Transgene IL-6 Enhances DC-Stimulated CTL Responses by Counteracting CD4+25+Foxp3+ Regulatory T Cell Suppression via IL-6-Induced Foxp3 Downregulation

Abstract
:1. Introduction
2. Results and Discussion
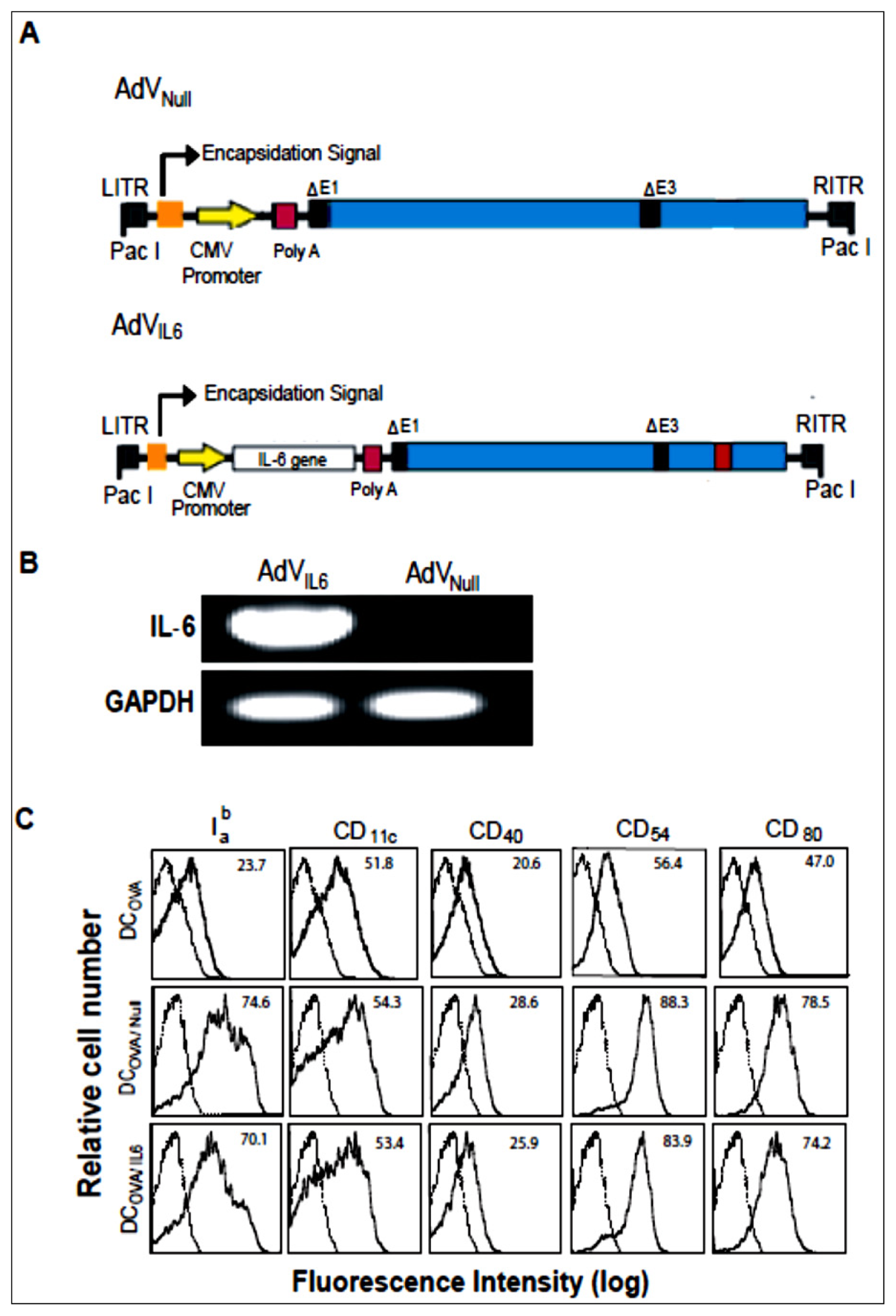
2.1. AdVIL-6-Transfected DCs Upregulate Expression of Iab, CD54, CD80 and IL-6
2.2. AdVIL-6-Transfected DCs Stimulate Potent CTL Responses
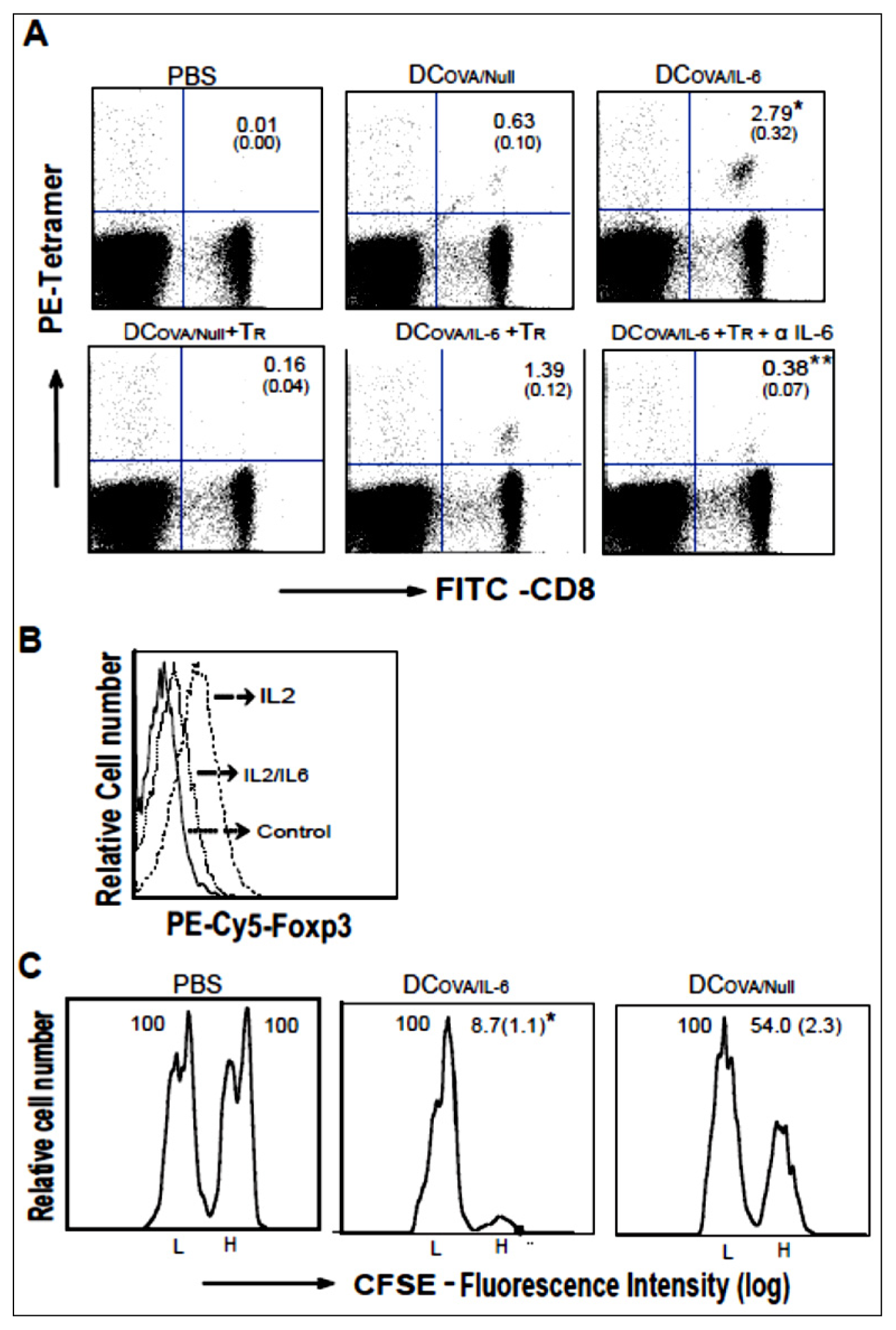
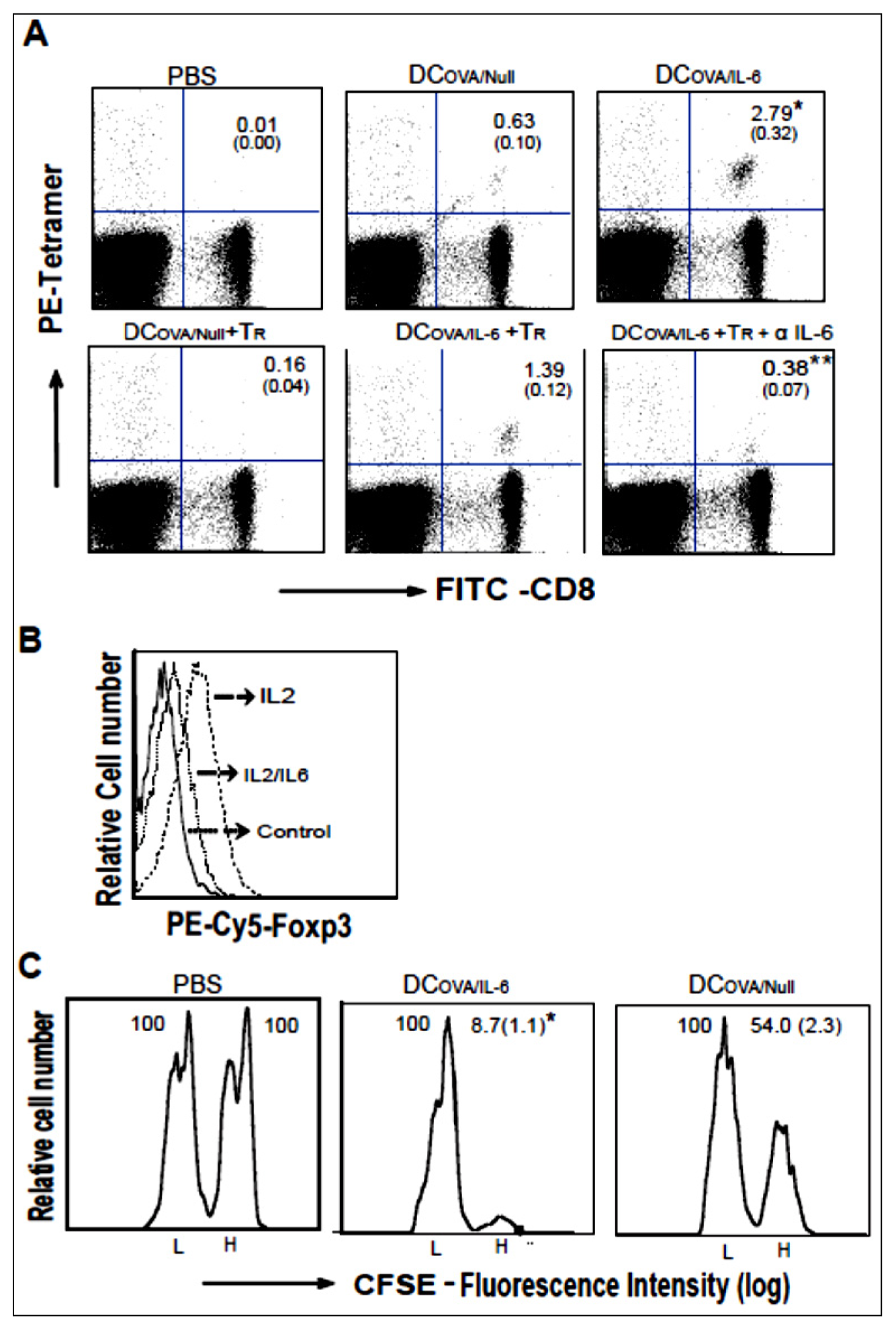
2.3. AdVIL-6-Transfected DCs Counteract CD4+25+Foxp3+ Tr Immunosuppression via Transgene Encoded IL-6 Signaling
2.4. IL-6 Induces Foxp3 down-Regulation of CD4+25+Foxp3+ Tr Cells
2.5. AdVIL-6-Transfected DC-Stimulated CD8+ T Cells Are Effector CTLs
2.6. AdVIL-6-Transfected DCs Induce Potent Antitumor Immunity
2.7. Discussion
3. Experimental Section
3.1. Reagents, Cell Lines and Animals
3.2. Recombinant Adenovirus Construction
3.3. Preparation of Dendritic Cells
3.4. Preparation of CD4+25+Foxp3+ Regulatory T (Tr) Cells
3.5. Adenovirus (AdV) Transfection of DCs
3.6. Flow Cytometric Analysis
3.7. In Vivo Cytotoxicity Assay
3.8. Animal Studies
3.9. Statistical Analyses
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ye, Z.; Tang, C.; Xu, S.; Zhang, B.; Zhang, X.; Moyana, T.; Yang, J.; Xiang, J. Type 1 CD8+ T cells are superior to type 2 CD8+ T cells in tumor immunotherapy due to their efficient cytotoxicity, prolonged survival and type 1 immune modulation. Cell. Mol. Immunol 2007, 4, 277–285. [Google Scholar]
- Klebanoff, C.A.; Acquavella, N.; Yu, Z.; Restifo, N.P. Therapeutic cancer vaccines: Are we there yet? Immunol. Rev 2011, 239, 27–44. [Google Scholar]
- Palucka, K.; Ueno, H.; Banchereau, J. Recent developments in cancer vaccines. J. Immunol 2011, 186, 1325–1331. [Google Scholar]
- Galluzzi, L.; Senovilla, L.; Vacchelli, E.; Eggermont, A.; Fridman, W.H.; Galon, J.; Sautès-Fridman, C.; Tartour, E.; Zitvogel, L.; Kroemer, G. Trial watch: Dendritic cell-based interventions for cancer therapy. Oncoimmunology 2012, 1, 1111–1134. [Google Scholar]
- Melief, C.J. Cancer immunotherapy by dendritic cells. Immunity 2008, 29, 372–383. [Google Scholar]
- Rosenberg, S.A.; Yang, J.C.; Restifo, N.P. Cancer immunotherapy: Moving beyond current vaccines. Nat. Med 2004, 10, 909–915. [Google Scholar]
- Schluns, K.S.; Lefrancois, L. Cytokine control of memory T-cell development and survival. Nat. Rev. Immunol 2003, 3, 269–279. [Google Scholar]
- Akiyama, Y.; Watanabe, M.; Maruyama, K.; Ruscetti, F.W.; Wiltrout, R.H.; Yamaguchi, K. Enhancement of antitumor immunity against B16 melanoma tumor using genetically modified dendritic cells to produce cytokines. Gene Ther 2000, 7, 2113–2121. [Google Scholar]
- Ojima, T.; Iwahashi, M.; Nakamura, M.; Matsuda, K.; Naka, T.; Nakamori, M.; Ueda, K.; Ishida, K.; Yamaue, H. The boosting effect of co-transduction with cytokine genes on cancer vaccine therapy using genetically modified dendritic cells expressing tumor-associated antigen. Int. J. Oncol 2006, 28, 947–953. [Google Scholar]
- Tourkova, I.L.; Yurkovetsky, Z.R.; Gambotto, A.; Makarenkova, V.P.; Perez, L.; Balkir, L.; Robbins, P.D.; Shurin, M.R.; Shurin, G.V. Increased function and survival of IL-15-transduced human dendritic cells are mediated by up-regulation of IL-15Ralpha and Bcl-2. J. Leukoc. Biol 2002, 72, 1037–1045. [Google Scholar]
- Zhang, W.; Chen, Z.; Li, F.; Kamencic, H.; Juurlink, B.; Gordon, J.R.; Xiang, J. Tumour necrosis factor-alpha (TNF-alpha) transgene-expressing dendritic cells (DCs) undergo augmented cellular maturation and induce more robust T-cell activation and anti-tumour immunity than DCs generated in recombinant TNF-alpha. Immunology 2003, 108, 177–188. [Google Scholar]
- Pasare, C.; Medzhitov, R. Toll pathway-dependent blockade of CD4+CD25+ T cell-mediated suppression by dendritic cells. Science 2003, 299, 1033–1036. [Google Scholar]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006, 441, 235–238. [Google Scholar]
- Schuler, G.; Steinman, R.M. Dendritic cells as adjuvants for immune-mediated resistance to tumors. J. Exp. Med 1997, 186, 1183–1187. [Google Scholar]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor α. J. Exp. Med 1994, 17, 1109–1118. [Google Scholar]
- Palucka, K.; Banchereau, J. Dendritic-cell-based therapeutic cancer vaccines. Immunity 2013, 39, 38–48. [Google Scholar]
- Strioga, M.M.; Felzmann, T.; Powell, D.J., Jr.; Ostapenko, V.; Dobrovolskiene, N.T.; Matuskova, M.; Michalek, J.; Schijns, V.E. Therapeutic dendritic cell-based cancer vaccines: The state of the art. Crit. Rev. Immunol 2013, 33, 489–547. [Google Scholar]
- Huang, C.; Ramakrishnan, R.; Trkulja, M.; Ren, X.; Gabrilovich, D.I. Therapeutic effect of intratumoral administration of DCs with conditional expression of combination of different cytokines. Cancer Immunol. Immunother 2012, 61, 573–579. [Google Scholar]
- Vogt, A.; Sievers, E.; Lukacs-Kornek, V.; Decker, G.; Raskopf, E.; Meumann, N.; Büning, H.; Sauerbruch, T.; Strassburg, C.P.; Schmidt-Wolf, I.G.; et al. Improving immunotherapy of hepatocellular carcinoma (HCC) using dendritic cells (DC) engineered to express IL-12 in vivo. Liver Int. 2013. [Google Scholar] [CrossRef]
- Qu, Y.; Taylor, J.L.; Bose, A.; Storkus, W.J. Therapeutic effectiveness of intratumorally delivered dendritic cells engineered to express the pro-inflammatory cytokine, interleukin (IL)-32. Cancer Gene Ther 2011, 18, 663–673. [Google Scholar]
- Taga, T.; Kishimoto, T. Gp130 and the interleukin-6 family of cytokines. Annu. Rev. Immunol 1997, 15, 797–819. [Google Scholar]
- Naugler, W.E.; Karin, M. The wolf in sheep’s clothing: The role of interleukin-6 in immunity, inflammation and cancer. Trends Mol. Med 2008, 14, 109–119. [Google Scholar]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol 2005, 6, 1123–1132. [Google Scholar]
- Weaver, C.T.; Hatton, R.D.; Mangan, P.R.; Harrington, L.E. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu. Rev. Immunol 2007, 25, 821–852. [Google Scholar]
- Ankathatti Munegowda, M.; Xu, S.; Freywald, A.; Xiang, J. CD4+ Th2 cells function alike effector Tr1 and Th1 cells through the deletion of a single cytokine IL-6 and IL-10 gene. Mol. Immunol 2012, 51, 143–149. [Google Scholar]
- Liu, Y.; Zhang, X.; Zhang, W.; Chen, Z.; Chan, T.; Ali, K.; Jia, Z.; Xiang, J. Adenovirus-mediated CD40 ligand gene-engineered dendritic cells elicit enhanced CD8(+) cytotoxic T-cell activation and antitumor immunity. Cancer Gene Ther 2002, 9, 202–208. [Google Scholar]
- Hirschowitz, E.A.; Weaver, J.D.; Hidalgo, G.E.; Doherty, D.E. Murine dendritic cells infected with adenovirus vectors show signs of activation. Gene Ther 2000, 7, 1112–1120. [Google Scholar]
- Rouard, H.; Leon, A.; Klonjkowski, B.; Marquet, J.; Tenneze, L.; Plonquet, A.; Agrawal, S.G.; Abastado, J.P.; Eloit, M.; Farcet, J.P.; et al. Adenoviral transduction of human “clinical grade” immature dendritic cells enhances costimulatory molecule expression and T-cell stimulatory capacity. J. Immunol. Methods 2000, 241, 69–81. [Google Scholar]
- Morelli, A.E.; Larregina, A.T.; Ganster, R.W.; Zahorchak, A.F.; Plowey, J.M.; Takayama, T.; Logar, A.J.; Robbins, P.D.; Falo, L.D.; Thomson, A.W. Recombinant adenovirus induces maturation of dendritic cells via an NF-κB-dependent pathway. J. Virol 2000, 74, 9617–9628. [Google Scholar]
- Philpott, N.J.; Nociari, M.; Elkon, K.B.; Falck-Pedersen, E. Adenovirus-induced maturation of dendritic cells through a PI3 kinase-mediated TNF-alpha induction pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 6200–6205. [Google Scholar]
- Bluestone, J.A.; Abbas, A.K. Natural vs. adaptive regulatory T cells. Nat. Rev. Immunol 2003, 3, 253–257. [Google Scholar]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol 2003, 4, 330–336. [Google Scholar]
- Hong, J.; Li, N.; Zhang, X.; Zheng, B.; Zhang, J.Z. Induction of CD4+CD25+ regulatory T cells by copolymer-I through activation of transcription factor Foxp3. Proc. Natl. Acad. Sci. USA 2005, 102, 6449–6454. [Google Scholar]
- Lipscomb, M.W.; Taylor, J.L.; Goldbach, C.J.; Watkins, S.C.; Wesa, A.K.; Storkus, W.J. DC expressing transgene Foxp3 are regulatory APC. Eur. J. Immunol 2010, 40, 480–493. [Google Scholar]
- Liyanage, U.K.; Moore, T.T.; Joo, H.G.; Tanaka, Y.; Herrmann, V.; Doherty, G.; Drebin, J.A.; Strasberg, S.M.; Eberlein, T.J.; Goedegebuure, P.S.; et al. Prevalence of regulatory T cells is increased in peripheral blood and tumor microenvironment of patients with pancreas or breast adenocarcinoma. J. Immunol 2002, 169, 2756–2761. [Google Scholar]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med 2004, 10, 942–949. [Google Scholar]
- Levings, M.K.; Sangregorio, R.; Roncarolo, M.G. Human CD25(+)CD4(+) t regulatory cells suppress naive and memory T cell proliferation and can be expanded in vitro without loss of function. J. Exp. Med 2001, 193, 1295–1302. [Google Scholar]
- Martin, B.; Banz, A.; Bienvenu, B.; Cordier, C.; Dautigny, N.; Bécourt, C.; Lucas, B. Suppression of CD4+ T lymphocyte effector functions by CD4+CD25+ cells in vivo. J. Immunol. 2004, 172, 3391–3398. [Google Scholar]
- Nishikawa, H.; Jäger, E.; Ritter, G.; Old, L.J.; Gnjatic, S. CD4+CD25+ regulatory T cells control the induction of antigen-specific CD4+ helper T cell responses in cancer patients. Blood 2005, 106, 1008–1011. [Google Scholar]
- Misra, N.; Bayry, J.; Lacroix-Desmazes, S.; Kazatchkine, M.D.; Kaveri, S.V. Cutting edge: Human CD4+CD25+ T cells restrain the maturation and antigen-presenting function of dendritic cells. J. Immunol 2004, 172, 4676–4680. [Google Scholar]
- Pardoll, D.M. Cancer vaccines. Nat. Med 1998, 4, 525–531. [Google Scholar]
- Ahmed, K.A.; Wang, L.; Munegowda, M.A.; Mulligan, S.J.; Gordon, J.R.; Griebel, P.; Xiang, J. Direct in vivo evidence of CD4+ T cell requirement for CTL response and memory via pMHC-I targeting and CD40L signaling. J. Leukoc. Biol 2012, 92, 289–300. [Google Scholar]
- Moulin, V.; Morgan, M.E.; Eleveld-Trancikova, D.; Haanen, J.B.; Wielders, E.; Looman, M.W.; Janssen, R.A.; Figdor, C.G.; Jansen, B.J.; Adema, G.J. Targeting dendritic cells with antigen via dendritic cell-associated promoters. Cancer Gene Ther 2012, 19, 303–311. [Google Scholar]
- Yewdall, A.W.; Drutman, S.B.; Jinwala, F.; Bahjat, K.S.; Bhardwaj, N. CD8+ T cell priming by dendritic cell vaccines requires antigen transfer to endogenous antigen presenting cells. PLoS One 2010, 5, e11144. [Google Scholar]
- Sato, M.; Chamoto, K.; Nishimura, T. A novel tumor-vaccine cell therapy using bone marrow-derived dendritic cell type 1 and antigen-specific Th1 cells. Int. Immunol 2003, 15, 837–843. [Google Scholar]
- Ben-Sasson, S.Z.; Makedonski, K.; Hu-Li, J.; Paul, W.E. Survival and cytokine polarization of naive CD4(+) T cells in vitro is largely dependent on exogenous cytokines. Eur. J. Immunol 2000, 30, 1308–1317. [Google Scholar]
- Curnow, S.J.; Scheel-Toellner, D.; Jenkinson, W.; Raza, K.; Durrani, O.M.; Faint, J.M.; Rauz, S.; Wloka, K.; Pilling, D.; Rose-John, S.; et al. Inhibition of T cell apoptosis in the aqueous humor of patients with uveitis by IL-6/soluble IL-6 receptor trans-signaling. J. Immunol 2004, 173, 5290–5297. [Google Scholar]
- Teague, T.K.; Schaefer, B.C.; Hildeman, D.; Bender, J.; Mitchell, T.; Kappler, J.W.; Marrack, P. Activation-induced inhibition of interleukin 6-mediated T cell survival and signal transducer and activator of transcription 1 signaling. J. Exp. Med 2000, 191, 915–926. [Google Scholar]
- Xia, D.; Hao, S.; Xiang, J. CD8+ cytotoxic T-APC stimulate central memory CD8+ T cell responses via acquired peptide-MHC class I complexes and CD80 costimulation, and IL-2 secretion. J. Immunol 2006, 177, 2976–2984. [Google Scholar]
- Chan, T.; Sami, A.; El-Gayed, A.; Guo, X.; Xiang, J. HER-2/neu-gene engineered dendritic cell vaccine stimulates stronger HER-2/neu-specific immune responses compared to DNA vaccination. Gene Ther 2006, 13, 1391–1402. [Google Scholar]
- Chen, Y.; Xie, Y.; Chan, T.; Sami, A.; Ahmed, S.; Liu, Q.; Xiang, J. Adjuvant effect of HER-2/neu-specific adenoviral vector stimulating CD8+ T and natural killer cell responses on anti-HER-2/neu antibody therapy for well-established breast tumors in HER-2/neu transgenic mice. Cancer Gene Ther 2011, 18, 489–499. [Google Scholar]
- Xie, Y.; Zhang, X.; Zhao, T.; Li, W.; Xiang, J. Natural CD8+25+ regulatory T cell-secreted exosomes capable of suppressing cytotoxic T lymphocyteYewdall-mediated immunity against B16 melanoma. Biochem. Biophys. Res. Commun 2013, 438, 152–155. [Google Scholar]


{kind=link}
{kind=link}
| Animal groups | Tumor cell challenge | Tumor bearing mice (%) | Median number of lung tumor colonies |
|---|---|---|---|
| DCOVA/Null | BL6-10OVA | 4/8 (50) | 49 ± 13 * |
| DCOVA/IL-6 | BL6-10OVA | 0/8 (0) | 0 |
| PBS | BL6-10OVA | 8/8 (100) | >300 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bhanumathy, K.K.; Zhang, B.; Ahmed, K.A.; Qureshi, M.; Xie, Y.; Tao, M.; Tan, X.; Xiang, J. Transgene IL-6 Enhances DC-Stimulated CTL Responses by Counteracting CD4+25+Foxp3+ Regulatory T Cell Suppression via IL-6-Induced Foxp3 Downregulation. Int. J. Mol. Sci. 2014, 15, 5508-5521. https://doi.org/10.3390/ijms15045508
Bhanumathy KK, Zhang B, Ahmed KA, Qureshi M, Xie Y, Tao M, Tan X, Xiang J. Transgene IL-6 Enhances DC-Stimulated CTL Responses by Counteracting CD4+25+Foxp3+ Regulatory T Cell Suppression via IL-6-Induced Foxp3 Downregulation. International Journal of Molecular Sciences. 2014; 15(4):5508-5521. https://doi.org/10.3390/ijms15045508
Chicago/Turabian StyleBhanumathy, Kalpana Kalyanasundaram, Bei Zhang, Khawaja Ashfaque Ahmed, Mabood Qureshi, Yufeng Xie, Min Tao, Xin Tan, and Jim Xiang. 2014. "Transgene IL-6 Enhances DC-Stimulated CTL Responses by Counteracting CD4+25+Foxp3+ Regulatory T Cell Suppression via IL-6-Induced Foxp3 Downregulation" International Journal of Molecular Sciences 15, no. 4: 5508-5521. https://doi.org/10.3390/ijms15045508