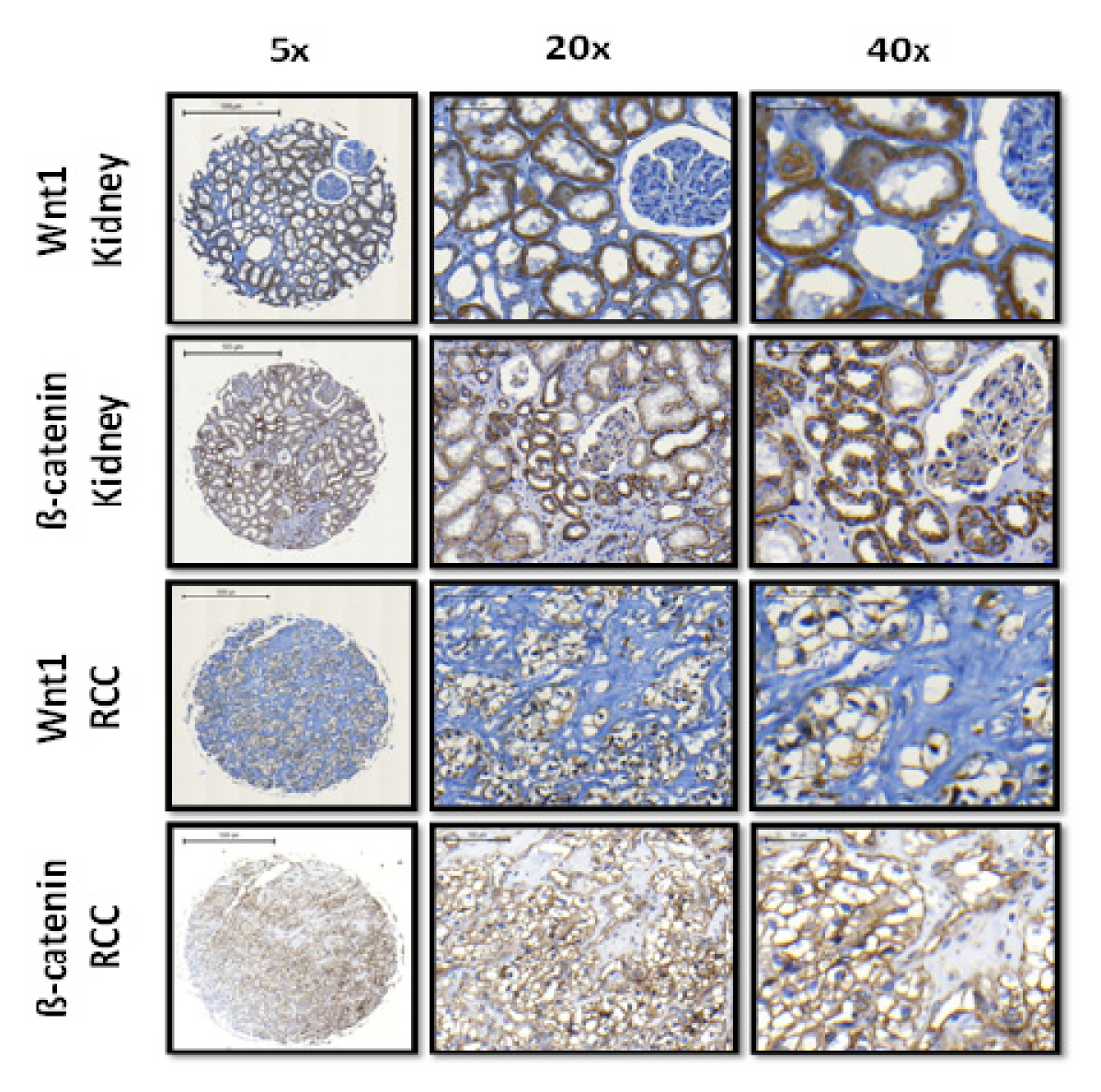{kind=link}
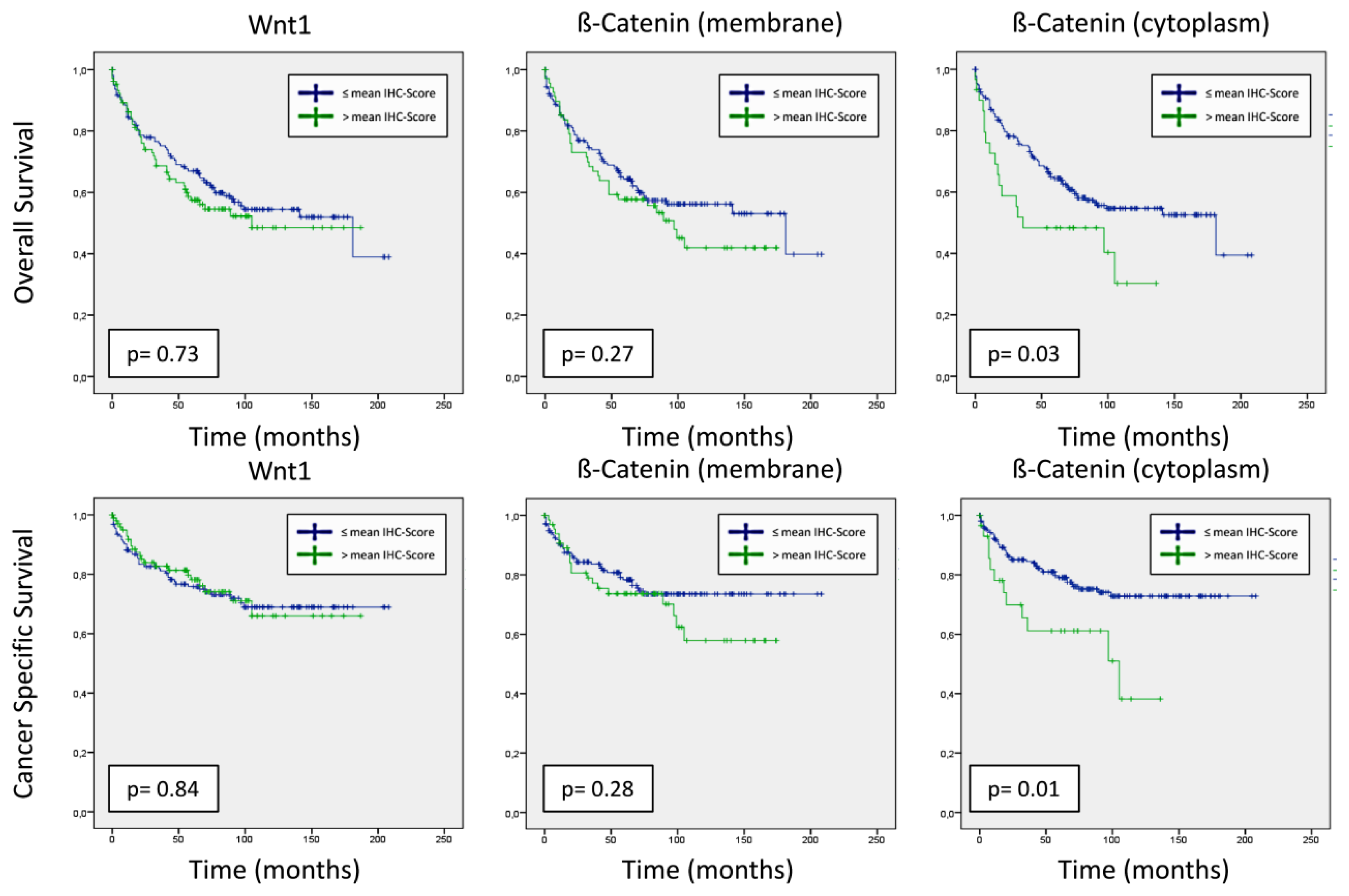{kind=link}
| Parameter | Overall (n = 278) | Wnt1 low # (n = 165) | Wnt1 high # (n = 106) | p-Value | Membranous β-Catenin low # (n = 186) | Membranous β-Catenin high # (n = 70) | p-Value | Cytoplasmic β-Catenin low # (n = 225) | Cytoplasmic β-Catenin high # (n = 31) | p-value |
|---|---|---|---|---|---|---|---|---|---|---|
| Age (mean ± SD; years) | 62.2 (±12.5) | 61.4 (±12.9) | 63.9 (±11.6) | 0.12 | 61.8 (±12.6) | 64.2 (±12.3) | 0.17 | 62.2 (±12.7) | 64.1 (±11.7) | 0.57 |
| Gender (Male/Female; %) | 194/84 (69.8/30.2) | 108/57 (65.5/34.5) | 80/26 (75.5/24.5) | 0.08 | 134/52 (72.0/28.0) | 43/27 (61.4/38.6) | 0.11 | 157/68 (69.8/30.2) | 20/11 (64.5/35.5) | 0.56 |
| Total deaths (%) | 113 (41.4) | 66 (59.5) | 45 (40.5) | 0.80 | 72 (67.9) | 34 (32.1) | 0.20 | 89 (84.0) | 17 (16.0) | 0.11 |
| Cancer related deaths (%) | 65 (23.8) | 41 (64.1) | 23 (35.9) | 0.56 | 39 (66.1) | 20 (33.9) | 0.24 | 47 (79.7) | 12 (20.3) | 0.04 * |
| Tumor Diameter (mean ± SD; cm) | 5.26 (±2.91) | 4.9 (±2.6) | 5.8 (±3.0) | 0.01 * | 5.3 (±2.8) | 5.4 (±2.8) | 0.87 | 5.2 (±2.7) | 6.4 (±2.9) | 0.01 * |
| Stage (T1/2 vs. T3/4, %) | 169/109 (60.8/39.2) | 114/51 (69.1/30.9) | 50/56 (47.2/52.8) | 0.004 * | 119/67 (64.0/36.0) | 34/36 (48.6/61.4) | 0.03 * | 144/81 (64.0/36.0) | 9/22 (29.0/71.0) | 0.003 * |
| Lymph Nodes (N0 vs. N1/2, %) | 265/13 (95.3/4.7) | 157/8 (95.2/4.8) | 101/5 (95.3/4.7) | 1.0 | 180/6 (96.8/3.2) | 64/6 (91.4/8.6) | 0.10 | 217/8 (96.4/3.6) | 27/4 (87.1/12.9) | 0.04 * |
| Distant Metastasis (M0 vs. M1, %) | 239/39 (86.0/14.0) | 140/25 (84.85/15.15) | 93/13 (86.0/14.0) | 0.59 | 159/27 (85.5/14.5) | 61/9 (87.1/12.9) | 0.84 | 193/32 (85.8/14.2) | 27/4 (87.1/12.9) | 1.0 |
| Grade (G1/2 vs. G3/4, %) | 234/44 (84.2/15.8) | 136/29 (82.0/17.6) | 92/14 (86.8/13.2) | 0.40 | 162/24 (87.1/12.9) | 53/17 (75.7/24.3) | 0.04 * | 199/26 (88.4/11.6) | 16/15 (51.6/48.4) | ≤0.001 * |
| Vascular invasion (no/yes, %) | 193/85 (69.4/30.6) | 124/41 (75.15/24.85) | 64/42 (60.4/39.6) | 0.02 * | 137/49 (73.7/26.3) | 39/31 (55.7/44.3) | 0.01 * | 163/62 (72.4/27.6) | 13/18 (41.9/58.1) | 0.002 * |
| Perinephric Invasion (%) | 47 (16.9) | 157/8 (95.1/4.9) | 94/12 (88.7/11.3) | 0.06 | 171/15 (91.9/8.1) | 66/4 (94.3/5.7) | 0.60 | 208/17 (92.4/7.6) | 29/2 (93.55/6.45) | 1.0 |
| Sinus Invasion (%) | 65 (23.4) | 143/22 (86.7/13.3) | 89/17 (84.0/16.0) | 0.60 | 157/29 (84.4/15.6) | 62/8 (88.6/11.4) | 0.55 | 194/31 (86.2/13.8) | 25/6 (80.65/19.35) | 0.42 |
| Necrosis (%) | 114 (41.0) | 100/65 (60.6/39.4) | 59/47 (55.7/44.3) | 0.45 | 114/72 (61.3/38.7) | 33/37 (47.1/52.9) | 0.04 * | 133/92 (59.1/40.9) | 14/17 (45.2/54.8) | 0.18 |
| Sarcomatoid features (%) | 19 (6.8) | 152/13 (92.1/7.9) | 100/6 (93.0/7.0) | 0.63 | 175/11 (94.1/5.9) | 63/7 (93.0/7.0) | 0.28 | 213/12 (94.7/5.3) | 25/6 (80.65/19.35) | 0.01 * |
| Hazard Ratio (95% CI) | |||||
|---|---|---|---|---|---|
| Variable | Categories | Overall survival | p-value | Cancer specific survival | p-value |
| Age | ±median | 1.69 (1.15–2.48) | 0.007 | 1.00 (0.62–1.64) | 0.97 |
| Gender | female vs. male | 0.90 (0.59–1.26) | 0.6 | 0.58 (0.31–1.07) | 0.08 |
| T-stage | T1/2 vs. T3/4 | 4.32 (2.93–6.37) | <0.0001 | 9.62 (5.20–17.77) | <0.0001 |
| N-stage | N0 vs. N1/2 | 3.22 (1.68–6.19) | <0.0001 | 5.01 (2.50–10.20) | <0.0001 |
| M-stage | M0 vs. M1 | 5.51 (3.59–8.47) | <0.0001 | 9.57 (5.65–16.19) | <0.0001 |
| Nuclear Grade | G1/2 vs. G3/4 | 3.80 (2.52–5.72) | <0.0001 | 6.78 (4.13–11.13) | <0.0001 |
| Tumor Necrosis | yes vs. no | 1.80 (1.24–2.60) | 0.002 | 2.81 (1.70–4.66) | <0.0001 |
| Sarcomatoid Differentiation | yes vs. no | 4.60 (2.72–7.76) | <0.0001 | 7.36 (4.08–13.25) | <0.0001 |
| Vascular Invasion | yes vs. no | 3.10 (2.13–4.50) | <0.0001 | 5.67 (3.39–9.50) | <0.0001 |
| Perinephric Invasion | yes vs. no | 2.53(1.23–4.11) | 0.008 | 2.20 (1.00–4.84) | 0.5 |
| Sinus Invasion | yes vs. no | 2.29 (1.46–3.58) | <0.0001 | 2.78 (1.59–4.85) | <0.0001 |
| Wnt1 Score tumor | ±mean | 1.18 (0.81–1.73) | 0.4 | 0.95 (0.57–1.58) | 0.84 |
| β-catenin Score membrane tumor | ±mean | 1.26 (0.84–1.89) | 0.27 | 1.35 (0.79–2.30) | 0.28 |
| β-catenin Score cytoplasm tumor | ±mean | 1.75 (1.04–2.94) | 0.035 | 2.26 (1.20–4.27) | 0.012 |
| Hazard Ratio (95% CI) | |||||
|---|---|---|---|---|---|
| Variable | Categories | Overall survival | p-value | Cancer specific survival | p-value |
| Age | ±median | 1.88 (1.25–2.83) | 0.002 | 1.18 (0.69–2.00) | 0.56 |
| T-stage | T1/2 vs. T3/4 | 2.63 (1.63–4.23) | <0.0001 | 4.30 (2.03–9.10) | <0.0001 |
| N-stage | N0 vs. N1/2 | 1.59 (0.75–3.42) | 0.23 | 1.89 (0.83–4.32) | 0.13 |
| M-stage | M0 vs. M1 | 3.43 (2.09–5.62) | <0.0001 | 4.58 (2.46–8.52) | <0.0001 |
| Nuclear Grade | G1/2 vs. G3/4 | 0.98 (0.54–1.80) | 0.96 | 1.37 (0.69–2.73) | 0.37 |
| Tumor Necrosis | yes vs. no | 1.23 (0.83–1.84) | 0.3 | 1.65 (0.94–2.90) | 0.08 |
| Sarcomatoid Differentiation | yes vs. no | 1.63 (0.83–3.20) | 0.15 | 1.62 (1.68–3.45) | 0.16 |
| β-catenin Score cytoplasm tumor | ±mean | 1.00 (0.55–1.84) | 0.99 | 0.88 (0.43–1.83) | 0.75 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kruck, S.; Eyrich, C.; Scharpf, M.; Sievert, K.-D.; Fend, F.; Stenzl, A.; Bedke, J. Impact of an Altered Wnt1/β-Catenin Expression on Clinicopathology and Prognosis in Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2013, 14, 10944-10957. https://doi.org/10.3390/ijms140610944
Kruck S, Eyrich C, Scharpf M, Sievert K-D, Fend F, Stenzl A, Bedke J. Impact of an Altered Wnt1/β-Catenin Expression on Clinicopathology and Prognosis in Clear Cell Renal Cell Carcinoma. International Journal of Molecular Sciences. 2013; 14(6):10944-10957. https://doi.org/10.3390/ijms140610944
Chicago/Turabian StyleKruck, Stephan, Christian Eyrich, Marcus Scharpf, Karl-Dietrich Sievert, Falco Fend, Arnulf Stenzl, and Jens Bedke. 2013. "Impact of an Altered Wnt1/β-Catenin Expression on Clinicopathology and Prognosis in Clear Cell Renal Cell Carcinoma" International Journal of Molecular Sciences 14, no. 6: 10944-10957. https://doi.org/10.3390/ijms140610944